Pelagornithidae
Los pelagornítidos (Pelagornithidae), también conocidos como pseudodontornes, aves de dientes óseos, aves de dientes falsos o aves pseudodentadas, son una familia extinta de grandes aves marinas. Sus restos fósiles se han hallado en muchas partes del mundo en rocas que datan de finales del Paleoceno hasta el límite entre el Plioceno y el Pleistoceno.[2]
| Pelagornithidae | ||
|---|---|---|
| Rango temporal: 58,7 Ma - 2,5 Ma Paleoceno - Neógeno | ||
 Réplica de un esqueleto de Pelagornis miocaenus en el NMNH | ||
| Estado de conservación | ||
| Extinto en época prehistórica | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Clase: | Aves | |
| Subclase: | Neornithes | |
| Infraclase: | Neognathae | |
| Orden: |
†Odontopterygiformes Howard, 1957 | |
| Familia: |
†Pelagornithidae Fürbringer, 1888 | |
| Sinonimia | ||
Muchos de sus nombres comunes se refieren a la característica más notable de estas aves: puntas en forma de dientes en los bordes del pico, que a diferencia de los verdaderos dientes contenían conductos de Volkmann y crecían desde los huesos premaxilares y mandibulares. Incluso las especies más pequeñas de aves pseudodentadas eran del tamaño de albatros; las mayores eran verdaderamente gigantescas, y con envergaduras estimadas entre 5–6 metros estuvieron entre las mayores aves voladoras que hayan vivido. Estas aves marinas fueron dominantes en muchos océanos a través de la mayor parte del Cenozoico, y los humanos modernos aparentemente no llegaron a encontrase con estas solo por un breve espacio de tiempo evolutivo: los últimos pelagornítidos conocidos fueron contemporáneos de Homo habilis y el comienzo de la historia de la tecnología.[3]
Descripción y ecología

Aparte del gigantesco teratornítido Argentavis magnificens, las mayores aves pseudodentadas fueron las más grandes aves voladoras conocidas. Casi todas[4] las especies conocidas del Neógeno son inmensas, pero en el Paleógeno hubo algunos pelagornítidos que tenían un tamaño similar al del albatros errante (género Diomedea) o incluso algo más pequeñas. Una especie sin describir, provisionalmente llamada "Odontoptila inexpectata"[5] – del límite Paleoceno-Eoceno de Marruecos – es la menor ave pseudodentada descubierta a la fecha y era solo un poco mayor que un petrel de barba blanca (Procellaria aequinoctialis).[6]
Los Pelagornithidae tenían huesos con paredes extremadamente delgadas ampliamente pneumatizadas con extensiones de los sacos aéreos de los pulmones. Muchos huesos fósiles de los miembros están aplastados por esta razón. En vida, los delgados huesos y la extensa neumatización les permitía a estas aves alcanzar un gran tamaño mientras retenían una carga alar crítica. Aunque se considera que 25 kg/m² (5 lb/ft2) es la máxima carga alar que puede haber para un ave que haga vuelo propulsado, hay evidencia de estas aves usaban el planeo dinámico de forma casi exclusiva: el extremo proximal del húmero tenía una forma de diagonal alargada que difícilmente pudo haber permitido el movimiento necesario para el típico vuelo con aleteo de las aves; su peso por tanto no es fácil de estimar. Las posiciones de sujeción de los músculos responsables de sostener el brazo extendido de forma recta estaban particularmente bien desarrollados, y en conjunto toda la anatomía parece permitir la habilidad de sostener las alas de manera rígida en la articulación glenoide de una manera sin paralelo en ninguna otra ave conocida. Esto es especialmente prominente en los pelargornítidos del Neógeno, y menos desarrollado en las formas más antiguas del Paleógeno. El esternón tenía una forma alta y corta típica de los planeadores dinámicos, y extensiones óseas en el margen anterior de la quilla del esternón seguramente anclaban la fúrcula.[7]
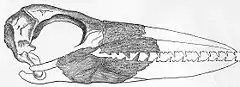
Las patas eran proporcionalmente cortas, los pies probablemente tenían una membrana y el hallux era vestigial o enteramente ausente; los tarsometatarsos (huesos del tobillo) se parecían a los de los albatros aunque la disposición de los pies era similar a la de los fulmares. Típico de las aves pseudodentadas era un segundo dedo que se sujetaba hacia la rodilla y estaba extendido hacia afuera con respecto a los demás. Los "dientes" estaban probablemente cubiertos por la ranfoteca en vida, y había dos surcos corriendo a lo largo del envés del pico superior justo dentro de los bordes en los que estaban los "dientes". Por tanto, cuando el pico se cerraba solo los "dientes" superiores eran visibles, mientras que los inferiores se ocultaban detrás de estos. Dentro de las cuencas oculares de al menos algunas aves pseudodentadas – quizás solo en las especies más recientes – había glándulas salinas bien desarrolladas.[8]
La mayor parte del cuerpo de los pelagornítidos no es conocido de un esqueleto bien preservado asociado y mucho del mejor material preservado consiste de un único hueso; por otra parte la extensa presencia y el gran tamaño hace que haya algunos restos de aves individuales bastante completos (aunque mayormente aplastados y distorsionados) que fueron sepultados mientras estaban muertas, con algunas plumas fosilizadas. Grandes secciones de cráneo y algunas piezas del pico se hallan con cierta frecuencia. En febrero de 2009, un cráneo fosilizado casi completo de un presunto Odontopteryx del límite Chasiquense-Huayqueriense hace cerca de 9 millones de años (Ma) fue dado a conocer en Lima. Había sido hallado unos pocos meses antes en el distrito de Ocucaje de la provincia de Ica (Perú). De acuerdo al paleontólogo Mario Urbina, quien descubrió el espécimen, y sus colegas Rodolfo Salas, Ken Campbell y Daniel T. Ksepka, el cráneo de Ocucaje es uno de los cráneos de pelagornítidos mejor preservados que hayan hallado.[9]
Ecología y extinción

A diferencia de los dientes verdaderos de aves mesozoicas como Archaeopteryx o Aberratiodontus, los pseudodientes de los Pelagornithidae no parecen haber sido aserrados o tener alguna clase de borde cortante especializado, siendo útiles para sostener presas escurridizas para tragarlas enteras más que para cortar trozos de estas. Dado que los dientes era huecos o estaban llenos de hueso esponjoso y se desgastaban o rompían con facilidad en los fósiles, se conjetura que no eran especialmente resistentes en vida. Las presas de los pelagornítidos debieron ser presas de cuerpos blandos, y habría abarcado principalmente cefalópodos[10] y peces de piel blanda.[11] Sus presas pueden haber alcanzado un tamaño considerable. Aunque algunas reconstrucciones muestran a los pelagornítidos como aves buceadoras a la manera de los alcatraces, las delgadas paredes de los huesos altamente neumatizados se deben haber fracturado fácilmente juzgando por el estado de muchos de los fósiles, lo cual lo hace un método de alimentación improbable, si no totalmente peligroso. En vez de ello, las presas pudieron haber sido capturadas desde arriba de la superficie del océano mientras las aves lo sobrevolaban o nadaban, y estas probablemente sumergían solo el pico en la mayoría de ocasiones. Su articulación del hueso cuadrado con la mandíbula inferior se parecía a la de los pelícanos u otras aves que pueden abrir su pico ampliamente. Las aves pseudodentadas pueden haber llenado un nicho ecológico casi idéntico al de los mayores pterosaurios piscívoros, cuya extinción al final del Cretácico puede haber despejado el camino para el exitoso dominio de 50 millones de años de los Pelagornithidae. Como los actuales albatros (pero quizás de forma diferente a los pterosaurios en los distintos climas y placas tectónicas del Mesozoico), las aves pseudodentadas pudieron haber usado el sistema de corrientes oceánicas y la circulación atmosférica para tomar varias vías planeando de ida y vuelta sobre los océanos abiertos, regresando a aparearse solo cada pocos años. A diferencia de los albatros, los cuales evitan las corrientes ecuatoriales tropicales con sus doldrums, los Pelagornithidae han sido hallados en todo tipo de climas, y los registros de hace 40 Ma se extienden desde Bélgica pasando por Togo hasta la Antártida. Es notable que los pingüinos y los plotoptéridos – ambos grupos de aves buceadoras que se alimentaban sobre la plataforma continental – son casi invariablemente halladas en compañía de las aves pseudodentadas. Por tanto, estas parecen haberse reunido en regiones con surgencias, presumiblemente para alimentarse para quizás reproducirse en las cercanías.[12]

