Ácido γ-aminobutírico
El ácido γ-aminobutírico o ácido gamma-aminobutírico, conocido generalmente por sus siglas en inglés, GABA (gamma-aminobutyric acid), es un neurotransmisor que se encuentra presente ampliamente en microorganismos, plantas y animales. Es el principal neurotransmisor inhibidor en el sistema nervioso central (SNC) de mamíferos.[3][4]
| Ácido gamma-aminobutírico | ||
|---|---|---|
 Fórmula estructural simplificada | ||
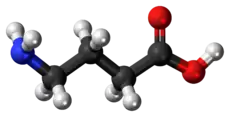 C=negro, H=blanco, O=rojo, N=azul | ||
| Nombre IUPAC | ||
| Ácido 4-aminobutanoico | ||
| General | ||
| Fórmula semidesarrollada | C4H9NO2 | |
| Fórmula molecular | ? | |
| Identificadores | ||
| Número CAS | 56-12-2[1] | |
| Número RTECS | ES6300000 | |
| ChEBI | 16865 | |
| ChEMBL | CHEMBL96 | |
| ChemSpider | 116 | |
| DrugBank | DB02530 | |
| PubChem | 119 | |
| UNII | 2ACZ6IPC6I | |
| KEGG | D00058 | |
|
C(CC(=O)O)CN
| ||
| Propiedades físicas | ||
| Apariencia | Polvo blanco cristalino | |
| Densidad | 1110 kg/m³; 1,11 g/cm³ | |
| Masa molar | 103,12 g/mol | |
| Punto de fusión | 203,7 °C (477 K) | |
| Punto de ebullición | 247,9 °C (521 K) | |
| Propiedades químicas | ||
| Acidez | 4.23 (carboxyl), 10.43 (amino)[2] pKa | |
| Solubilidad en agua | 130 g/100 mL | |
| log P | −3.17 | |
| Riesgos | ||
| Riesgos principales | Irritante, dañino | |
| LD50 | 12,680 mg/kg (ratón, oral) | |
| Valores en el SI y en condiciones estándar (25 ℃ y 1 atm), salvo que se indique lo contrario. | ||
Desempeña el papel principal en la reducción de excitabilidad neuronal a lo largo del sistema nervioso. En humanos, GABA es directamente responsable de la regulación del tono muscular y en aves migratorias de su hiperactividad nocturna y disminución del tiempo dedicado al sueño.[4][5]
A pesar de que, en términos químicos, es un aminoácido, en las comunidades científica y médica rara vez se refieren a GABA como tal debido a que el término "aminoácido" por convención se refiere a los α aminoácidos y GABA no lo es. Además no se considera como parte de alguna proteína.
En la diplejía espástica en humanos desde temprana edad, la absorción de GABA se ve afectada de forma negativa por los nervios dañados por la lesión en las neuronas superiores motoras propias de la condición lo cual lleva a desarrollar hipertonía muscular señalizada por aquellos nervios que son incapaces de absorber GABA.
Historia
El ácido γ-aminobutírico fue sintetizado por primera vez en 1883, y fue conocido originalmente únicamente como producto metabólico de plantas y microorganismos. En 1950, sin embargo, se descubrió que GABA era una parte integral del sistema nervioso central.[6]
El GABA natural fue descubierto el año 1949 en el tejido de la patata.[3]
En el año 1950, fue encontrado en el cerebro de mamíferos por el Dr. Jorge Awapara et al, y por Roberts y Frankel. Estos últimos descubrieron que el GABA se producía a partir del glutamato.[4]
En los años 1970, se descubrió que el GABA está involucrado en la esporulación de Neurospora crassa y Bacillus megaterium.[3]
A fines de los años 1990, se descubrió que el GABA confiere resistencia al pH ácido a bacterias como E. coli, Lactococcus lactis, Listeria monocytogenes, Mycobacterium y Clostridium perfringens.[3]
Estructura
GABA se encuentra principalmente como ion dipolar o zwitterion, esto es, con un grupo carboxilo desprotonado y el grupo amino protonado. Su conformación depende de su entorno. En el estado gaseoso, una conformación altamente plegada es fuertemente favorecida debido a la atracción electrostática entre ambos grupos funcionales. Su estabilización es aproximadamente 50 kcal/mol, de acuerdo a cálculos de química cuántica. En su estado sólido, se favorece una conformación más extendida, con una conformación trans en el grupo amino terminal y una conformación gauche en el grupo carboxilo terminal. Esto se debe a las interacciones de su forma plegada con las moléculas vecinas. En solución, se encuentran cinco conformaciones distintas, algunas plegadas y otras extendidas, como resultado de los efectos de solvatación. La flexibilidad conformacional de GABA es importante para su función biológica, puesto que se ha encontrado unido a diferentes receptores con diferentes conformaciones. Muchos análogos de GABA con aplicaciones farmacéuticas tienen estructuras más rígidas para poder controlar la unión a receptores de mejor manera.[7][8]
Biosíntesis
