Microbiota vaginal
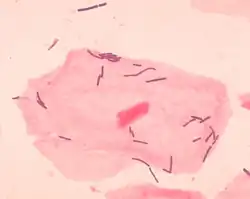
La flora vaginal, la microbiota vaginal o el microbioma vaginal son los microorganismos que colonizan la vagina. Fueron descubiertos por el ginecólogo alemán Albert Döderlein en 1892[1] y son parte de la flora humana general. La cantidad y el tipo de bacterias presentes tienen implicaciones significativas para la salud general de una mujer.[2] Los Lactobacillus son los microorganismos más frecuentemente aislado de la vagina humana sana,[3] esto incluye Lactobacillus crispatus, Lactobacillus gasseri, Lactobacillus iners y Lactobacillus jensenii[4] y se cree que el ácido láctico que producen protege contra la infección por especies patógenas.[5]
| Microbiota humana | ||
|---|---|---|
| Flora humana | ||
| Trastornos y terapias | ||
| Relacionado: | ||
Lactobacilos
Las principales bacterias colonizadoras de un individuo sano son del género Lactobacillus.[3] Desde la primera descripción de los lactobacilos por Döderlein, los lactobacilos se han considerado generalmente los guardianes del ecosistema vaginal. Se ha demostrado que los lactobacilos inhiben el crecimiento in vitro de microorganismos patógenos, por ejemplo, Bacteroides fragilis, Escherichia coli, Gardnerella vaginalis, Mobiluncus spp., Neisseria gonorrhoeae, Peptostreptococcus anaerobius, P. bivia y Staphylococcus aureus. En general, se acepta que esto se logra principalmente a través de la acción del ácido láctico.[6][7][8][9] Además, los lactobacilos normalmente ayudan a prevenir la colonización a largo plazo de la vagina al adherirse a las células epiteliales vaginales. Esto generalmente reduce la infección de patógenos en el epitelio vaginal.[10]
Además de la producción de ácido láctico y la competencia por la adherencia, otros mecanismos antagónicos incluyen la producción de peróxido de hidrógeno (un antimicrobiano de amplio espectro) y bacteriocinas (antimicrobianos específicos del objetivo).[11][12]
La producción de ácido láctico

Generalmente se acepta que el pH bajo es el mecanismo principal que controla la composición de la microflora vaginal. Aunque el ácido láctico producido por los lactobacilos contribuye a la acidez vaginal, todavía no se ha demostrado que sea la fuente principal de pH vaginal bajo, pero el hecho es que la mayoría de los lactobacilos prosperan mejor a un pH <4.5.[13][14][15]
Peróxido de hidrógeno
La producción de peróxido de hidrógeno (H2O2) es un mecanismo bien conocido para el antagonismo bacteriano,[16][17][18] inhibir el crecimiento de microorganismos a través de la interacción directa o mediante mieloperoxidasa humana.[19] [20][21] Se ha demostrado que los lactobacilos productores de peróxido de hidrógeno inactivan el VIH-1, el virus del herpes simple tipo 2 (HSV-2), Trichomonas vaginalis, G. vaginalis, P. bivia y E. coli. O'Hanlon[12] y Baeten[22] encontraron que el 96% de las especies de Lactobacillus a partir de un ecosistema vaginal sano produce H2O2 (L. jensenii y L. vaginalis produce los niveles más altos de H2O2),[11][23] mientras que sólo el 6% de los lactobacilos se recuperó de las mujeres con BV produce H2O2. De acuerdo con esto, L. iners, más frecuentemente asociado con microflora vaginal perturbado,[24][25] es un pobre productor de H2O2.[26][27] La colonización vaginal por lactobacilos productores de H2O2 se ha asociado con una disminución en la aparición de vaginosis bacteriana (BV).[28] Sin embargo, O'Hanlon et al.