Se ha afirmado en ocasiones que así como otras aves marinas (como por ejemplo los no voladores plotoptéridos), la radiación evolutiva de los cetáceos y los pinípedos superó a las aves pseudodentadas y las llevó a su extinción. Aunque esto puede ser cierto para los plotoptéridos, para los pelagornítidos esto no es muy probable por dos razones: Primero, los Pelagornithidae continuaron evolucionando por 10 millones de años después de que los misticetoss evolucionaran, y a mediados del Mioceno Pelagornis coexistió con Aglaocetus y con Eobalaenoptera harrisoni más allá de la costa oriental del Atlántico, mientras que en el Pacífico Osteodontornis habitó en los mismos mares que Balaenula y Morenocetus; el pequeño género ancestral de cachalote Aulophyseter (y/o Orycterocetus) aparecen en los océanos del Hemisferio Norte de esa época, mientras que el cachalote mediano Brygmophyseter recorría el Pacífico Norte. En lo que concierne a los pinípedos del Mioceno, una amplia diversidad de morsas antiguas[13] y leones marinos como Thalassoleon habitaban el noreste, mientras que la antigua foca leopardo Acrophoca es una especie notable del Pacífico sureste. Por otra parte, los pinípedos están limitados a aguas cercanas a las costas mientras que las aves pseudodentadas vivían en los mares abiertos, al igual que los grandes cetáceos, y como todos los grandes carnívoros estos tres grupos eran estrategas de la K con densidades de población moderadas o muy bajas.[14]
Por tanto, la competencia directa por comida entre estas aves y los cetáceos o los pinípedos no pudo haber sido muy severa.Tanto las aves como los pinípedos podrían necesitar terrenos cerca al mar planos para tener a sus crías, y la competencia puede haber tenido un impacto en la población de aves. Las especializaciones para el planeo dinámico pudo haber de hecho restringido los posibles sitios de anidación para las aves, pero por otro lado las tierras altas o en sierras litorales pudieron haberle provisto territorios de cría a los Pelagornithidae que eran inaccesibles para los pinípedos; de la misma manera que muchos albatros hoy en día anidan en las tierras altas d islas (por ej. las Galápagos o Torishima). Las aves de dientes óseos probablemente requerían fuertes corrientes ascendentes para despegar y pudieron haber preferido en cualquier caso sitios altos por esta razón, lo que hace mínima a la competencia con las colonias de pinípedos. En cuanto a terrenos de cría, fragmentos de cáscara de huevos gigantes de las montañas Famara en Lanzarote (Islas Canarias) fueron tentativamente atribuidos a aves pseudodentadas del Mioceno tardío. En la Arcilla de Londres del Ypresiense de la Isla de Sheppey, en donde los fósiles de pelagornítidas son hallados con frecuencia, estos estaban depositados en un mar epicontinental poco profundo durante el Máximo termal del Paleoceno-Eoceno con altos niveles de mar. Los presuntos sitios de anidación no pueden haber estado muy alejados de la costa como lo son muchas colonias de aves marinas de hoy, ya que la región estaba encerrada entre los Alpes y los Montes Grampianos y los Alpes escandinavos, en un mar menos ancho que el actual mar Caribe. Las aves pseudodentadas del Neógeno son comunes a lo largo de las costas americanas cerca de los Montes Apalaches y montañas de las cordillerass, y estas especies presumiblemente anidaban no muy lejos de la costa o incluso en las propias montañas. A este respecto la presencia de hueso medular en los especímenes de Lee Creek Mine en Carolina del Norte (EE.UU.) es notable, y entre las aves esto generalmente se halla en hembras grávidas, indicando que los terrenos de anidación no se encontraban muy lejos. Por otra parte, en las islas de origen volcánico del océano Pacífico la erosión de los últimos millones de años destruyó cualquier resto de las colonias de anidación de los pelagornítidos que puedan haber existido alguna vez en el océano abierto. Por ejemplo, la Isla Necker era de considerable tamaño hace diez millones de años, cuando Osteodontornis recorría el Pacífico.[15]

No hay entonces una sola razón obvia para la extinción de las aves pseudodentadas. Un escenario de cambio ecológico generalizado – exacerbado por el inicio de las eras del hielo y los cambios en las corrientes oceánicas debido a la deriva continental (como la aparición de la Corriente Circumpolar Antártica o el cierre del istmo de Panamá) – es más probable, siendo las pelagornítidas remanentes de una fauna mundial del Paleógeno que estaba fallando en adaptarse. En este sentido puede ser significativo que algunos linajes de cetáceos, como los primitivos delfines de la familia Kentriodontidae o los Squalodontidae, florecieron junto con los Pelagornithidae y terminaron extinguiéndose en una época cercana. De la misma manera, la actual diversidad de géneros de pinípedos y cetáceos evolucionó mayormente durante el límite Mioceno-Plioceno, lo que sugiere que muchos nichos ecológicos aparecieron o quedaron vacantes. Adicionalmente, lo que fuera que causara la disrupción del Mioceno Medio y la crisis salina del Mesiniense afectó no poco la cadena trófica de los océanos de la Tierra, y este último evento llevó a una amplia extinción de aves marinas. En conjunto, estos factores llevaron a que los animales del Neógeno finalmente reemplazaran a los últimos remanentes de la fauna paleógena en el Plioceno. De hecho, resulta conspicuo el hecho de que las aves pseudodentadas antiguas son típicamente halladas en los mismos depósitos que los plotoptéridos y los pingüinos, mientras que las formas más recientes son simpátricas con alcas, albatros, pingüinos y los Procellariidae – los cuales, sin embargo, experimentaron una radiación adaptativa de considerable extensión coincidente con la caída de la cadena trófica de tipo Paleógeno (y probablemente causada por esta). Aunque el registro fósil es siempre incompleto, parece que los cormoranes y gaviotas son muy raramente hallados en asociación con los Pelagornithidae.[16]
.jpg.webp)
Independientemente de la causa de su extinción, durante el largo periodo de su existencia estas aves se volvieron presas de otros grandes depredadores. Pocas de las aves que coexistieron con estas eran lo bastante grandes como para dañarlas mientras volaban; como es evidenciado por Limnofregata del Eoceno Inferior, las fragatas coevolucionaron con los Pelagornithidae y pueden haber hostigado a alguna de las especies pequeñas por comida en ocasiones, como hoy en día roban a los albatros. En los estratos del Mioceno medio o principios del Plioceno de Lee Creek Mine, se han hallado restos de aves pseudodentadas que probablemente cayeron víctimas de tiburones mientras se alimentaban. Los mayores miembros de la abundante fauna de tiburones de Lee Creek Mine que cazaban cerca de la superficie del agua incluyen al tiburón vaca de hocico corto (Notorynchus cepedianus), los tiburones de arena Carcharias, los tiburones mako Isurus y Cosmopolitodus, los tiburones blancos Carcharodon.[17] Otro tiburones incluyen al gáleo alargado Hemipristis serra, tiburones tigre (Galeocerdo), tiburones balleneros Carcharhinus, el tiburón limón (Negaprion brevirostris) y tiburones martillos (Sphyrna), y quizás (dependiendo de la edad de los fósiles de las aves) también el pez sierra Pristis, los tiburones tigre de arena Odontaspis, y los tiburones macarela Lamna y Parotodus benedeni. Es notable que los fósiles de aves buceadoras más pequeñas – como alcas, somorgujos y cormoranes – así como los de albatros son hallados mucho más comúnmente en egagrópilas de tiburones que de aves pseudodentadas, apoyando la idea de que estas últimas tenían bajas densidades de población y capturaban la mayor parte de su comida en pleno vuelo.[18]