El GABA endógeno no penetra la barrera hematoencefálica; es sintetizado en el cerebro a partir del glutamato mediante la enzima ácido glutámico descarboxilasa (GAD) y el piridoxal fosfato (una forma activa de la vitamina B6) como cofactor. GABA se transforma de nuevo en glutamato mediante una ruta metabólica llamada GABA shunt. Este proceso convierte el glutamato, el neurotransmisor excitatorio principal, en el neurotransmisor inhibidor principal (GABA).[9][10]
Catabolismo
La enzima GABA transaminasa cataliza la conversión del ácido 4-aminobutanóico (GABA) y el 2-oxoglutarato (α cetoglutarato) en semialdehído succínico y glutamato. El semialdehído succinico es posteriormente oxidado en ácido succínico mediante acción de la enzima AKR7A2 y como tal entra al ciclo de Krebs como fuente de energía.[11]
Función
Neurotransmisor
En vertebrados, el GABA actúa en las sinapsis inhibidoras del cerebro uniéndose a receptores transmembranales específicos en la membrana plasmática tanto de los procesos presinápticos como postsinápticos. Esta unión provoca la apertura de canales iónicos que permiten el flujo tanto de iones cloruro hacia dentro como el flujo de cationes de potasio hacia fuera. Esto resulta en un cambio negativo en el potencial transmembranal, generalmente provocando una hiperpolarización.
Receptor
Se conocen dos clases principales de receptores GABA:
- el GABAA, en el que el receptor forma parte de un complejo de canal iónico regulado por ligando, y
- el receptor metabotrópico GABAB los cuales son receptores acoplados a proteínas G que abren o cierran los canales iónicos por medio de proteínas G intermediarias.

Las neuronas que producen y secretan GABA son conocidas como neuronas GABAérgicas, y tienen principalmente funciones de inhibición en los receptores de vertebrados adultos. Las células espinosas medias (MDC por sus siglas en inglés) son el típico ejemplo de células GABAérgicas inhibidoras del SNC. En cambio, GABA tiene funciones tanto inhibidoras como excitatorias en insectos, mediando la activación muscular en las sinapsis entre los nervios y las células musculares, y también en la estimulación de ciertas glándulas.[12] En mamíferos, algunas neuronas GABAérgicas, tales como las neuronas candelabro, son también capaces de excitar sus contrapartes glutamaérgicas.[13]
Los receptores de GABAA son canales de cloruro activados por la unión a ligando; esto es, permiten el paso a través de la membrana de iones cloruro una vez activados por la unión a GABA. El que este flujo sea excitatorio/despolarizante (hacer la diferencia de potencial (voltaje) menos negativa), shunting (no tiene efecto alguno en la membrana) o inhibitorio/hiperpolarizante (hacer la diferencia de potencial más negativa) depende en la dirección del flujo de los iones cloruro. Cuando el flujo neto de iones cloruro es hacia el exterior, GABA cumple su función excitatoria/despolarizante. Cuando el flujo neto es hacia el interior de la célula, GABA funge como inhibidor/hiperpolarizante. Cuando el flujo neto del cloruro es cercano a cero, la acción de GABA se conoce como shunting. Este tipo de inhibición shunting no tiene efecto directo en el potencial de la membrana de la célula; sin embargo, minimiza el efecto de cualquier estímulo sináptico simultáneo al reducir la resistencia eléctrica de la membrana celular (en esencia, equivalente a la ley de Ohm). Un interruptor en la maquinaria molecular que controla la concentración de cloruro, y por lo tanto la dirección del flujo iónico, es responsable de los cambios en la función de GABA entre las etapas neonatal y adulta. Esto es que el papel que desempeña GABA cambia de excitatorio a inhibidor conforme el cerebro se desarrolla hacia la adultez.[14]
Desarrollo cerebral
Aunque GABA es un neurotransmisor inhibitorio en el cerebro maduro, sus funciones son principalmente excitatorias en el cerebro en desarrollo.[14][15] El gradiente de cloruro se invierte en las neuronas inmaduras y su potencial inverso es mayor que el potencial de membrana en reposo; por lo tanto, la activación de los receptores de GABAA provoca un flujo de salida de la célula de iones cloruro (una corriente despolarizante). El gradiente diferencial de cloruro en las neuronas inmaduras se debe principalmente a las altas concentraciones de cotransportadores de NKCC1 respecto a los cotransportadores de KCC2 en las células inmaduras. GABA es parcialmente responsable por sí mismo de orquestar la maduración de bombas iónicas.[16] Las interneuronas GABAérigicas maduran más rápido en el hipocampo y la maquinaria de señalización de GABA aparece en una etapa más temprana que la transmisión glutamaérgica. Por lo tanto, GABA es el neurotransmisor excitatorio más importante en muchas regiones del cerebro durante la maduración de las sinapsis glutamaérgicas. No obstante, se ha cuestionado esta teoría basándose en evidencia que muestra que en rebanadas de cerebro de ratones incubadas en líquido cefalorraquídeo artificial (modificado de tal forma que toma en cuenta la composición normal del entorno neuronal de crías que aún no han sido destetadas al añadir una fuente de energía diferente de la glucosa, el ácido betahidroxibutírico) la función de GABA cambia de excitatoria o inhibidora.[17]