[29] demostraron que el fluido cervicovaginal y semen tienen una significativa actividad bloqueante de H2O2 bloqueante, también demostraron que las concentraciones fisiológicas de H2O2 por debajo de 100μM fallan para inactivar cualquiera de las 17 bacterias asociadas a la BV, analizadas, por ejemplo A. vaginae, G. vaginalis, Mobiluncus spp., P. bivia, Prevotella corporis, Mycoplasma hominis, incluso en presencia de mieloperoxidasa humana, conocida para aumentar la actividad microbicida de H2O2. Solo las concentraciones suprafisiológicas de H2O2 exógeno (0.34% p/v, 100 mM) fueron suficientes para inactivar las bacterias asociadas a BV a cuya concentración inactivaba más potentemente los lactobacilos vaginales (L. crispatus, L. gasseri, L. iners y L . jensenii). Una concentración de H2O2 de100 mM es aproximadamente 50 veces mayor que la que los lactobacilos son capaces de producir incluso en condiciones óptimas aeróbicas, bajas en antioxidantes, y aproximadamente 5,000 veces mayor que la concentración estimada de H2O2 in vivo. Aún más notable, la adición de solo 1% de fluido vaginal bloqueó la actividad microbicida de 1 M de H2O2. Las posibles explicaciones pueden ser que el fluido cervicovaginal y el semen contienen proteínas, glicoproteínas, polisacáridos, lípidos y otras moléculas con el potencial de reaccionar con e inactivar el H2O2. Además, la vagina es hipóxica la mayor parte del tiempo, mientras que los lactobacilos requieren oxígeno para producir peróxido de hidrógeno. También es notable que la catalasa, que proporciona protección bacteriana contra el H2O2 tóxico, esté ausente en los lactobacilos,[30] y, como tales, no estarían protegidos contra su propia producción de H2O2. Por el contrario, en condiciones óptimas de crecimiento anaeróbico, las concentraciones fisiológicas de ácido láctico inactivaron los patógenos asociados a la BV sin afectar los lactobacilos vaginales. En resumen, aunque la producción de peróxido de hidrógeno de los lactobacilos se ha considerado como un componente antimicrobiano importante, contribuyendo a la resistencia a la colonización proporcionado por los lactobacilos,[31] y, aunque no parece haber una relación entre lactobacilos productores de H2O2 y microflora vaginal normal, los datos recientes no apoyan este papel para el H2O2.
Bacteriocinas
Los lactobacilos vaginales producen péptidos antimicrobianos, es decir, bacteriocinas como la lactocina 160 y crispasina.[13] con actividad inhibitoria que va desde estrechas (especies de Lactobacillus estrechamente relacionadas) hasta amplias (diversos grupos de bacterias, incluidas G. vaginalis y P. bivia),[8] y sustancias similares a las bacteriocinas, con un espectro de actividad más amplio que las bacteriocinas (por ejemplo, un péptido resistente al calor producido por Lactobacillus salivarius subsp. salivarius CRL 1328). Varios estudios han indicado que la actividad de las bacteriocinas se ve favorecida por el bajo pH.
Las sustancias inhibidoras producidas por el Lactobacillus vaginal son un factor primario en la protección de la microbiota vaginal, con ácidos orgánicos, bacteriocinas y peróxido de hidrógeno. Estos actúan sinérgicamente contra la infección por patógenos. No todos los Lactobacillus spp. y no todas las cepas dentro de una especie de Lactobacillus exhiben los 3 mecanismos.[13] Las especies de Lactobacillus difieren en mujeres premenopáusicas, es decir L. crispatus, L. jensenii, L. iners, L. gasseri (y posiblemente L. vaginalis ), según lo evaluado a través de técnicas dependientes e independientes del cultivo.[24][25][26][32] Se ha demostrado que los lactobacilos vaginales muestran un vaginotropismo pronunciado, y sus pili actúan como ligandos para la unión a los receptores de las células epiteliales vaginales. El número limitado de Lactobacillus spp. El hallazgo en la vagina humana es notable, lo que lleva a la posibilidad de que haya factores del huésped que seleccionan organismos específicos, que estas especies tienen características inusuales que les permiten colonizar con éxito la vagina, o ambas.[33] Sin embargo, el vaginotropismo no solo se aplica a este grupo seleccionado de lactobacilos que representan una vagina sana, sino también a las especies bacterianas asociadas con la VB.[34] La microbiota detectada en el genital humano y el intestino económico no parece crecer fuera de su huésped y es probable que dependa del contacto cercano entre los padres y sus hijos para la transmisión, por ejemplo, la transmisión materno-neonata de la microflora genital, muy probablemente también con microflora intestinal distribuida homogéneamente sobre el cuerpo del bebé, incluida la piel, la cavidad oral, la nasofaringe y las heces.[35]