Apariencia externa



No se sabe nada certero acerca de la coloración de estas aves, ya que no dejaron descendientes vivos. Pero se pueden hacer algunas inferencias basándose en su filogenia: si eran miembros del grupo de "aves acuáticas superiores" (ver más adelante), entonces tendrían un plumaje similar al representado en la reconstrucción de Osteodontornis orri – los Procellariiformes y Pelecaniformes en el moderno sentido (o Ciconiiformes, si los Pelecaniformes son incluidos en estos) difícilmente carotenoides o colores estructurales en su plumaje, y generalmente carecen incluso de feomelaninas. Por tanto, los únicos colores comúnmente hallados en estas aves son negro, blanco y varios tonos de gris. Algunos tienen parches de plumas iridiscentes, o matices pardos o rojizos, pero estos son raros y limitados en extensión, y aquellas especies en las cuales son hallados (por ej. avetoros, los ibis o el ave martillo) son usualmente encontradas en hábitats de agua dulce.[19]
Si las aves pseudodentadas eran miembros de Galloanserae, las feomelaninas pueden haber sido más probables de encontrar en sus plumas, pero es notable que los linajes más basales de Anseriformes típicamente son de color gris y negro o blanco y negro. Entre las aves oceánicas en general, la parte superior tiende a ser mucho más oscura que la inferior (incluyendo las alas) – aunque algunos petreles son totalmente de gris oscuro, una combinación de un gris más o menos oscuro en la parte superior y un blanco en la inferior y (usualmente) la cabeza es de una coloración ampliamente hallada en las aves marinas y puede haber sido plesiomórfica para las "aves acuáticas superiores" o, quizás más probablemente, ser una adaptación para proveer de camuflaje, en particular para evitar ser destinguida contra el cielo por sus presas en el mar. Es notable que al menos las plumas primarias rémiges, así como otras plumas de vuelo, son generalmente de color negro en muchas aves; incluso si el resto del plumaje es completamente blanco, como en algunos pelícanos o el mainá de Rothschild (Leucopsar rothschildi). Esto se debe al hecho de que las melaninas polimerizan las plumas, haciendo a las plumas oscuras muy robustas; dadas las grandes fuerzas a las que se exponen las plumas de aves en el vuelo, la gran cantidad de melanina les da una mejor resistencia contra el daño por el uso. En las aves planeadoras dependientes de los fuertes vientos como lo eran las pelagornítidas, las puntas de las alas y tal vez de las colas serían negras.[20][19]
En lo que concierne a las partes descubiertas, todos los presuntos parientes de Pelagornithidae muy frecuentemente tienen colores rojizos brillantes, en particular en el pico. La falta de cereza sobre sus relaciones filogenéticas que rodean a estas aves no permiten inferir si poseían un saco gular similar al de los pelícanos. Si lo tenían, este probablemente era anaranjado o rojo, y puede haber sido usado en comportamientos de exhibición. El dimorfismo sexual era posiblemente casi inexistente, como es típico entre los Anseriformes basales y las "aves acuáticas superiores".[19]
Taxonomía, sistemática y evolución

El nombre "pseudodontornes" se refiere al género Pseudodontornis, la cual sirvió por algún tiempo como denominación de la familia. Sin embargo, el nombre en uso en el presente Pelagornithidae antecede Pseudodontornithidae, y por tanto los autores modernos generalmente prefieren usar "pelagornítidos" sobre "pseudodontornes". Este último nombre es generalmente usado en la literatura de mediados del siglo XX.[21]
Históricamente, los dispares huesos de las aves pseudodentadas de las aves pseudodentadas estaban distribuidos en seis grupos: algunos géneros descritos a partir de huesos de patas fueron situados en la familia Cyphornithidae, y eran considerados parientes cercanos de la familia de los pelícanos (Pelicanidae). Estos fueron unidos más tarde en la superfamilia Pelecanides en el suborden Pelecanae, o más tarde en (después de que los sufijos de las categorías taxonómicas fueran fijados a los estándares actuales) Pelecanoidea en el suborden Pelecani. Subsecuentemente, algunos los incluyeron en la falsa "familia" "Cladornithidae" en un "suborden" de pelecaniformes "Cladornithes". Los géneros conocidos de material craneano fueron típicamente asignados a una o dos familias (Odontopterygidae y a veces también Pseudodontornithidae) en un suborden pelecaniforme Odontopteryges o Odontopterygia. Mientras tanto, Pelagornis, descrito de huesos del ala, fue tradicionalmente situado en una familia pelecaniforme monotípica, Pelagornithidae. Esta fue frecuentemente asignada tanto al suborden de los alcatraces y cormoranes Sulae (el cual fue anteriormente tratado como la superfamilia Sulides en el suborden Pelecanae), o a los Odontopterygia. El esternón de Gigantornis fue situado en la familia de los albatros (Diomedeidae) en el orden de las aves marinas con nariz tubular (Procellariiformes).[22]
La mayor confusión taxonómica y sistemática ha afectado a Dasornis. Este género fue establecido basándose en una pieza grande craneal, la cual por largo tiempo fue situada en la familia Gastornithidae debido meramente a su tamaño. Hoy en día se reconoce que Argillornis pertenece a Dasornis; este fue descrito de huesos del ala, y generalmente incluido en Sulae como parte de "Elopterygidae", otra "familia" sin validez, y su género tipo no es considerado como un ave de tipo moderno por los autores actuales. Algunos fragmentos óseos adicionales del tarsometatarso (tobillo) fueron situados en el género Neptuniavis y asignados a Procellariidae en los Procellariiformes. Apenas en 2008 se mostró que todos estos restos pertenecían al género de ave pseudodentada Dasornis.[23]
Sistemática y filogenia
La sistemática de las aves de dientes óseos ha sido sujeto de considerable debate. Inicialmente, estos fueron relacionados con los (por entonces parafiléticos) "Pelecaniformes" (pelícanos y sus presuntos parientes, como el alcatraz y las fragatas) y los Procellariiformes (aves de nariz de tubo como los albatros y petreles), debido a su anatomía general similar. Algunos de los primeros restos del enorme Dasornis fueron tomados por los de una ratites y más tarde como un gastornítido. Estos restos fueron incluso usados para sugerir una relación cercana entre estos dos grupos – y ciertamente, los pelícanos y los procelariformes, así como los otros "Pelecaniformes" (cormoranes y parientes) los cuales son preferiblemente separados hoy en día en los Phalacrocoraciformes, los Ciconiiformes (cigüeñas y/o garzas e ibis o los Pelecaniformes "corona") y los Gaviiformes (colimbos/zampullines) parece conformar una radiación, posiblemente un clado, de "aves acuáticos superiores". Sin embargo, los Pelagornithidae ya no son más considerados como el eslabón perdido entre pelícanos y albatros, sino como mucho cercanos a los primeros y solo convergentes con los últimos en ecomorfología.[24]
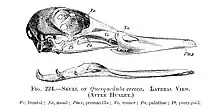

Notése también las impresiones de la glándula salina supraorbital de los Charadriiformes.