Este efecto se repitió en las rebanadas de cerebro al añadir otras fuentes de energía en el medio suplementado con glucosa, como piruvato y lactato.[18] Investigaciones posteriores del metabolismo del piruvato[19] y el lactato[20] demostraron que los resultados originales no se debieron a las cuestiones relacionadas con las fuentes de energía sino a cambios en el pH como resultado de las acciones de los sustratos como ácidos débiles. Estos argumentos fueron refutados posteriormente sobre la base de otros hallazgos[21][22] que demostraban que cambios en el pH aún mayores a los provocados por las fuentes de energía no afectaban al cambio en las funciones de GABA descrito en la presencia líquido cefalorraquídeo artificial suplementado con otras fuentes de energía y que el modo de acción del ácido betahidroxibutírico, el piruvato y lactato (evaluado por las mediciones del consumo de NAD(P)H y oxígeno) estaba relacionado con el metabolismo de estos.[23]
En las etapas de desarrollo que preceden a la formación de contactos sinápticos, GABA es sintetizado por neuronas y actúa tanto como mediador de señalización autocrina (actúa en la misma célula que lo secreta) como paracrina (actúa en las células próximas).[24][25] La eminencia gangliónica (EG) también contribuye en gran parte en la generación de población de células GABAérgicas corticales.[26]
GABA regula la proliferación,[27][28] migración,[29] y diferenciación de progenitores neurales,[16][30] la elongación de neuritas[31] y la formación de sinapsis.[32]
GABA también regula el crecimiento de las células madre embrionarias y neurales. GABA puede influir en el desarrollo de progenitores neurales por medio de la expresión del factor neurotrófico derivado del cerebro (BDNF por sus siglas en inglés).[33] GABA también es capaz de provocar la detención del ciclo celular en la fase S al activar los receptores GABAA, limitando el crecimiento.[34]
Más allá del sistema nervioso

Se han demostrado mecanismos GABAérgicos en distintos tejidos y órganos periféricos incluyendo el intestino, estómago, páncreas, las trompas de falopio, el útero, ovario, testículos, riñón, vejiga, pulmón e hígado.[36]
En el 2007, un sistema excitatorio GABAérgico fue descrito en el epitelio de las vías respiratorias. El sistema se activa después de la exposición a alergénos y puede estar involucrado en los mecanismos del asma.[37] Los sistemas GABAérgicos también se han encontrado en los testículos[38] y en el cristalino de los ojos.[39]
Farmacología
Los medicamentos que actúan como moduladores alostéricos de los receptores GABA (conocidos como análogos de GABA o medicamentos GABAérgicos) o aumentan la cantidad disponible de GABA generalmente tienen efectos de relajación, combaten la ansiedad y tienen efectos anticonvulsivantes.[40][41] Se sabe que muchas de estas sustancias provocan amnesia anterógrada y retrógrada.[42]
En general GABA no cruza la barrera hematoencefálica,[43] aunque algunas zonas del cerebro que no poseen una barrera hematoencefálica efectiva, tal como el núcleo periventricular, pueden ser afectadas por estos compuestos como la GABA inyectada de manera sistémica.[44] Al menos un estudio sugiere que GABA administrado oralmente incrementa la cantidad de hormona humana de crecimiento.[45] Se ha reportado que GABA inyectado directamente al cerebro tiene efectos tanto estimulantes como inhibidores en la producción de hormona del crecimiento, dependiendo de la fisiología del individuo.[44] Se han desarrollado algunos profármacos de GABA (por ejemplo, picamilon) para permear la barrera hematoencefálica y separarse en una molécula de GABA y su molécula acarreadora una vez dentro del cerebro. Esto permite un incremento en los niveles de GABA a lo largo del cerebro, de manera que se puede monitorear el patrón de distribución del profármaco antes de su metabolismo.
