Migración neuronal
La migración neuronal es el proceso de la etapa embrionaria del sistema nervioso, que consiste en el desplazamiento de las neuronas desde el lugar en el que «han nacido» hasta su zona de destino final.
Características
La migración neuronal es un proceso muy complejo, y para muchas neuronas significa desplazarse una larga distancia y salvar grandes obstáculos.
Las neuronas derivadas del tubo neural siguen un mecanismo distinto de migración celular que las células derivadas de la cresta neural.[1][2][3]
Tubo neural
La mayoría de las neuronas derivadas del tubo neural migran a través de la glía radial, cuyas células sirven de soporte mecánico a las neuronas inmaduras para su desplazamiento a través del neuroepitelio. Las neuronas se desplazan por las prolongaciones gliales empleando un movimiento ameboide.
- El mecanismo de migración está controlado por moléculas de la membrana celular como por ejemplo diversas glucoproteínas que se expresan en fases tempranas del desarrollo; las llamadas moléculas de adhesión celular neurona-glía (MAC-Ng), realizan el reconocimiento de las prolongaciones de la glía radial para iniciar la migración y controlan la adhesión de las neuronas a las mismas, para permitir el desplazamiento de la neurona.[4]
Cresta neural
En el caso de las células derivadas de la cresta radial el mecanismo de migración emplea moléculas de la matriz extracelular. El inicio de la migración de estas células está determinado por tanto por la maduración de la matriz extracelular que las rodea y son las moléculas de esta matriz las que les van a marcar el camino a seguir.

Vía dorsolateral en rojo, región del cráneo.
Vía ventral en azul, región del tronco.
Para las células de la cresta radial hay dos vías de migración.
- La que siguen las células de la región craneal del embrión que migran a través de una vía dorsolateral y cuya matriz extracelular determina que se diferencien en células no neurales.
- Otra vía es la que siguen las células de la cresta neural de la región del tronco, que lo hacen por una vía ventral y cuya matriz extracelular va a hacer que se determinen como células del SNP y de la médula suprarrenal.
- Entre las moléculas de la matriz que intervienen en este proceso están las fibronectinas, que aportan lugares de adhesión a los receptores de la membrana y facilitan el desplazamiento de las células migratorias, y las lamininas en los sitios donde se produce la agregación. El balance entre estas moléculas hace que las células continúen la migración o se agrupen formando ganglios. Además se sabe que las MAC están inactivadas en las células de la cresta neural que están migrando y se activan cuando se agregan para formar ganglios.
Tipos de migración
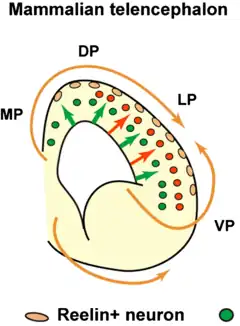
Flechas naranjas: migración tangencial.
Flechas verdes: migración radial.
MP= medial pallium; DP= dorsal pallium; LP= lateral pallium; VP= ventral pallium.
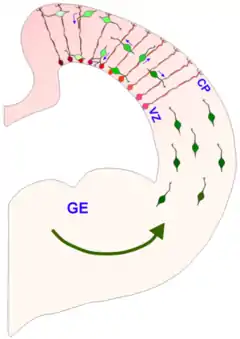
En el sistema nervioso en desarrollo, se han identificado dos modos de migración: radial y tangencial.
La migración radial es el principal modo de migración de neuronas piramidales en la corteza cerebral en desarrollo.
La migración tangencial es de las interneuronas que expresan GABA, que surgen en el telencéfalo ventral hacia la corteza.[5][6]
Migración radial en la neocorteza
Las neuronas nacen cercanas a la zona en contacto con el ventriculo cerebral del tubo neural del embrión y poco después comienzan su migración.
Al inicio de este proceso las neuronas se sitúan entre la zona ventricular y la zona marginal formando la zona intermedia que no es más que una ubicación transitoria.
Al comienzo del desarrollo cortical, las neuronas más tempranas forman la pre-placa, una capa de neuronas diferenciadas por encima de la zona ventricular.