En 2005, un análisis cladístico propuso una cercana relación entre las aves pseudodentadas y los Anseriformes. Estas no son parte de las "aves acuáticas superiores" sino de Galloanserae, un linaje basal de las aves neognatas. Algunas características, mayormente del cráneo, apoyan esta hipótesis. Por ejemplo, los pelagornítidos carecen de una cresta sobre el lado inferior del hueso palatino, mientras que en las Neoaves – el clado hermano de los Galloanserae que incluye a las "aves acuáticas superiores" y las "aves terrestres superiores" – tienen esa cresta. También, como en los patos, gansos y cisnes los pelagornítidos solo tienen dos cóndilos en el proceso mandibular del hueso cuadrado, con el cóndilo medio hacia atrás del cóndilo lateral. Su articulación del basipterigoideo es similar a la de los Galloanseres. En el lado de la lámina del parasfenoides, hay una amplia plataforma como en los Anseriformes. El punto de sujeción de la parte coronoidal del músculo aductor externo de la mandíbula de las aves de dientes óseos estaba localizado en la línea media, el proceso rostropterigoide tenía un apoyo en su base y el hueso mesetmoide tenía una profunda depresión en la concha caudal, como en los Gallonaserae.[25]
En lo que concieren a otras partes del esqueleto, las sinapomorfias propuestas de los pelagornítidos y los Galloanserae son hallados mayormente en los brazos y huesos de la mano: la ulna tienen una parte superior posterior fuertemente convexa en el extremo del codo – en el extremo de la mano en el que los músculos escapulotricipitales se sujetan –, un cótilo dorsal acabado en punta y solo una leve depresión para alojar el menisco entre la ulna y el radio; hacia el codo, el surco intercondilar de la ulna se vuelve amplio y limita por un largo margen devanado en la zona ventral. El radio, sin embargo, tiene un borde convexo ventral en el cótilo humeral, el cual continua prominentemente al borde interno de la protuberancia donde el músculo biceps brachii se sujeta; hacia la parte superior del hueso radio la superficie se vuelve aplastada y triangular hacia la superficie articular de la ulna. El carpometacarpo tanto de los Anseriformes como de las aves pseudodentadas tienen un prominente proceso pisiforme, el cual se extiende desde la tróclea carpal lejos de los dedos a lo largo del lado delantero del hueso. Sobre el lado bajo del carpometacarpo, donde hay una larga pero estrecha sínfisis de los metacarpos distales, con el hueso metacarpal grande con un borde medio en el hueso que tiene un borde dorsal se curva exteriormente hacia el extremo del mismo, y la articulación del pulgar tiene una protuberancia bien desarrollada en la parte trasera de su superficie articular. Las patas y los huesos del pie, como es de esperarse en estas aves no son tan especializados para nadar como lo son los de las aves acuáticas, y por tanto muestran menos similitudes entre Anseriformes y las aves pseudodentadas: en el tibiotarso hay una incisión amplia entre los cóndilos y el centro del cóndilo es más estrecho que el cóndilo lateral y hacia delante sobresale, el tarsometatarso tiene un foramen bajo distal vascular con la apertura empotrada en su superficie plantar y una tróclea en el dedo medio que es alargada, ligeramente oblicua, proyectándose en la parte inferior del pie y formando una punta en el extremo.[26]
No es claro si estas apomorfias realmente unen a los Anseriformes y las aves de dientes óseos, pero por otra parte, el esternón, la parte distal del húmero y los huesos de las patas de los pelagornítidos parecen mostrar apomorfias típicas de las "aves acuáticas superiores". Mientras que los detalles de los huesos del neurocráneo son muy informativos para la clasificación filogenéticamente, los rasgos del cráneo en que ambos grupos son similares se relacionan generalmente al punto en donde el pico se sujeta al cráneo, y por tanto pueden estar sujetas a las fuerzas selectivas producto de arrebatar a sus presas desde la capa superior de agua. Las aparentes características no neoavianas que distinguen a los pelagornítidos podrían ser solo plesiomorfias retenidas o atavismos; ya que las "aves acuáticas superiores" son Neoaves muy antiguas y ninguno de los supuestos miembros basales de su radiación (véase también "Graculavidae") fueron incluidos en el análisis, no se sabe con certeza cuando las condiciones derivadas típicas de las Neoaves modernas fueron adquiridas. Los rasgos de los huesos del pie son notoriamente propensos a ser afectados por las fuerzas de selección en las aves, por tanto la evolución convergente puede inhibir o incluso invalidar los análisis cladísticos; sin embargo, las aparentes autopomorfias del antebrazo y los huesos de la mano son difíciles de explicar como no sea por una relación evolutiva real. La localización de las glándulas salinas dentro de las órbitas oculares de Osteodontornis y Pelagornis (y probablemente otros géneros) muestra que cualquiera que fueran sus relaciones fiologenéticas, los pelagornítidos estaban adaptados al hábitat oceánico de forma independiente de los pingüinos y los procelariformes, los cuales por su parte tienen glándulas salinas supraorbitales. El hallux vestigial o no existente– como en los patos, pero a diferencia de los pelícanos los cuales tienen cuatro dedos totalmente desarrollados y con membrana – fue una característica usada para negar una relación cercana con los pelícanos. Pero como se sabe actualmente, los pelícanos son más cercanos a las cigüeñas (las cuales tienen un hallux pero no membranas) que a las aves pseudodentadas y evolucionaron dedos del pie con membranas independientemente. Siendo el hallux hipertrofiado y con membrana tanto apomórfico y parafilético, su ausencia en las aves pseudodentadas no provee mucha información sobre su parentesco.[27]