Se ha visto que GABA influye en el catabolismo de la serotonina hacia N-acetilserotonina y melatonina en ratas.[46] Se sospecha que tiene una función reguladora en la producción de melatonina en humanos.
Medicamentos GABAérgicos
- Ligandos de receptores GABAA
- Agonistas o moduladores alostéricos positivos: etanol,[47][48][49] barbitúricos, benzodiazepinas, cariosprodol, hidrato de cloral, etomidato, glutetimida, kava, metacualona, etacualona, muscimol, neuroesteroides, fármacos z, propofol, scutellaria lateriflora, valeriana común, teanina y anestésicos inhalados/volátiles.
- Antagonistas o modulares alostéricos negativos: bicuculina, cicutoxina, flumazenil, furosemida, gabazina, enantotoxina, picrotoxina, Ro15-4513 y tujona.
- Ligandos de receptores GABAB
- Agonistas: baclofen, gama butirolactona (GBL), propofol, ácido γ-hidroxibutírico (GHB)[50] y fenibut.
- Antagonistas: faclofen y saclofen.
- Inhibidores de recaptura de GABA: hiperforina y tiagabina.
- Inhibidores de la GABA transaminasa: gabaculina, fenelzina, ácido valproico, vigabatrina, y Melissa officinalis.[51]
- Análogos de GABA: pregabalina y gabapentina.
- Otros: GABA por sí mismo, L-glutamina, picamilon y progabide.
GABA como suplemento
Se usa un gran número de formulaciones comerciales de GABA como suplemento alimenticio, algunas veces por administración sublingual. Estos productores aseguran que tiene un efecto calmante. Esto no es completamente irracional dada la naturaleza de GABA, pero aislado por sí mismo no ha sido reconocido científicamente como agente tranquilizante y esto solo se ha demostrado de manera irregular. Por ejemplo, hay evidencia que muestra que se puede observar GABA puro en el cerebro después de una administración oral de GABA como suplemento.[52] Sin embargo, hay evidencia más relevante de que GABA no cruza la barrera hematoencefálica en niveles significantes terapéuticamente en ratones tratados con un inhibidor de GABA transaminasa. En animales no tratados (ratones y conejos) los marcadores de GABA (H3) se encontraron distribuidos en distintos metabolitos no identificados (excepto la glutamina), lo cual parece moverse hacia y desde el cerebro.[43]
Aunque GABA podría no cruzar la barrera hematoencefálica, es importante considerar que los estudios que han demostrado que en individuos con la barrera hematoencefálica dañada (temporalmente con propósitos experimentales, o como resultados de otros problemas), GABA, de hecho, tiene un efecto positivo, aunque con efectos secundarios.[53] La única manera de administrar GABA efectivamente es burlar la barrera hematoencefálica. De hecho, hay un pequeño y limitado número de suplementos disponibles que son derivados de GABA, tales como fenibut y picamilon. Picamilon combina niacina y GABA para cruzar la barrera hematoencefálica como profármaco que después se hidroliza para dar lugar a GABA y niacina.[54]
En plantas
GABA también se puede encontrar en plantas. Es el aminoácido más abundante en el apoplasto del tomate.[55] También podría tener un papel en la señalización en plantas.[56][57]
Referencias
- Número CAS
- Dawson RMC, Elliot DC, Elliot WH, Jones KM, ed. (1959). Data for Biochemical Research. Oxford: Clarendon Press.