Otra microbiota
La microbiota vaginal sana y normal que está dominada por lactobacilos puede diferir entre algunos grupos étnicos. Las especies vaginales no patógenas son parte de la microbiota normal de algunas mujeres.[24][36] Varios estudios han demostrado que una proporción significativa (7–33%) de mujeres asintomáticas sanas (especialmente mujeres negras e hispanas)[37] carecen de un número apreciable de especies de Lactobacillus en la vagina,[33][38] y en cambio tienen una microbiota vaginal que consisten en otras bacterias productoras de ácido láctico, es decir, especies de los géneros Atopobium, Leptotrichia, Leuconostoc, Megasphaera, Pediococcus, Streptococcus y Weissella,[32] Todas las poblaciones étnicas tienen comunidades de microflora vaginal que contienen bacterias productoras de ácido láctico. Esto implica que no todas las comunidades pueden ser igualmente resistentes, de modo que si la resistencia de una comunidad vaginal es baja, los cambios transitorios en la estructura de estas comunidades pueden ocurrir más fácilmente en respuesta a disturbios de diversos tipos, incluyendo menstruaciones, relaciones sexuales, duchas vaginales y prácticas anticonceptivas. Estas diferencias en la estructura y composición de las comunidades microbianas pueden ser la base de diferencias bien conocidas en la susceptibilidad de las mujeres en estos grupos raciales a BV y varias infecciones vaginales vaginal.[39][40] Aunque la microflora vaginal puede verse alterada y otras especies producen ácido láctico,[41] el pH vaginal y la producción de ácido establecen una microbiota vaginal normal. El pH disminuye aún más durante el embarazo.[42]
Otras especies bacterianas vaginales
Otras especies bacterianas se encuentran con frecuencia en la vagina, como los cocos Gram positivos: Atopobium vaginae, Peptostreptococcus spp., Staphylococcus spp., Streptococcus spp., Y Bacteroides spp., Fusobacterium spp., Gardnerella vaginalis, Mobiluncus, Prevotella spp. y organismos entéricos gramnegativos, como Escherichia coli.[24][25] Micoplasma y Ureaplasma se encuentran con frecuencia en la vagina. Algunas de las bacterias anaerobias obligatorias y facultativas están asociadas con la BV.[38]
El embarazo
Se debate el efecto del uso de tampones en la flora vaginal, pero la aplicación de tampones parece no modificar significativamente el equilibrio de la presencia bacteriana. embarazo altera la microbiota con una reducción en la diversidad de especies/géneros.[43]
La prevención de enfermedades
Un microbioma vaginal saludable ayuda a prevenir la vaginosis bacteriana, las infecciones por hongos y otros posibles problemas al mantener un pH ácido (<4.5) que es desfavorable para el crecimiento de patógenos comunes, como Gardnerella vaginalis. Los lactobacilos presentes en un microbioma vaginal saludable también ocupan el nicho ecológico que de otro modo estaría disponible para su explotación por organismos patógenos. Sin embargo, las bacterias dañinas o un desequilibrio en las bacterias (por factores como la menstruación, el embarazo, la práctica sexual, el uso incontrolado de antibióticos y las duchas vaginales) pueden provocar infecciones infecciones vaginales como la vaginosis bacteriana (BV), infecciones de transmisión sexual (ITS) o la candidiasis vulvovaginal (VVC).[4]
La vaginosis bacteriana se asocia con la presencia de Gardnerella vaginalis y Peptostreptococcus anaerobius[44] y una disminución en el número de especies de Lactobacillus que comprenden la microbiota vaginal sana.[43][45][46][47]
Investigación
Las investigaciones han encontrado que la presencia de bacterias dominadas por lactobacilos en la vagina se asocia con una menor incidencia de infecciones de transmisión sexual.[48][49]
Véase también
Referencias
- David, M. (2006). «Albert und Gustav Döderlein – ein kritischer Blick auf zwei besondere Lebensläufe deutscher Ordinarien». Zentralblatt für Gynäkologie 128 (2): 56-59. ISSN 0044-4197. PMID 16673245. doi:10.1055/s-2006-921412.