Aunque los Galloanserae gigantes fueron comunes y diversos en el Paleógeno en particular, estas (como Gastornis y Dromornis) eran aves terrestres no voladoras; esto quizás es significativo dado que las únicas otras ave con "dientes óseos" conocidas son las dos especies del género de moa-nalo Thambetochen, grandes patos no voladores de las Islas Hawaianas. En cualquier caso, el análisis cladístico de 2005 usó una muestra representativa de los Procellariiformes y recuperó un clado bien soportado que respalda el actual consenso. La presunta relación cercana entre los pelagornítidos y los procelariformes puede ser descartada después de todo. En lo que concierne a los "Pelecaniformes", los análisis no recuperaron la filogenia correcta y no incluyó al picozapato (Balaeniceps rex, un "eslabón perdido" entre los pelícanos y cigüeñas); claramente, la radiación adaptativa del linaje de pelícanos-cigüeñas induce a error al análisis. Adicionalmente, los Galloanserae no fueron recuperados como monofiléticos. En 2007, un análisis cladístico mucho más extenso de la anatomía de las aves incluyendo algunas formas fósiles (aunque no los cruciales[28] taxones del Cretácico Superior, los cuales son usualmente conocidos solamente de restos fragmentarios) resolvió algo mejor la radiación de las "aves acuáticas superiores"; aun así, el problema de los rasgos de la pata y el pie llevando a confusiones en el análisis sigue siendo notable.[29]
Ya que las relaciones de estas aves tanto con las Galloanserae como las "aves acuáticas superiores", son por ahora situadas en un orden distinto, Odontopterygiformes a manera de compromiso, en vez de ser el suborden Odontopterygia de los pelecaniformes/ciconiiformes o de los anseriformes, o incluso una familia de los Anseriformes, Ciconiiformes o Pelecaniformes. En cualquier caso dicha clasificación es difícilmente errónea, ya que las aves pseudodentadas son distintas tanto de los Presbyornithidae como los Scopidae, hoy en día generalmente considerados como las ramas más basales de los Anseriformes y el grupo de los pelícanos y cigüeñas, respectivamente. Esto también deja la opción de separar a los Pelagornithidae en varias familias apropiadamente. Es interesante notar que cuando Boris Spulski estableció a los Odontopterygia en 1910, él lo hizo en parte porque él notó algunas similitudes entre las aves pseudodentadas y las Galloanserae ya mencionadas antes. Dasornis fue durante un buen tiempo confundido con un Gastornithiformes, que ahora son considerados como muy cercanos a los Anseriformes. Asimismo, el pelagornítido Palaeochenoides mioceanus fue inicialmente considerado como un anseriforme, y lo mismo podría ser cierto para el supuesto cisne del Oligoceno Guguschia nailiae. Sin embargo, en el caso anterior, un "mucho más que convincente"[30] análisis para una clasificación externa a los Galloanserae fue publicada ya al año después de su descripción. Muchos huesos no reconocidos de pelagornítidos fueron inicialmente asignados a las familias de "aves acuáticas superiores", típicamente al (por entonces parafilético) "Pelecaniformes", pero en particular el tarsometatarso fue considerado como de un procelariiforme. Los Odontopterygiformes fueron establecidos originalmente cuando Osteodontornisfue descrito a partir de uno de los pocos esqueletos razonablemente completos de estas aves. Hildegarde Howard encontró que, sin importar que algunas de sus características se parecieran a las de otras aves, su combinación era muy distinta a la de cualquier otra neognata conocida.[31]
Géneros y especímenes sin identificar
Debido al estado fragmentario y aplastado de muchos de los restos de aves pseudodentadas, no es claro si la docena de géneros que han sido nombrados son todos válidos. Solo los picos son lo bastante robustos y distintivos como para permitir una buena delimitación taxonómica, e incluso estos son usualmente hallados como piezas rotas. Por ejemplo, se ha descubierto recientemente que Argilliornis y Neptuniavis son los huesos del brazo y de las patas, respectivamente, de Dasornis, que hasta entonces solo era conocido de huesos craneales. Se considera usualmente que el tamaño es un marcador confiable de la diversidad genérica, pero teniendo cuidado de determinar si los especímenes menores no proceden de aves jóvenes.[32]
Tentativamente, los siguientes géneros son reconocidos:[33]
- Pseudodontornis (Paleoceno Superior ?–? Oligoceno Superior de Charleston, Carolina del Sur, EE.UU.) – ¿polifilético (la especie tipo en Palaeochenoides/Pelagornis)?
- "Odontoptila" (Paleoceno Superior/Eoceno Inferior de la Cuenca Ouled Abdoun, Marruecos) – un nomen nudum; preocupado por Odontoptila[34]
- Odontopteryx (Paleoceno Superior/Eoceno Inferior de la Cuenca Ouled Abdoun, Marruecos – Eoceno Medio de Uzbekistán) – incluyendo "Neptuniavis" minor, puede incluir "Pseudodontornis" longidentata, "P." tschulensis y Macrodontopteryx
- Dasornis (Eoceno Inferior de la Arcilla de Londres de la Isla de Sheppey, Inglaterra) – incluyendo Argillornis, "Lithornis" emuinus y "Neptuniavis" miranda; puede incluir "Odontopteryx gigas" (un nomen nudum), "Pseudodontornis" longidentata y Gigantornis
- Macrodontopteryx (Eoceno Inferior de la Arcilla de Londres en Inglaterra) – puede incluir a "Pseudodontornis" longidentata y/o pertenecer a Odontopteryx
- cf. Odontopteryx (Eoceno Inferior de Virginia, EE.UU.)[35]
- Gigantornis (Eoceno Medio de Ameki, Nigeria) – puede pertenecer a Dasornis
- cf. Odontopteryx (Eoceno Medio de México)[36]
- Pelagornithidae gen. et sp. indet. (Eoceno Medio del Monte Discovery, Antártida) – es el mismo gran espécimen de la Isla Seymour/Dasornis/Gigantornis?[37]
- Pelagornithidae gen. et sp. indet. (Eoceno Medio de Etterbeek, Bélgica) – Dasornis/Macrodontopteryx?[38]
- "Aequornis" (Eoceno Medio de Kpogamé-Hahotoé, Togo) – un nomen nudum[39]
- Pelagornithidae gen. et spp. indet. (Eoceno Medio/Superior de La Meseta en la Isla Seymour, Antártida) – ¿dos especies? ¿corresponde al espécimen del Monte Discovery /Dasornis/Gigantornis, Odontopteryx?[40]
- Pelagornithidae gen. et sp. indet. (Eoceno Superior de Francia)[41]
- Pelagornithidae gen. et sp. indet. (Eoceno Superior de Kazajistán) – puede pertenecer a Zheroia[42]
- Pelagornithidae gen. et sp. indet. (Eocen de las Islas Shetland del Sur, Atlántico Sur)[43]
- cf. Dasornis[44] (Eoceno Superior/Oligoceno Inferior de Oregon, EE.UU.) – Cyphornis?[45]
- cf. Macrodontopteryx (Oligoceno Inferior de Hamstead, Inglaterra) – puede pertenecer a Proceriavis[46]
- Pelagornithidae gen. et sp. indet. (Oligoceno Inferior de Japón)[47]
- Caspiodontornis (Oligoceno Superior de Pirəkəşkül, Azerbaiyán) – puede pertenecer a Guguschia
- Palaeochenoides (Oligoceno Superior de Carolina del Sur, EE.UU.) – puede incluir a Pseudodontornis longirostris o pertenecer a Pelagornis
- Pelagornithidae gen. et sp. indet. (Oligoceno Superior de Carolina del Sur, EE.UU.)[48]
- Pelagornithidae gen. et sp. indet. (Yamaga Oligoceno Superior de Kitakyushu, Japón) – Osteodontornis?[49]
- Tympanonesiotes (Oligoceno Superior o Mioceno Inferior del Río Cooper, EE.UU.)