- Dhakal, Radhika; Bajpai, Vivek K.; Baek, Kwang-Hyun (octubre a diciembre de 2012). «Production of gaba (γ – Aminobutyric acid) by microorganisms: a review» [Producción de GABA (ácido aminobutírico) por microorganismos: una revisión]. Braz J Microbiol (en inglés) (Elsevier) 43 (4): 1230-1241. PMID 24031948. doi:10.1590/S1517-83822012000400001. Consultado el 22 de agosto de 2017.
- Watanabe, Masahito; Maemura, Kentaro; Kanbara, Kiyoto; Tamayama, Takumi; Hayasaki, Hana (2002). «GABA and GABA receptors in the central nervous system and other organs» [GABA y receptores GABA en el sistema nervioso central y otros órganos]. Int Rev Cytol (en inglés) (Osaka, Japón: Academic Press) 213: 1-47. PMID 11837891. doi:10.1016/S0074-7696(02)13011-7. Consultado el 22 de agosto de 2017.
- Frias-Soler RC, Pildaín LV, Pârâu LG, Wink M, Bairlein F. Transcriptome signatures in the brain of a migratory songbird. Comp Biochem Physiol Part D Genomics Proteomics. 2020 Mar;34 100681. doi:10.1016/j.cbd.2020.100681. PMID: 32222683
- Roth RJ, Cooper JR, Bloom FE (2003). The Biochemical Basis of Neuropharmacology. Oxford [Oxfordshire]: Oxford University Press. p. 106. ISBN 0-19-514008-7.
- Majumdar Devashis, Guha Sephali (1988). «Conformation, electrostatic potential and pharmacophoric pattern of GABA (γ-aminobutyric acid) and several GABA inhibitors». Journal of Molecular Structure: THEOCHEM 180: 125-140. doi:10.1016/0166-1280(88)80084-8.
- Sapse A-M (2000). Molecular Orbital Calculations for Amino Acids and Peptides. Birkhäuser. p. 28. ISBN 978-0-8176-3893-1.
- Petroff OA (diciembre de 2002). «GABA and glutamate in the human brain». Neuroscientist 8 (6): 562-573. PMID 12467378. doi:10.1177/1073858402238515.
- Schousboe A, Waagepetersen HS (2007). «GABA: homeostatic and pharmacological aspects». Prog. Brain Res. Progress in Brain Research 160: 9-19. ISBN 978-0-444-52184-2. PMID 17499106. doi:10.1016/S0079-6123(06)60002-2.
- Bown AW, Shelp BJ (septiembre de 1997). «The Metabolism and Functions of γ-Aminobutyric Acid». Plant Physiol. 115 (1): 1-5. PMC 158453. PMID 12223787. doi:10.1104/pp.115.1.1 (inactivo 2015-01-01).
- Ffrench-Constant RH, Rocheleau TA, Steichen JC, Chalmers AE (junio de 1993). «A point mutation in a Drosophila GABA receptor confers insecticide resistance». Nature 363 (6428): 449-51. Bibcode:1993Natur.363..449F. PMID 8389005. doi:10.1038/363449a0.
- Szabadics J, Varga C, Molnár G, Oláh S, Barzó P, Tamás G (enero de 2006). «Excitatory effect of GABAergic axo-axonic cells in cortical microcircuits». Science 311 (5758): 233-235. Bibcode:2006Sci...311..233S. PMID 16410524. doi:10.1126/science.1121325.
- Li K, Xu E (junio de 2008). «The role and the mechanism of γ-aminobutyric acid during central nervous system development». Neurosci Bull 24 (3): 195-200. PMID 18500393. doi:10.1007/s12264-008-0109-3.
- Ben-Ari Y, Gaiarsa JL, Tyzio R, Khazipov R (octubre de 2007). «GABA: a pioneer transmitter that excites immature neurons and generates primitive oscillations». Physiol. Rev. 87 (4): 1215-1284. PMID 17928584. doi:10.1152/physrev.00017.2006.