- D'Ippolito, Silvia; Di Nicuolo, Fiorella; Pontecorvi, Alfredo; Gratta, Matteo; Scambia, Giovanni; Di Simone, Nicoletta (2018). «Endometrial microbes and microbiome: Recent insights on the inflammatory and immune "players" of the human endometrium». American Journal of Reproductive Immunology. e13065 (6): e13065. PMID 30375712. doi:10.1111/aji.13065.
- Vasquez, A.; Jakobsson, T.; Ahrne, S.; Forsum, U.; Molin, G. (2002). «Vaginal Lactobacillus Flora of Healthy Swedish Women». Journal of Clinical Microbiology 40 (8): 2746-2749. PMC 120688. PMID 12149323. doi:10.1128/JCM.40.8.2746-2749.2002.
- Chee, Wallace Jeng Yang; Chew, Shu Yih; Than, Leslie Thian Lung (7 de noviembre de 2020). «Vaginal microbiota and the potential of Lactobacillus derivatives in maintaining vaginal health». Microbial Cell Factories 19 (1): 203. ISSN 1475-2859. PMC 7648308. PMID 33160356. doi:10.1186/s12934-020-01464-4.
- Witkin, S. S.; Linhares, I. M.; Giraldo, P. (2007). «Bacterial flora of the female genital tract: Function and immune regulation». Best Practice & Research Clinical Obstetrics & Gynaecology 21 (3): 347-354. PMID 17215167. doi:10.1016/j.bpobgyn.2006.12.004.
- Graver M., Wade J. (2011). «The role of acidification in the inhibition of Neisseria gonorrhoeae by vaginal lactobacilli during anaerobic growth». Ann. Clin. Microbiol. Antimicrob 10: 8. PMC 3045876. PMID 21329492. doi:10.1186/1476-0711-10-8.
- Matu M. N., Orinda G. O., Njagi E. N. M., Cohen C. R., Bukusi E. A. (2010). «In vitro inhibitory activity of human vaginal lactobacilli against pathogenic bacteria associated with bacterial vaginosis in Kenyan women». Anaerobe 16 (3): 210-215. PMID 19925874. doi:10.1016/j.anaerobe.2009.11.002.
- Skarin A., Sylwan J. (1986). «Vaginal Lactobacilli inhibiting growth of Gardnerella vaginalis, Mobiluncus and other bacterial species cultured from vaginal content of women with bacterial vaginosis». APMIS 94 (1–6): 399-403. doi:10.1111/j.1699-0463.1986.tb03074.x.
- Strus, M., M. Malinowska, and P. B. Heczko. 2002. In vitro antagonistic effect of Lactobacillus on organisms associated with bacterial vaginosis. J. Reprod. Med. 47:41–46.
- Boris S., Barbes C. (2000). «Role played by lactobacilli in controlling the population of vaginal pathogens». Microb. Infect 2 (5): 543-546. doi:10.1016/s1286-4579(00)00313-0.
- Martin R., Suarez J. E. (2010). «Biosynthesis and degradation of H2O2 by vaginal lactobacilli». Appl. Environ. Microbiol. 76: 400-405. doi:10.1128/AEM.01631-09.
- O'Hanlon D., Moench T., Cone R. (2011). «In vaginal fluid, bacteria associated with bacterial vaginosis can be suppressed with lactic acid but not hydrogen peroxide». BMC Infect. Dis. 11: 200. PMC 3161885. PMID 21771337. doi:10.1186/1471-2334-11-200.
- Aroutcheva A.; Gariti D.; Simon M.; Shott S.; Faro J.; Simoes J. A.; Gurguis A.; Faro S. (2001). «Defense factors of vaginal lactobacilli». Am. J. Obstet. Gynecol. 185 (2): 375-379. PMID 11518895. doi:10.1067/mob.2001.115867.