- Cyphornis (Mioceno Inferior de Carmanah Point, Isla de Vancouver, Canadá) – puede incluir a Osteodontornis
- Osteodontornis (Mioceno Inferior – Plioceno Inferior) – puede pertenecer a Cyphornis
- Pelagornis (Mioceno Inferior de Armagnac, Francia – Pleistoceno Inferior de Ahl al Oughlam, Marruecos) – puede incluir a Pseudodontornis longirostris, Palaeochenoides
- Pelagornithidae gen. et spp. indet. (¿Mioceno Inferior? – Plioceno Inferior de EE.UU.) – ¿2-3 especies? Pelagornis?[50]
- cf. Osteodontornis (Mioceno Medio de Capadare en la Cueva del Zumbador, Venezuela)[51]
- cf. Osteodontornis/Pelagornis (¿Mioceno Medio/Superior? de North Canterbury, Nueva Zelanda)[52]
- cf. Pelagornis (Mioceno Medio de Bahía Inglesa en Chile – Plioceno Inferior de Chile y Perú) – ¿2 especies?[53]
- cf. Osteodontornis (Mioceno Medio de Pisco – Plioceno Inferior de Perú) – ¿2 especies?[54]
- "Pseudodontornis" stirtoni (Mioceno o Plioceno de la Playa Motunau, Nueva Zelanda) – a veces Neodontornis
- Pelagornithidae gen. et sp. indet. (Plioceno Inferior de Yushima en Maesawa, Japón) – Osteodontornis?[55]
- cf. "Pseudodontornis" stirtoni (Plioceno Medio de la Lodolita Tangahoe en Hawera, Nueva Zelanda)[56]
- Pelagornithidae gen. et sp. indet. (Pleistoceno Inferior de Dainichi en Kakegawa, Japón) – ¿Osteodontornis?[57]
- Pelagornis sp. (Plioceno de California, EE.UU.: Boessenecker and Smith; 2011)
Algunas otras aves del Paleógeno (y en un caso posiblemente del Cretácico Superior), típicamente taxones conocidos solo a partir de restos fragmentarios, pueden ser también pelagornítidos. Estos no son usualmente situados aquí, pero el gran tamaño de los fósiles y las similitudes conocidas con ciertos huesos de aves pseudodentadas a los de otros linajes justifican estudios adicionales. Los géneros en cuestión son Laornis, Proceriavis, Manu y Protopelicanus.[58]
Referencias
- Brodkorb (1963: p.262), Olson (1985: p.195), Mlíkovský (2002: p.81)
- Bourdon (2005), Mayr (2008), Boessenecker and Smith (2011)
- Hopson (1964), Olson (1985: pp.199-201), Bourdon (2005), Geraads (2006), Mayr (2009: pp.55,59), Mlíkovský (2009)
- "Pseudodontornis" stirtoni es la única notable excepción: Scarlett (1972) contra Mayr (2009: p.59)
- Publicada en una tesis y por lo tanto un nomen nudum. Asimismo, Odontoptila ya ha sido usado para un género de polilla: ICZN (1999), uBio (2005)
- Scarlett (1972), Olson (1985: pp.199-200), Bourdon (2005, 2006), Mayr (2008, 2009: pp.57,59), Mayr et al. (2008)
- Meunier (1951), Hopson (1964), Olson (1985: p.200) Mayr (2008, 2009: p.58)
- Woodward (1909): pp.86-87, Hopson (1964), Olson (1985: p.142), Bourdon (2005), Mayr (2009: p.58), Mayr et al. (2008)
- Olson (1985: pp.194-195), Mayr (2008), GG [2009]
- Las sepias (Sepiida) y calamares (Teuthida) se diversificaron a través del Paleógeno, y los Argonautoida (los actuales pulpos pelágicos) se originaron en esta época o algo antes. Los pulpos más basales como Keuppia, Palaeoctopus y Styletoctopus del Cretácico Superior eran también al menos parcialmente pelágicos. Como los primeros pelagornítidos tal vez aparecieron a finales del Cretácico, los por entonces diversos Vampyromorphida (de los cuales solo subsiste hoy el calamar vampiro Vampyroteuthis infernalis) eran presas potenciales de las primeras aves pseudodentadas también; aunque los mayores pelagornítidos solo se hallan en el Neógeno, quizás los juveniles del calamar vampiro gigante Tusoteuthis longa fueron devorados por las primeras de estas aves: PD [2009]
- Las anguilas verdaderas (Anguillidae) y los congrios (Congridae) son conocidos desde el Eoceno y pueden haberse originado a principios del Paleógeno o un poco antes. Los peces gato (Plotosidae), Ophidiiformes (brótulas y parientes), ciertamente los Blennioidei (blénidos) y quizás los zoárcidos (Zoarcidae) se conjetura que son de edad similar dado su origen a finales del Cretácico de estos o de los linajes relacionados. Si las aves pseudodentadas tienen un origen cretácico, como es probable, sus presas iniciales pueden haber incluido Enchodontoidei los cuales se extinguieron en el Extinción masiva del Cretácico-Terciario: PD [2009]
- Hopson (1964), Olson (1985: pp.200-201), Ono (1989), del Hoyo et al. (1992: pp.198,204), Warheit (1992, 2001), Rincón R. & Stucchi (2003), Sluijs et al. (2006), Chávez et al. (2007), Mayr (2008, 2009: pp.56-58,217-218)
- Como Desmatophoca, Gomphotaria, Imagotaria, Pelagiarctos y Pliopedia. Algunas de estas tenían cuatro colmillos, y algunas no tenían grandes colmillos en absoluto: PD [2009]
- Begon et al. (2005): pp.123-124, Mayr (2009: pp.217-218), PD [2009]
- Olson (1985: pp.195-199), del Hoyo et al. (1992), Olson & Rasmussen (2001), Price & Clague (2002), Mlíkovský (2002: pp.81-83, 2003), Rincón R. & Stucchi (2003), Sluijs et al. (2006), Mayr (2009: pp.6,56)
- Warheit (1992, 2001), Olson & Rasmussen (2001), Geraads (2006), Chávez et al. (2007), Mayr (2009: pp.217-218), GG [2009], Mlíkovský (2009)
- El enorme megalodonte (Carcharocles megalodon) probablemente podría haber encontrado que incluso las mayores aves pseudodentadas podían ser cazadas sin esfuerzo.