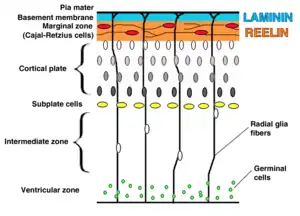
En la neocorteza en desarrollo, las células germinales (Células madre neurales) proliferan en la zona adyacente al ventrículo cerebral. Las primeras células postmitóticas que migran desde la primera capa cortical (pre-placa o pre-plate en inglés) están destinadas a convertirse en neurona de Cajal-Retzius y neuronas de la subplaca (o sub-plate).[7]
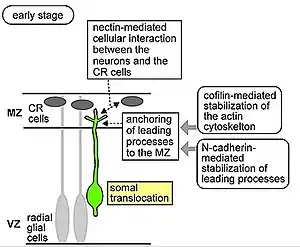
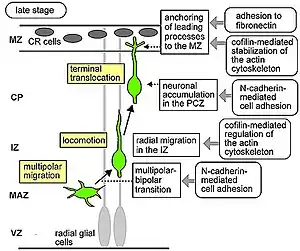
Estas células se mueven mediante «traslocación somática», las neuronas que emigran con este modo de locomoción son bipolares y se sujetan a la piamadre. El soma de estas células es transportado a la superficie de la piamadre mediante nucleocinesis, un proceso mediante el cual microtúbulos rodean el núcleo y se elongan o contraen para guiar el núcleo con su centrosoma a su posición final.[8]
Fibras radiales conocidas como glía radial (RG) puede translocarse a la placa cortical (o cortical plate) y diferenciarse en astrocitos o neuronas. La traslocación somática puede ocurrir en cualquier momento durante desarrollo.[9][10]
Posteriores olas de neuronas van a partir el preplate al migrar a lo largo de la glía radial formando la placa cortical (o cortical plate). Cada ola de células viajan a través de las capas que las preceden, de adentro hacia afuera, esto significa que las neuronas más jóvenes se van a localizar más cerca de la superficie del cerebro.[11][12] Está estimado que la migración guiada por glía representa entre un 80 y 90% de la totalidad de la población de neuronas que migran.[13]
Diferenciación neuronal
Una vez que la neurona alcanza su destino comienza su diferenciación, fase en la cual la neurona adquiere las características morfológicas y fisiológicas de neurona adulta. La conclusión a la que se ha llegado[¿quién?] sobre este tema es que aunque el patrón básico del tipo neuronal está predeterminado genéticamente, la completa diferenciación depende de las interacciones entre neuronas y de la formación de vías de conexión.
En el sistema nervioso de los vertebrados las neuronas se diferencian a partir de diferentes células progenitoras localizadas en lugares específicos en el tubo neural.
Hay una alta conservación en la especificación del destino de las células neuronales, que se da por diferentes moléculas de señalización.
- La formación de neuronas (neurogénesis)
fue dividida en 8 pasos por Goodman y Doe (1993) así:
- Inducción de las regiones neurogénicas (de formación de neuronas).
- Nacimiento y migración de las neuronas y células gliales.
- Especificación del destino celular
- Dirección del crecimiento de los conos de los axones hacia blancos específicos.
- Formación de conexiones sinápticas.
- Unión de factores trópicos para supervivencia y diferenciación.
- Reorganización competitiva de las sinapsis funcionales.
- Plasticidad sináptica continua durante la vida del organismo.
- La especificación de las neuronas
se da de manera jerárquica. Primero el epitelio del ectodermo se convierte en epitelio neuronal, posteriormente éste se convierte en células gliales o en neuronas. Después de que se ha decidido que el epitelio se convertirá en una neurona, debe decidirse qué tipo de neurona será ésta (motora, sensorial, de la comisura, etc.).
Neuronas motoras de los vertebrados
Para ilustrar los procesos de especificación Gilbert (1949) explica la especificación de las neuronas motoras de los vertebrados así:
La especificación neuronal se da por la vía Notch-Delta. La especificación del tipo neuronal parece ser controlada por la posición que tenga el precursor neuronal en el tubo neural y por su nacimiento. Las neuronas que se encuentran en el margen ventrolateral del tubo neural forman neuronas motoras, las células que se encuentran en la región dorsal del tubo neural se convierten en interneuronas. La especificación neuronal se da posiblemente en función de la posición de la célula en relación con la placa del piso (floor plate) dado que se ha observado que el injerto de células de la placa y de la notocorda las cuales secretan Sonic Hedgehog (Shh) en áreas laterales del tubo neural pueden re especificar células dorsolaterales en células motoras.
Ericson y colaboradores (1996) mostraron que las neuronas motoras se especifican por dos periodos de señalización de Shh. Un periodo temprano regulado por la secreción de Shh desde la notocorda en el que las células del margen ventrolateral se convierten en neuronas ventrales y un periodo tardío regulado por la secreción de Shh desde la placa del piso que induce a las neuronas ventrales en neuronas motoras. Por esto la proteína Sonic HedgeHog parece especificar a las neuronas motoras por la inducción de diferentes factores de transcripción a diferentes concentraciones. La posición que ocupe en el cuerpo dicha neurona motora es regulada por genes Hox y por otros genes que se encuentran en el cerebro.
Los cuerpos celulares de las neuronas motoras se agrupan en diferentes tipos de columnas longitudinales de la medula espinal. Las agrupaciones de células se dividen en columnas de Terni y columnas motoras lateral y dorsal. Las neuronas que están en lugares similares tienen blancos similares. En las extremidades posteriores del pollo, los músculos son inervados por los axones de la columna media lateral (neuronas laterales entran en la musculatura dorsal) y las neuronas motoras de la columna media inervan la musculatura ventral de las extremidades (Figura 1). Esta organización de las neuronas motoras es consistente en todos los vertebrados.