- Linhares I. M., Summers P. R., Larsen B., Giraldo P. C., Witkin S. S. (2011). «Contemporary perspectives on vaginal pH and lactobacilli». Am. J. Obstet. Gynecol. 204 (2): 120.e1-5. PMID 20832044. doi:10.1016/j.ajog.2010.07.010.
- Redondo-Lopez V.; Cook R. L.; Sobel J. D. (1990). «Emerging role of lactobacilli in the control and maintenance of the vaginal bacterial microflora». Rev. Infect. Dis. 12 (5): 856-872. PMID 2237129. doi:10.1093/clinids/12.5.856.
- Dahiya R. S.; Speck M. L. (1968). «Hydrogen peroxide formation by lactobacilli and its effect on Staphylococcus aureus». J. Dairy Sci. 51 (10): 1568-1572. PMID 5682478. doi:10.3168/jds.s0022-0302(68)87232-7.
- Thompson R.; Johnson A. (1951). «The inhibitory action of saliva on the diphtheria Bacillus: Hydrogen peroxide, the inhibitory agent produced by salivary streptococci». J. Infect. Dis. 88: 81-85. PMID 14803753. doi:10.1093/infdis/88.1.81.
- Wheater D. M.; Hirsch A.; Mattick A. T. R. (1952). «Possible identity of lactobacillin with hydrogen peroxide produced by lactobacilli». Nature 170 (4328): 623-624. Bibcode:1952Natur.170..623W. PMID 13002389. doi:10.1038/170623a0.
- Eschenbach D. A.; Davick P. R.; Williams B. L.; Klebanoff S. J.; Young-Smith K.; Critchlow C. M.; Holmes K. K. (1989). «Prevalence of hydrogen peroxide-producing Lactobacillus species in normal women and women with bacterial vaginosis». J. Clin. Microbiol. 27: 251-256.
- Hillier S. L.; Krohn M. A.; Klebanoff S. J.; Eschenbach D. A. (1992). «The relationship of hydrogen peroxide-producing lactobacilli to bacterial vaginosis and genital microflora in pregnant women». Obstet. Gynecol. 79 (3): 369-373. PMID 1738516. doi:10.1097/00006250-199203000-00008.
- Klebanoff SJ (1970). «Peroxidase-mediated antimicrobial activity of rat uterine fluid». Gynecol Invest 1: 21-30. doi:10.1159/000301903.
- Baeten J. M.; Hassan W. M.; Chohan V.; Richardson B. A.; Mandaliya K.; Ndinya-Achola J. O.; Jaoko W.; McClelland R. S. (2009). «Prospective study of correlates of vaginal Lactobacillus colonisation among high-risk HIV-1 seronegative women». Sex. Transm. Infect. 85 (5): 348-353. PMC 2837477. PMID 19329442. doi:10.1136/sti.2008.035451.
- Wilks M., Wiggins R., Whiley A., Hennessy E., Warwick S., Porter H., Corfield A., Millar M. (2004). «Identification and H2O2 production of vaginal lactobacilli from pregnant women at high risk of preterm birth and relation with outcome». J. Clin. Microbiol. 42 (2): 713-717. PMC 344438. PMID 14766841. doi:10.1128/jcm.42.2.713-717.2004.
- Verhelst R., Verstraelen H., Claeys G., Verschraegen G., Simaey L. Van, De Ganck C., De Backer E., Temmerman M., Vaneechoutte M. (2005). «Comparison between Gram stain and culture for the characterization of vaginal microflora: Definition of a distinct grade that resembles grade I microflora and revised categorization of grade I microflora». BMC Microbiol. 5: 61. PMC 1266370. PMID 16225680. doi:10.1186/1471-2180-5-61.
- De Backer E., Verhelst R., Verstraelen H., Alqumber M. A., Burton J. P., Tagg J. R., Temmerman M., Vaneechoutte M. (2007). «Quantitative determination by real-time PCR of four vaginal Lactobacillus species, Gardnerella vaginalis and Atopobium vaginae indicates an inverse relationship between L. gasseri and L. iners». BMC Microbiol. 7: 115. PMC 2233628. PMID 18093311. doi:10.1186/1471-2180-7-115.