- Olson & Rasmussen (2001), Purdy et al. (2001)
- del Hoyo et al. (1992)
- En las aves acuáticas de cola corta y en las "aves acuáticas superiores" se encuentran frecuentemente colas blancas o claras: del Hoyo et al. (1992)
- Olson (1985: p.198), Mlíkovský (2002: p.81), Mayr (2009: pp.55-59)
- Lanham (1947), Wetmore (1956: pp.12-14), Brodkorb (1963: pp.241,262-264), Hopson (1964), Olson (1985: pp.195-199), Mlíkovský (2002: p.81), Mayr (2009: p.59)
- Lanham (1947), Brodkorb (1963: pp.248-249, 1967: p.141-143), Olson (1985: p.195), Mlíkovský (2002: pp.78,81-83), Mayr (2008, 2009: p.59)
- Woodward (1909): p.87), Brodkorb (1967: p.142), Olson (1985: pp.195,199), Bourdon (2005), Christidis & Boles (2008: p.100), Mayr (2009: p.59)
- Bourdon (2005), Mayr (2008), Mayr (2009: p.59)
- Bourdon (2005)
- Wetmore (1917), Hopson (1964), Olson (1985: pp.199-200), González-Barba et al. (2002), Bourdon (2005), Christidis & Boles (2008: pp.100,105), Mayr (2008, 2009: p.59), Mayr et al. (2008), TZ [2009]
- En particular el enigmático Laornis edvardsianus: Mayr (2009: p.21)
- Bourdon (2005), Livezey & Zusi (2007), Mayr (2009: p.59)
- Stone (1918)
- Howard (1957), Hopson (1964), Brodkorb (1967: p.142), Wetmore (1917), Bourdon (2005), Mayr (2008, 2009), Mayr et al. (2008), TZ [2009]
- Olson (1985: pp.194-195), Mayr (2008), Mayr (2009: pp.55-59)
- Olson (1985: pp.195-199), Mlíkovský (2002: pp.81-84), Mayr (2009: pp.55-59)
- "Odontopteryx n. sp. 1" of Bourdon (2005). Assorted skull and limb bones in the OCP and Rhinopolis Association collections. The smallest pseudotooth bird known as of mid-2009: Bourdon (2005, 2006), Mayr (2009: p.56)
- González-Barba et al. (2002), Mlíkovský (2002: p.81), Mayr (2009: p.57)
- Espécimen MHN-UABCS Te5/6–517. Extremo distal del húmero de una pequeña ave pseudodentada – con un tamaño similar al de un pelícano marrón (Pelecanus occidentalis) – hallado en la Formación Tepetate cerca de El Cien (Baja California Sur, México): González-Barba et al. (2002)
- Una pieza del eje de un húmero de una especie grande: Stilwell et al. (1998)
- Una especie de tamaño medio, al menos en parte anteriormente incluida en Argillornis: Brodkorb (1963: pp.248-249), Mayr (2009: p.56), Mlíkovský (2002: p.83, 2009)
- Una especie grande: Bourdon (2006), Mayr (2009: p.56)
- Una pieza grande de la mandíbula superior y una pieza pequeña de la inferior: Olson (1985: pp.196,199), Tonni (1980), Tonni & Tambussi (1985), Stilwell et al. (1998), Mayr (2009: p.58)
- MP19 (Priaboniense). Sin detalles adicionales: Mlíkovský (2002: p.81)
- Piezas del pico: Mayr (2009: p.56)
- Mlíkovský (2002: p.81)
- Como Argillornis: Goedert (1989)
- Espécimen LACM 128462, un extremo proximal casi completo de una ulna izquierda de la Formación Keasey de Washington County, Oregon; presumiblemente también LACM 127875, fragmentos de los extremos proximales del húmero, la parte proximal de la ulna derecha y el radio de un único individuo de la Formación Pittsburg Bluff cerca de Mist. Una especie grande, quizás la mayor ave pseudodentada conocida. Warheit (2001) lista 2 especies y da "Eoceno Medio" como edad, pero esto es equivocado: Goedert (1989), González-Barba et al. (2002), Mayr (2009: p.57)
- Extremo distal del radio de una especie grande. El material de Proceriavis es una pieza de vértebra cervical (espécimen BMNH A-4413) y quizás una falange del dedo del pie: Mlíkovský (1996, 2002: p.269), Mayr (2009: p.31). "E. helveticus" es un lapsus.
- Una punta del pico de la Formación Iwaki de Ogawa en Iwaki, Fukushima, y material adicional del Grupo Kishima de Kyushu: Ono (1989), Matsuoka et al. (1998), Mayr (2009: p.58)
- Una especie grande, comparable a Osteodontornis y Pelagornis en tamaño: Warheit (2001)
- Extremo distal del húmero izquierdo y algunos fragmentos óseos del ala. La edad del "Mioceno Inferior" en Warheit (2001) es probablemente un error: Matsuoka et al. (1998), González-Barba et al. (2002), Mayr (2009: p.58)
- "Pelagornithidae sp. A" y "Pelagornithidae sp. B" en Warheit (2001); Pelagornis sp. 1 y Pelagornis sp. 2 en Olson & Rasmussen (2001). Mayormente piezas del fémur y el húmero de aves algo menores que Osteodontornis y otras algo mayores que Tympanonesiotes; también se conocen otros huesos. Una tróclea izquierda tarsometatarsal del Mioceno Inferior USNM 476044 es menor y puede ser una tercera especie: Olson (1985: p.198), Rasmussen (1998), Olson & Rasmussen (2001)
- Espécimen MBLUZ-P-5093, una pieza muy grande del premaxilar: Rincón R. & Stucchi (2003)
- Espécimen CMNZ AV 24,960, una pieza proximal del húmero (inicialmente identificada como la pieza distal) del húmero de una especie grande: Scarlett (1972), Olson (1985: p.199), Mlíkovský (2002: p.84)
- MPC 1001 a 1006 (varias piezas del pico y el cráneo, una ulna proximal izquierda y dos vértebras cervicales) del Mioceno Medio de la Formación Bahía Inglesa; antes asignados a Pseudodontornis longirostris por error. UOP/01/81 (la primera falange del segundo dedo izquierdo), UOP/01/79 y UOP/01/80 (un tarsometatarso) derecho dañado, y un [coracoides]] derecho distal del límite Mioceno-Plioceno de la Formación Bahía Inglesa Formation. MNHN tiene los extremos proximal del carpometacarpo y del húmero derecho de la Formación Pisco: Walsh (2000), Walsh & Hume (2001), Chávez & Stucchi (2002), Rincón R. & Stucchi (2003), Chávez et al. (2007)
- MUSM 210 (fragmentos del pico y una vértebra atlas), MUSM 666 (cabeza proximal del húmero derecho), MUSM 667 (ulna proximal) de un ave algo menor que Pelagornis miocaenus; antes asignado a Pseudodontornis por error. El bien preservado cráneo develado en 2009 también muestra "dientes" como los de Osteodontornis pero aparentemente de un ave mayor: Palmer (1999: p.180), Chávez & Stucchi (2002), Chávez et al. (2007), GG [2009]
- Un húmero derecho fragmentario: Ono (1989), Matsuoka et al. (1998), González-Barba et al. (2002)
- radio derecho proximal (colección McKee A080 183) y húmero derecho proximal (colección McKee A111 182) de una especie grande: McKee (1985), Goedert (1989)
- Espécimen MFM 1801, un fémur distal derecho de una especie grande. Inicialmente confundido con un albatros: Ono (1980, 1989), Matsuoka et al. (1998)
- Olson (1985: pp.173,202,208), Mlíkovský (2002: pp.269-270), Mayr (2009: p.21,31,77,80)
Bibliografía
 Wikimedia Commons alberga una categoría multimedia sobre Pelagornithidae.