- Ganguly K, Schinder AF, Wong ST, Poo M (mayo de 2001). «GABA itself promotes the developmental switch of neuronal GABAergic responses from excitation to inhibition». Cell 105 (4): 521-32. PMID 11371348. doi:10.1016/S0092-8674(01)00341-5.
- Rheims S, Holmgren CD, Chazal G, Mulder J, Harkany T, Zilberter T, Zilberter Y (agosto de 2009). «GABA action in immature neocortical neurons directly depends on the availability of ketone bodies». J. Neurochem. 110 (4): 1330-8. PMID 19558450. doi:10.1111/j.1471-4159.2009.06230.x.
- Holmgren CD, Mukhtarov M, Malkov AE, Popova IY, Bregestovski P, Zilberter Y (febrero de 2010). «Energy substrate availability as a determinant of neuronal resting potential, GABA signaling and spontaneous network activity in the neonatal cortex in vitro». J. Neurochem. 112 (4): 900-12. PMID 19943846. doi:10.1111/j.1471-4159.2009.06506.x.
- Tyzio R, Allene C, Nardou R, Picardo MA, Yamamoto S, Sivakumaran S, Caiati MD, Rheims S, Minlebaev M, Milh M, Ferré P, Khazipov R, Romette JL, Lorquin J, Cossart R, Khalilov I, Nehlig A, Cherubini E, Ben-Ari Y (enero de 2011). «Depolarizing actions of GABA in immature neurons depend neither on ketone bodies nor on pyruvate». J. Neurosci. 31 (1): 34-45. PMID 21209187. doi:10.1523/JNEUROSCI.3314-10.2011.
- Ruusuvuori E, Kirilkin I, Pandya N, Kaila K (noviembre de 2010). «Spontaneous network events driven by depolarizing GABA action in neonatal hippocampal slices are not attributable to deficient mitochondrial energy metabolism». J. Neurosci. 30 (46): 15638-42. PMID 21084619. doi:10.1523/JNEUROSCI.3355-10.2010.
- Mukhtarov M, Ivanov A, Zilberter Y, Bregestovski P (enero de 2011). «Inhibition of spontaneous network activity in neonatal hippocampal slices by energy substrates is not correlated with intracellular acidification». J. Neurochem. 116 (2): 316-21. PMID 21083663. doi:10.1111/j.1471-4159.2010.07111.x.
- Ivanov A, Mukhtarov M, Bregestovski P, Zilberter Y (2011). «Lactate Effectively Covers Energy Demands during Neuronal Network Activity in Neonatal Hippocampal Slices». Front Neuroenergetics 3: 2. PMC 3092068. PMID 21602909. doi:10.3389/fnene.2011.00002.
- Khakhalin AS (mayo de 2011). «Questioning the depolarizing effects of GABA during early brain development». J Neurophysiol 106 (3): 1065-7. PMID 21593390. doi:10.1152/jn.00293.2011.
- Purves D, Fitzpatrick D, Hall WC, Augustine GJ, Lamantia A-S, ed. (2007). Neuroscience (4th edición). Sunderland, Mass: Sinauer. pp. 135, box 6D. ISBN 0-87893-697-1.
- Jelitai M, Madarasz E (2005). «The role of GABA in the early neuronal development». Int. Rev. Neurobiol. International Review of Neurobiology 71: 27-62. ISBN 9780123668721. PMID 16512345. doi:10.1016/S0074-7742(05)71002-3.
- Marín O, Rubenstein JL (noviembre de 2001). «A long, remarkable journey: tangential migration in the telencephalon». Nat. Rev. Neurosci. 2 (11): 780-90. PMID 11715055. doi:10.1038/35097509.
- LoTurco JJ, Owens DF, Heath MJ, Davis MB, Kriegstein AR (diciembre de 1995). «GABA and glutamate depolarize cortical progenitor cells and inhibit DNA synthesis». Neuron 15 (6): 1287-1298. PMID 8845153. doi:10.1016/0896-6273(95)90008-X.
- Haydar TF, Wang F, Schwartz ML, Rakic P (agosto de 2000). «Differential modulation of proliferation in the neocortical ventricular and subventricular zones». J. Neurosci. 20 (15): 5764-74. PMID 10908617.
- Behar TN, Schaffner AE, Scott CA, O'Connell C, Barker JL (agosto de 1998). «Differential response of cortical plate and ventricular zone cells to GABA as a migration stimulus». J. Neurosci. 18 (16): 6378-87. PMID 9698329.