- Antonio M. A.; Hawes S. E.; Hillier S. L. (1999). «The identification of vaginal Lactobacillus species and the demographic and microbiologic characteristics of women colonized by these species». J. Infect. Dis. 180 (6): 1950-1956. PMID 10558952. doi:10.1086/315109.
- Antonio M. A. D.; Rabe L. K.; Hillier S. L. (2005). «Colonization of the rectum by Lactobacillus species and decreased risk of bacterial vaginosis». J. Infect. Dis. 192 (3): 394-398. PMID 15995952. doi:10.1086/430926.
- Hawes S. E.; Hillier S. L.; Benedetti J.; Stevens C. E.; Koutsky L. A.; Wolner-Hanssen P. L.; Holmes K. K. (1996). «Hydrogen peroxide-producing lactobacilli and acquisition of vaginal infections». J. Infect. Dis. 174 (5): 1058-1063. PMID 8896509. doi:10.1093/infdis/174.5.1058.
- O'Hanlon D. E., Lanier B. R., Moench T. R., Cone R. A. (2010). «Cervicovaginal fluid and semen block the microbicidal activity of hydrogen peroxide produced by vaginal lactobacilli». BMC Infect. Dis. 10: 120. PMC 2887447. PMID 20482854. doi:10.1186/1471-2334-10-120.
- Klebanoff S. J.; Hillier S. L.; Eschenbach D. A.; Waltersdorph A. M. (1991). «Control of the microbial flora of the vagina by H202-generating lactobacilli». J. Infect. Dis. 164: 94-100. PMID 1647428. doi:10.1093/infdis/164.1.94.
- Vallor A. C.; Antonio M. A.; Hawes S. E.; Hillier S. L. (2001). «Factors associated with acquisition of, or persistent colonization by, vaginal lactobacilli: Role of hydrogen peroxide production». J. Infect. Dis. 184 (11): 1431-1436. PMID 11709785. doi:10.1086/324445.
- Ravel, Jacques; Gajer, Pawel; Abdo, Zaid; Schneider, G. Maria; Koenig, Sara S. K.; McCulle, Stacey L.; Karlebach, Shara; Gorle, Reshma et al. (15 de marzo de 2011). «Vaginal microbiome of reproductive-age women». Proceedings of the National Academy of Sciences (en inglés) 108 (Supplement 1): 4680-4687. Bibcode:2011PNAS..108.4680R. ISSN 0027-8424. PMC 3063603. PMID 20534435. doi:10.1073/pnas.1002611107. Consultado el 27 de mayo de 2016.
- Zhou X.; Bent S. J.; Schneider M. G.; Davis C. C.; Islam M. R.; Forney L. J. (2004). «Characterization of vaginal microbial communities in adult healthy women using cultivation-independent methods». Microbiology 150 (8): 2565-2573. PMID 15289553. doi:10.1099/mic.0.26905-0.
- Danielsson D.; Teigen P. K.; Moi H. (2011). «The genital econiche: Focus on microbiota and bacterial vaginosis». Ann. N. Y. Acad. Sci. 1230 (1): 48-58. Bibcode:2011NYASA1230...48D. PMID 21824165. doi:10.1111/j.1749-6632.2011.06041.x.
- Dominguez-Bello M. G.; Costello E. K.; Contreras M.; Magris M.; Hidalgo G.; Fierer N.; Knight R. (2010). «Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns». Proc. Natl. Acad. Sci. USA 107 (26): 11971-11975. Bibcode:2010PNAS..10711971D. PMC 2900693. PMID 20566857. doi:10.1073/pnas.1002601107.
- Lopes, Santos Santiago G., Cools P., Verstraelen H., Trog M., Missine G., Aila N. El, Verhelst R., Tency I., Claeys G., Temmerman M., Vaneechoutte M. (2011). «Longitudinal study of the dynamics of vaginal microflora during two consecutive menstrual cycles». PLOS ONE 6 (11): e28180. Bibcode:2011PLoSO...628180L. PMC 3227645. PMID 22140538. doi:10.1371/journal.pone.0028180.
- Zhou X.; Brown C. J.; Abdo Z.; Davis C. C.; Hansmann M. A.; Joyce P.; Foster J. A.; Forney L. J. (2007). «Differences in the composition of vaginal microbial communities found in healthy Caucasian and black women». ISME J. 1 (2): 121-133. PMID 18043622. doi:10.1038/ismej.2007.12.
