Factor sensible a la N-etilmaleimida
El factor sensible a la N-etilmaleimida, o proteína de fusión sensible a la N-etilmaleimida, conocida a menudo como NSF —siglas de su nombre en inglés N-ethylmaleimide-sensitive factor— es una enzima perteneciente a la familia de las ATPasas. Se encuentra en el citoplasma de las células eucariotas y desempeña una función importante en el proceso de fusión de membranas.[1][2] En el ser humano, es el producto del gen NSF.[3]
| Factor sensible a la N-etilmaleimida o NSF | |||||||||
|---|---|---|---|---|---|---|---|---|---|
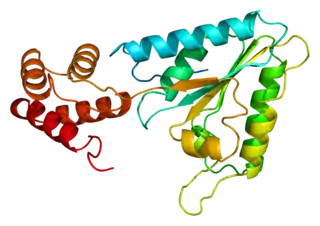 Fragmento de NSF de Cricetulus griseus (PDB 1D2N) | |||||||||
| Estructuras disponibles | |||||||||
| PDB |
Buscar ortólogos: Lista de códigos PDB 3J94, 3J95
| ||||||||
| Identificadores | |||||||||
| Nomenclatura |
Otros nombres Proteína de fusión sensible a la N-etilmaleimida
| ||||||||
| Identificadores externos |
Bases de datos de enzimas
| ||||||||
| Número EC | 3.6.4.6 | ||||||||
| |||||||||
| Estructura/Función proteica | |||||||||
| Peso molecular | 82 594 (Da) | ||||||||
| Tipo de proteína | ATPasa | ||||||||
| Datos enzimáticos | |||||||||
| Cofactor(es) | Mg(II) | ||||||||
| Datos del Receptor/Ligando | |||||||||
| Agonistas | ADP | ||||||||
| Información adicional | |||||||||
| Localización subcelular | Citoplasma celular | ||||||||
| Ortólogos | |||||||||
| Especies |
| ||||||||
| UniProt |
| ||||||||
| RefSeq (proteína) NCBI |
| ||||||||
| PubMed (Búsqueda) |
| ||||||||
| PMC (Búsqueda) |
| ||||||||
Historia
En 1987, el grupo de investigación de James Rothman en la Universidad de Stanford observaron que la adición de N-etilmaleimida inactivaba una molécula citoplasmática participante en las fusiones de membranas, por lo que recibió el nombre de factor sensible a la N-etilmaleimida.[4] En 1988 purificaron la proteína, y un año más tarde descubrieron que se asociaba con un receptor de membrana y una proteína a la que denominaron SNAP (soluble NSF attachment protein o proteína de fijación soluble de NSF), ambos implicados también en la fusión de membrana. Rothman obtuvo la secuencia el gen NSF a partir de células del ovario del hámster chino y descubrió que era similar a la del gen SEC18 de la levadura Saccharomyces cerevisiae, esencial para el transporte de vesículas en este organismo, lo que sugiere que el mecanismo de fusión de membranas es muy similar incluso entre especies eucariotas muy diferentes.[5]
En 1993, Rothman y Richard Scheller determinaron que las proteínas SNARE, otras componentes clave en el transporte de vesícula formaban un complejo muy estable que se disociable por hidrólisis mediada por la NSF en asociación con la proteína SNAP. La hipótesis aceptada actualmente es que la formación del complejo SNARE marca el inicio del proceso de fusión de membranas, y el rol de la NSF consiste en la separación del complejo para liberar los componentes y facilitar el inicio de un nuevo ciclo de fusión.[5]
En 1997 se publicaron los primeros estudios de la estructura de la NSF por microscopia de electrones y los cambios experimentados por la proteína en la ausencia y presencia de ATP y ADP y en combinación con el complejo SNARE.[6] En 1998 y 1999 dos grupos, ambos de la Universidad de Stanford, elucidaron las estructuras a alta resolución del dominio D2 o dominio de hexamerización con y sin ATP,[7][8] y del dominio N-terminal de la NSF.[9][10]
Función