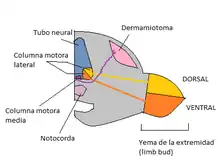
El destino celular en la médula espinal y en otros lugares del sistema nervioso central depende de dos sistemas de señalización que son activados simultáneamente junto con un sistema de inducción neuronal. Los dos sistemas de señalización se interceptan a lo largo de los ejes rostrocaudal y dorsoventral del tubo neural estableciendo así un set de redes con señales de posicionamiento específicas. El destino de las células progenitoras que se encuentran a lo largo de estos dos ejes se define por la concentración de las señales de inducción a las que están expuestas.
La señalización a lo largo del eje cabeza-cola (rostrocaudal) del tubo neural establece las subdivisiones principales del sistema nervioso central que son el prosencéfalo, el mesencéfalo, el romboencéfalo y la médula espinal.
La señalización espalda-pecho (dorsoventral) establece la diversidad de tipos celulares en las subdivisiones rostrocaudales.
Sin embargo, existen muchos otros mecanismos que permiten la generación de diferentes tipos neuronales durante el desarrollo. Por ejemplo, para la formación de los subtipos neuronales que se encuentran en la medula espinal se transmiten las señales de forma local entre las células en desarrollo.[14][15][16]
Véase también
Referencias
- «El mapa del cerebro según Pasko Rakic.». NEXciencia. Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. 2011.

- Hernández-Flórez C.E.; Beltrán M.A.; Contreras G.A. (2018). «Desarrollo neuroembriológico: el camino desde la proliferación hasta la perfección». Universitas Médica, (Pontificia Universidad Javeriana) 59 (3). Consultado el 28 de octubre de 2022.

- «Descubren nuevos mecanismos implicados en la migración neuronal». SINC Servicio de Información y Noticias Científicas. Fundación Española para la Ciencia y la Tecnología. setiembre de 2008.
- Alan R Crossman; David Neary (2019). Neuroanatomía. Texto y Atlas en Color. Elsevier Health Sciences.
- Yokota Y.; Ghashghaei HT.; Han C.; Watson H.; Campbell KJ.; Anton E. (2007). «Radial Glial Dependent and Independent Dynamics of Interneuronal Migration in the Developing Cerebral Cortex.». PLoS ONE 2 (8): e794. doi:10.1371/journal.pone.0000794.
- Hirota Y.; Nakajima K. (2017). «Control of Neuronal Migration and Aggregation by Reelin Signaling in the Developing Cerebral Cortex». Front Cell Dev Biol. (en inglés) 5 (40). doi:10.3389/fcell.2017.00040. Consultado el 30 de octubre de 2022.
- «Nucleokinesis illuminated». Nature Neuroscience 7 (11): 1169-70. November 2004. PMID 15508010. doi:10.1038/nn1104-1169. (requiere suscripción).
- «Radial glia: multi-purpose cells for vertebrate brain development.». Trends in Neurosciences 25 (5): 235-238. May 2002. PMID 11972958. doi:10.1016/S0166-2236(02)02156-2. (requiere suscripción).
- «Two modes of radial migration in early development of the cerebral cortex». Nature Neuroscience 4 (2): 143-150. February 2001. PMID 11175874. doi:10.1038/83967. (requiere suscripción).
- «Modes of neuronal migration in the developing cerebral cortex». Nature Reviews. Neuroscience 3 (6): 423-432. June 2002. PMID 12042877. doi:10.1038/nrn845. (requiere suscripción).
- «Mode of cell migration to the superficial layers of fetal monkey neocortex». The Journal of Comparative Neurology 145 (1): 61-83. May 1972. PMID 4624784. doi:10.1002/cne.901450105.
- Hatten, Mary (1999). Central Nervous System Neuronal Migration. Annual Reviews in Neuroscience.
- Garcés-Vieira M.V.; Suárez-Escudero J.C. (2014). «Neuroplasticidad: aspectos bioquímicos y neurofisiológicos». CES Med. (REVISIÓN) (Medellín: SciELO) 28 (1). Consultado el 28 de octubre de 2022.

- «Identifican un nuevo modelo de desarrollo de la corteza cerebral». CSIC. Neurociencias. 2022.
- Medina Alva M.delP.; Caro Kahn I.; Muñoz Huerta P.; Leyva Sánchez J.; Moreno Calixto J.; Vega Sánchez S.M. (2015). «Neurodesarrollo infantil: características normales y signos de alarma en el niño menor de cinco años». Rev. perú. med. exp. salud publica (Simposio) (Lima: SciELO) 32 (3). Consultado el 28 de octubre de 2022.
Bibliografía
- Purves, D. et al. "Invitación a la neurociencia". Editorial Medica Panamericana. Enero 2001, Argentina.
- Scott F. Gilbert. Developmental Biology Ninth Edition. Sinauer Associates, Inc. 2010.
- Thomas M. Jessell. Neuronal specification in the Spinal Cord: inductive signals and Transcriptional Codes. Nature (2000) Vol. 1.
| Control de autoridades |
|
|---|
 Datos: Q6013753
Datos: Q6013753