- Hummelen R.; Fernandes A. D.; Macklaim J. M.; Dickson R. J.; Changalucha J.; Gloor G. B.; Reid G. (2010). «Deep sequencing of the vaginal microbiota of women with HIV». PLOS ONE 5 (8): e12078. Bibcode:2010PLoSO...512078H. PMC 2920804. PMID 20711427. doi:10.1371/journal.pone.0012078.
- Martin J. A.; Hamilton B. E.; Sutton P. D.; Ventura S. J.; Mathews T. J.; Kirmeyer S.; Osterman M. J. (2010). «Births: Final data for 2007». National Vital Statistics Reports 58: 1-85.
- Ness R. B.; Hillier S.; Richter H. E.; Soper D. E.; Stamm C.; Bass D. C.; Sweet R. L.; Rice P. (2003). «Can known risk factors explain racial differences in the occurrence of bacterial vaginosis?». J. Natl. Med. Assoc. 95: 201-212.
- Mirmonsef P.; Gilbert D.; Veazey R. S.; Wang J.; Kendrick S. R.; Spear G. T. (2012). «A comparison of lower genital tract glycogen and lactic acid levels in women and macaques: Implications for HIV and SIV susceptibility». AIDS Res. Hum. Retroviruses 28 (1): 76-81. PMC 3251838. PMID 21595610. doi:10.1089/aid.2011.0071.
- Hillier S. L.; Nugent R. P.; Eschenbach D. A.; Krohn M. A.; Gibbs R. S.; Martin D. H.; Cotch M. F.; Edelman R. et al. (1995). «Association between bacterial vaginosis and preterm delivery of a lowbirth-weight infant». N. Engl. J. Med. 333 (26): 1737-1742. PMID 7491137. doi:10.1056/nejm199512283332604.
- Clark, Natalie; Tal, Reshef; Sharma, Harsha; Segars, James (2014). «Microbiota and Pelvic Inflammatory Disease». Seminars in Reproductive Medicine 32 (1): 043-049. ISSN 1526-8004. PMC 4148456. PMID 24390920. doi:10.1055/s-0033-1361822.
- «Bacterial Vaginosis (BV): Condition Information». National Institute of Child Health and Human Development. 21 de mayo de 2013. Consultado el 3 de marzo de 2015.
- Nardis, C.; Mastromarino, P.; Mosca, L. (September 2013). «Vaginal microbiota and viral sexually transmitted diseases». Annali di Igiene 25 (5): 443-56. PMID 24048183. doi:10.7416/ai.2013.1946.
- «What are the symptoms of bacterial vaginosis?». National Institute of Child Health and Human Development. 21 de mayo de 2013. Consultado el 22 de mayo de 2016.
- Ravel, J. (2010). «Colloquium Paper: Vaginal microbiome of reproductive-age women». Proceedings of the National Academy of Sciences 108 (Supplement_1): 4680-4687. PMC 3063603. PMID 20534435. doi:10.1073/pnas.1002611107.
- Nunn, Kenetta L.; Wang, Ying-Ying; Harit, Dimple; Humphrys, Michael S.; Ma, Bing; Cone, Richard; Ravel, Jacques; Lai, Samuel K. (6 de octubre de 2015). «Enhanced Trapping of HIV-1 by Human Cervicovaginal Mucus Is Associated with Lactobacillus crispatus-Dominant Microbiota». mBio 6 (5): e01084-15. PMC 4611035. PMID 26443453. doi:10.1128/mBio.01084-15.
- Anderson, Deborah J.; Marathe, Jai; Pudney, Jeffrey (1 de junio de 2014). «The Structure of the Human Vaginal Stratum Corneum and its Role in Immune Defense». American Journal of Reproductive Immunology (en inglés) 71 (6): 618-623. ISSN 1600-0897. PMC 4024347. PMID 24661416. doi:10.1111/aji.12230.
Enlaces externos
| Control de autoridades |
|
|---|
 Datos: Q537478
Datos: Q537478 Multimedia: Vaginal flora / Q537478
Multimedia: Vaginal flora / Q537478