NSF es una ATPasa de la superfamilia AAA (ATPasas Asociadas a diversas Actividades). NSF está conformada por seis subunidades idénticas y participa en el proceso de fusión de membranas.[11] Se halla abundantemente en el citoplasma de las células eucarióticas y desempeña un papel importante en el transporte de vesículas entre diversos compartimentos celulares, que conlleva la fusión de las membranas de la vesícula y su compartimento de destino. Durante este proceso, las proteínas SNARE presentes en las membranas a fusionar forman un complejo muy estable. La NSF deshace este complejo una vez la fusión ha tenido lugar, liberando las proteínas SNARE para futuras fusiones.[12][13]
Debido a su papel en la secreción de neurotransmisores, la inhabilitación de la NSF puede conllevar el bloqueo de la función neuronal. Esto se ha observado en Drosophila melanogaster, donde una mutación del gen NSF causa el coma en las moscas, motivo por el que el gen se conoce como «comatoso» en esta especie. En la ameba Dictyostelium discoideum, mutaciones semejantes causan el cese del movimiento celular, lo que indica que el transporte de membrana desempeña un papel en la migración celular. La asociación de NSF a la subunidad GluR2 del receptor de glutamato AMPA, involucrado en la detección del neurotransmisor glutamato, indica un posible para NSF en la expresión y transporte de los receptores AMPA en la sinapsis.[14] Además del receptor AMPA, NSF se asocia con tors receptores celulares, como el receptor adrenérgico β2, el receptor de dopamina el receptor de adrnomedulina y el receptor de GABA, y con diversas proteínas. No se conoce con exactitud el papel de NSF en todas sus interacciones, aunque se sospecha que su función como chaperona está relacionada con la formación de complejos entre proteínas y su disociación.[15]
Estructura
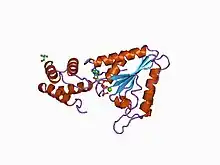
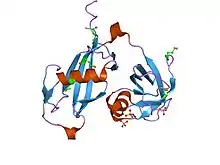
El factor sensible a la N-etilmaleimida es un hexámero en forma de cilindro hueco compuesto por seis unidades idénticas. Cada una de las componentes consta de tres dominios:[15]
- El dominio aminoterminal, o NSF-N, que se asocia a las proteínas SNAP y SNARE mediante un surco en su superficie.
- El primer dominio de unión al ATP (NSF-D1), implicado en la disociación del complejo SNARE. Aunque se desconoce su estructura, se piensa que es similar a la del segundo dominio de unión al ATP por sus similares secuencias de aminoácidos.
- El segundo dominio de unión al ATP (NSF-D2), necesario para la formación del hexámero. El NSF-D2 fue la primera proteína AAA (ATPasas Asociadas a diversas Actividades) de estructura conocida y consta de un subdominio con dos motivos Walker (A y B) y otro subdominio C-terminal con cuatro hélices alfa.[7][8]
El hexámero adopta formas ligeramente diferentes cuando se une a ATP o ADP.[15]
Referencias
- Wilson DW, Whiteheart SW, Wiedmann M, Brunner M, Rothman JE (mayo de 1992). «A multisubunit particle implicated in membrane fusion». The Journal of Cell Biology (en inglés) 117 (3): 531-538. PMC 2289450. PMID 1315316. doi:10.1083/jcb.117.3.531.
- Hoyle J, Phelan JP, Bermingham N, Fisher EM (Noviembre de 1996). «Localization of human and mouse N-ethylmaleimide-sensitive factor (NSF) gene: a two-domain member of the AAA family that is involved in membrane fusion». Mammalian Genome (en inglés) 7 (11): 850-852. PMID 8875895. doi:10.1007/s003359900249.
- «Entrez Gene: NSF N-ethylmaleimide-sensitive factor» (en inglés).