- Barbin G, Pollard H, Gaïarsa JL, Ben-Ari Y (abril de 1993). «Involvement of GABAA receptors in the outgrowth of cultured hippocampal neurons». Neurosci. Lett. 152 (1–2): 150-154. PMID 8390627. doi:10.1016/0304-3940(93)90505-F.
- Maric D, Liu QY, Maric I, Chaudry S, Chang YH, Smith SV, Sieghart W, Fritschy JM, Barker JL (abril de 2001). «GABA expression dominates neuronal lineage progression in the embryonic rat neocortex and facilitates neurite outgrowth via GABA(A) autoreceptor/Cl− channels». J. Neurosci. 21 (7): 2343-60. PMID 11264309.
- Ben-Ari Y (septiembre de 2002). «Excitatory actions of gaba during development: the nature of the nurture». Nat. Rev. Neurosci. 3 (9): 728-739. PMID 12209121. doi:10.1038/nrn920.
- Obrietan K, Gao XB, Van Den Pol AN (agosto de 2002). «Excitatory actions of GABA increase BDNF expression via a MAPK-CREB-dependent mechanism--a positive feedback circuit in developing neurons». J. Neurophysiol. 88 (2): 1005-15. PMID 12163549.
- Wang DD, Kriegstein AR, Ben-Ari Y (2008). «GABA regulates stem cell proliferation before nervous system formation». Epilepsy Curr 8 (5): 137-9. PMC 2566617. PMID 18852839. doi:10.1111/j.1535-7511.2008.00270.x.
- Popp A, Urbach A, Witte OW, Frahm C (2009). «Adult and embryonic GAD transcripts are spatiotemporally regulated during postnatal development in the rat brain». En Reh, Thomas A., ed. PLoS ONE 4 (2): e4371. Bibcode:2009PLoSO...4.4371P. PMC 2629816. PMID 19190758. doi:10.1371/journal.pone.0004371.
- Erdö SL, Wolff JR (febrero de 1990). «γ-Aminobutyric acid outside the mammalian brain». J. Neurochem. 54 (2): 363-72. PMID 2405103. doi:10.1111/j.1471-4159.1990.tb01882.x.
- Xiang YY, Wang S, Liu M, Hirota JA, Li J, Ju W, Fan Y, Kelly MM, Ye B, Orser B, O'Byrne PM, Inman MD, Yang X, Lu WY (julio de 2007). «A GABAergic system in airway epithelium is essential for mucus overproduction in asthma». Nat. Med. 13 (7): 862-7. PMID 17589520. doi:10.1038/nm1604.
- Payne AH, Hardy MH (2007). The Leydig cell in health and disease. Humana Press. ISBN 1-58829-754-3.
- Kwakowsky A, Schwirtlich M, Zhang Q, Eisenstat DD, Erdélyi F, Baranyi M, Katarova ZD, Szabó G (diciembre de 2007). «GAD isoforms exhibit distinct spatiotemporal expression patterns in the developing mouse lens: correlation with Dlx2 and Dlx5». Dev. Dyn. 236 (12): 3532-44. PMID 17969168. doi:10.1002/dvdy.21361.
- Foster AC, Kemp JA (febrero de 2006). «Glutamate- and GABA-based CNS therapeutics». Curr Opin Pharmacol 6 (1): 7-17. PMID 16377242. doi:10.1016/j.coph.2005.11.005.
- Chapouthier G, Venault P (octubre de 2001). «A pharmacological link between epilepsy and anxiety?». Trends Pharmacol. Sci. 22 (10): 491-3. PMID 11583788. doi:10.1016/S0165-6147(00)01807-1.
- Campagna JA, Miller KW, Forman SA (mayo de 2003). «Mechanisms of actions of inhaled anesthetics». N. Engl. J. Med. 348 (21): 2110-24. PMID 12761368. doi:10.1056/NEJMra021261.
- Kuriyama K, Sze PY (enero de 1971). «Blood–brain barrier to H3-γ-aminobutyric acid in normal and amino oxyacetic acid-treated animals». Neuropharmacology 10 (1): 103-108. PMID 5569303. doi:10.1016/0028-3908(71)90013-X.