- Glick BS, Rothman JE (1987). «Possible role for fatty acyl-coenzyme A in intracellular protein transport». Nature 326 (6110): 309-12. PMID 3821906. doi:10.1038/326309a0.
- Hussain, S.; Davanger, S. (5 de septiembre de 2011). «The discovery of the soluble N-ethylmaleimide-sensitive factor attachment protein receptor complex and the molecular regulation of synaptic vesicle transmitter release: the 2010 Kavli Prize in neuroscience». Neuroscience (en inglés) 190: 12-20. doi:10.1016/j.neuroscience.2011.05.057.
- Hanson, Phyllis I; Roth, Robyn; Morisaki, Hiroshi; Jahn, Reinhard; Heuser, John E (8 de agosto de 1997). «Structure and Conformational Changes in NSF and Its Membrane Receptor Complexes Visualized by Quick-Freeze/Deep-Etch Electron Microscopy». Cell (en inglés) 90: 523-535. doi:10.1016/S0092-8674(00)80512-7.
- Lenzen, Christian U; Steinmann, Diana; Whiteheart, Sidney W; Weis, William I (21 de agosto de 1998). «Crystal Structure of the Hexamerization Domain of N-ethylmaleimide–Sensitive Fusion Protein». Cell (en inglés) 94: 523-535. doi:10.1016/S0092-8674(00)81593-7.
- Yu, P.I; Hanson; Jahn, R; Brunger, A.T (1998). «Structure of the ATP-dependent oligomerization domain of N-ethylmaleimide sensitive factor complexed with ATP». Nature Structural Biology (en inglés) 5: 803-811. doi:10.1038/1843.
- Yu, R.C; Jahn, R; Brunger, A.T (1999). «NSF N-terminal domain crystal structure: models of NSF function.». Mol. Cell (en inglés) 4: 97-107. doi:10.1016/S1097-2765(00)80191-4.
- May, A.P; Misura, K.M; Whiteheart, S.W; Weis, W.I (1999). «Crystal structure of the amino-terminal domain of N-ethylmaleimide-sensitive fusion protein». Nat.Cell Biol. (en inglés) 1: 175-182. doi:10.1038/11097.
- Furst J, Sutton RB, Chen J, Brunger AT, Grigorieff N (Sep 2003). «Electron cryomicroscopy structure of N-ethyl maleimide sensitive factor at 11 A resolution». The EMBO Journal (en inglés) 22 (17): 4365-74. PMC 202363. PMID 12941689. doi:10.1093/emboj/cdg420.
- Bustillo Merino, Diego; Gutiérrez Martín, Yolanda; Rodríguez Artalejo, Antonio (2009). «La maquinaria molecular de la exocitosis: ¿un nuevo marcador en las enfermedades neurodegenerativas?». Monografía XXV: Avances en Neurociencia: Neurotransmisores y Patologías Nerviosas (Real Academia Nacional de Farmacia): 75. Archivado desde el original el 7 de octubre de 2017. Consultado el 8 de julio de 2017.
- Voet, Donald; Voet, Judith G. (2006). «Fusión de vesículas». Bioquímica. Médica Panamericana. pp. 445-450. ISBN 9789500623018.
- Noel J, Ralph GS, Pickard L, Williams J, Molnar E, Uney JB, Collingridge GL, Henley JM (Jun 1999). «Surface expression of AMPA receptors in hippocampal neurons is regulated by an NSF-dependent mechanism». Neuron 23 (2): 365-76. PMID 10399941. doi:10.1016/S0896-6273(00)80786-2.
- Zhao, Chunxia; Slevin, John T.; Whiteheart, Sidney W. (2007). «Cellular Functions of NSF: Not Just SNAPs and SNAREs». FEBS Letters (en inglés) 581 (11): 2140-2149. doi:10.1016/j.febslet.2007.03.032.
 Datos: Q14860336
Datos: Q14860336