- Müller EE, Locatelli V, Cocchi D (abril de 1999). «Neuroendocrine control of growth hormone secretion». Physiol. Rev. 79 (2): 511-607. PMID 10221989.
- Powers ME, Yarrow JF, McCoy SC, Borst SE (enero de 2008). «Growth hormone isoform responses to GABA ingestion at rest and after exercise». Medicine and science in sports and exercise 40 (1): 104-10. PMID 18091016. doi:10.1249/mss.0b013e318158b518.
- http://www.ncbi.nlm.nih.gov/pubmed/6844712
- Dzitoyeva S, Dimitrijevic N, Manev H (2003). «γ-aminobutyric acid B receptor 1 mediates behavior-impairing actions of alcohol in Drosophila: adult RNA interference and pharmacological evidence». Proc. Natl. Acad. Sci. U.S.A. 100 (9): 5485-5490. Bibcode:2003PNAS..100.5485D. PMC 154371. PMID 12692303. doi:10.1073/pnas.0830111100.
- Mihic SJ, Ye Q, Wick MJ, Koltchine VV, Krasowski MD, Finn SE, Mascia MP, Valenzuela CF, Hanson KK, Greenblatt EP, Harris RA, Harrison NL (1997). «Sites of alcohol and volatile anaesthetic action on GABAA and glycine receptors». Nature 389 (6649): 385-389. Bibcode:1997Natur.389..385M. PMID 9311780. doi:10.1038/38738.
- Boehm SL, Ponomarev I, Blednov YA, Harris RA (2006). «From gene to behavior and back again: new perspectives on GABAAreceptor subunit selectivity of alcohol actions». Adv. Pharmacol. 54 (8): 1581-1602. PMID 17175815. doi:10.1016/j.bcp.2004.07.023.
- Dimitrijevic N, Dzitoyeva S, Satta R, Imbesi M, Yildiz S, Manev H (2005). «Drosophila GABAB receptors are involved in behavioral effects of gamma-hydroxybutyric acid (GHB)». Eur. J. Pharmacol. 519 (3): 246-252. PMID 16129424. doi:10.1016/j.ejphar.2005.07.016.
- Awad R, Muhammad A, Durst T, Trudeau VL, Arnason JT (agosto de 2009). «Bioassay-guided fractionation of lemon balm (Melissa officinalis L.) using an in vitro measure of GABA transaminase activity». Phytother Res 23 (8): 1075-81. PMID 19165747. doi:10.1002/ptr.2712.
- Abdou AM, Higashiguchi S, Horie K, Kim M, Hatta H, Yokogoshi H (2006). «Relaxation and immunity enhancement effects of gamma-aminobutyric acid (GABA) administration in humans». BioFactors 26 (3): 201-8. PMID 16971751. doi:10.1002/biof.5520260305.
- «A GABA-EEG test of the blood-brain barrier near epileptic foci.».
- Dorofeev BF, Kholodov LE (1991). «Фармакокинетика пикамилона у животных». Farmakologiia i toksikologiia (en ruso) 54 (2): 66-9. PMID 1884802.
- Park DH, Mirabella R, Bronstein PA, Preston GM, Haring MA, Lim CK, Collmer A, Schuurink RC (octubre de 2010). «Mutations in γ-aminobutyric acid (GABA) transaminase genes in plants or Pseudomonas syringae reduce bacterial virulence». Plant J. 64 (2): 318-30. PMID 21070411. doi:10.1111/j.1365-313X.2010.04327.x.
- Bouché N, Fromm H (marzo de 2004). «GABA in plants: just a metabolite?». Trends Plant Sci. 9 (3): 110-5. PMID 15003233. doi:10.1016/j.tplants.2004.01.006.
- Roberts MR (septiembre de 2007). «Does GABA Act as a Signal in Plants?: Hints from Molecular Studies». Plant Signal Behav 2 (5): 408-9. PMC 2634229. PMID 19704616. doi:10.4161/psb.2.5.4335.
Enlaces externos
 Wikimedia Commons alberga una categoría multimedia sobre Ácido γ-aminobutírico.
Wikimedia Commons alberga una categoría multimedia sobre Ácido γ-aminobutírico.
| Control de autoridades |
|
|---|
 Datos: Q210021
Datos: Q210021- Multimedia: Gamma-Aminobutyric acid / Q210021