Wikimedia Commons alberga una categoría multimedia sobre Pelagornithidae.- Begon, Michael; Townsend, Colin A. & Harper, John Lander (2005): Ecology: From Individuals to Ecosystems (4th ed.). Wiley-Blackwell. ISBN 978-1-4051-1117-1 Excerpt at Google Books
- Boessenecker, Robert W. and N. Adam Smith (2011): Latest Pacific basin record of a bony-toothed bird (Aves, Pelagornithidae) from the Pliocene Purísima Formation of California, U.S.A. Journal of Vertebrate Paleontology 31(3): 652–657.
- Bourdon, Estelle (2005): Osteological evidence for sister group relationship between pseudo-toothed birds (Aves: Odontopterygiformes) and waterfowls (Anseriformes). Naturwissenschaften 92(12): 586–591. doi 10.1007/s00114-005-0047-0 (HTML abstract) Electronic supplement (requires subscription)
- Bourdon, Estelle (2006): L'avifaune du Paléogène des phosphates du Maroc et du Togo: diversité, systématique et apports à la connaissance de la diversification des oiseaux modernes (Neornithes) ["Paleogene avifauna of phosphates of Morocco and Togo: diversity, systematics and contributions to the knowledge of the diversification of the Neornithes"]. Doctoral thesis, Muséum national d'histoire naturelle [in French]. HTML abstract
- Brodkorb, Pierce (1963): Catalogue of fossil birds. Part 1 (Archaeopterygiformes through Ardeiformes). Bulletin of the Florida State Museum, Biological Sciences 7(4): 179-293. PDF or JPEG fulltext Archivado el 11 de marzo de 2007 en Wayback Machine.
- Brodkorb, Pierce (1967): Catalogue of Fossil Birds: Part 3 (Ralliformes, Ichthyornithiformes, Charadriiformes). Bulletin of the Florida State Museum 11(3): 99-220. PDF or JPEG fulltext Archivado el 23 de febrero de 2008 en Wayback Machine.
- Chávez, Martín & Stucchi, Marcelo (2002): El registro de Pelagornithidae del Pacífico sudeste [The record of Pelagornithidae in the southeast Pacific]. Actas del Congreso Latinoamericano de Paleontología de Vertebrados 1: 26. [in Spanish]. PDF fulltext
- Chávez, Martín; Stucchi, Marcelo & Urbina, Mario (2007): El registro de Pelagornithidae (Aves: Pelecaniformes) y la Avifauna Neógena del Pacífico Sudeste [The record of Pelagornithidae (Aves: Pelecaniformes) and the Neogene avifauna of the southeast Pacific]. Bulletin de l'Institut Français d’Études Andines 36(2): 175-197 [Spanish with French and English abstracts]. PDF fulltext
- Christidis, Les & Boles, Walter E. (2008): Systematics and Taxonomy of Australian Birds. CSIRO Publishing, CollingwoodVictoria, Australia. ISBN 978-0-643-06511-6
- del Hoyo, Josep; Elliott, Andrew & Sargatal, Jordi (eds.) (1992): Handbook of Birds of the World (Volume 1: Ostrich to Ducks). Lynx Edicions, Barcelona. ISBN 84-87334-10-5
- Geraads, Denis (2006): The late Pliocene locality of Ahl al Oughlam, Morocco: vertebrate fauna and interpretation. Transactions of the Royal Society of South Africa 61(2): 97-101. PDF fulltext doi 10.1080/00359190609519958
- Goedert, James L. (1989): Giant Late Eocene Marine Birds (Pelecaniformes: Pelagornithidae) from Northwestern Oregon. J. Paleontol. 63(6): 939-944. Abstract and first page text
- González-Barba, Gerardo; Schwennicke, Tobias; Goedert, James L. & Barnes, Lawrence G. (2002): Earliest Pacific Basin record of the Pelagornithidae (Aves, Pelecaniformes). J. Vertebr. Paleontol. 22(2): 722-725. doi [0722:EPBROT2.0.CO;2 10.1671/0272-4634(2002)022[0722:EPBROT]2.0.CO;2] HTML abstract
- Hopson, James A. (1964): Pseudodontornis and other large marine birds from the Miocene of South Carolina. Postilla 83: 1–19. Fulltext at the Internet Archive
- Howard, Hildegarde (1957): A gigantic "toothed" marine bird from the Miocene of California. Santa Barbara Museum of Natural History Bulleting (Geology Department) 1: 1-23.
- International Commission on Zoological Nomenclature (ICZN) (1999): International Code of Zoological Nomenclature (4th ed.). International Trust for Zoological Nomenclature, London. ISBN 0-85301-006-4 HTML fulltext
- Lanham, Urless N. (1947): Notes on the phylogeny of the Pelecaniformes. Auk 64(1): 65–70. DjVu fulltext PDF fulltext
- Livezey, Bradley C. & Zusi, Richard L. (2007): Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. II. Analysis and discussion. Zool. J. Linn. Soc. 149(1): 1–95. doi 10.1111/j.1096-3642.2006.00293.x PDF fulltext
- Matsuoka, Hirosige; Sakakura, Fujio & Ohe, Fumio (1998): A Miocene pseudodontorn (Pelecaniformes: Pelagornithidae) from the Ichishi Group of Misato, Mie Prefecture, Central Japan. Paleontological Research 2(4): 246-252. [http://ci.nii.ac.jp/lognavi?name=nels&lang=en&type=pdf&id=ART0002971529 (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última). PDF fulltext]
- Mayr, Gerald (2008): A skull of the giant bony-toothed bird Dasornis (Aves: Pelagornithidae) from the Lower Eocene of the Isle of Sheppey. Palaeontology 51(5): 1107-1116. doi 10.1111/j.1475-4983.2008.00798.x (HTML abstract)
- Mayr, Gerald (2009): Paleogene Fossil Birds. Springer-Verlag, Heidelberg & New York. ISBN 3-540-89627-9 Preview at Google Books
- Mayr, Gerald; Hazevoet, Cornelis J.; Dantas, Pedro; & Cachāo, Mário (2008): A sternum of a very large bony-toothed bird (Pelagornithidae) from the Miocene of Portugal. J. Vertebr. Paleontol. 28(3): 762-769. DOI:10.1671/0272-4634(2008)28[762:ASOAVL]2.0.CO;2 HTML abstract
- McKee, Joseph W.A. (1985): A pseudodontorn (Pelecaniformes: Pelagornithidae) from the middle Pliocene of Hawera, Taranaki, New Zealand. New Zealand Journal of Zoology 12(2): 181-184. Fulltext (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última). at Google Books
- Meunier, K. (1951): Korrelation und Umkonstruktionen in den Größenbeziehungen zwischen Vogelflügel und Vogelkörper ["Correlation and restructuring in the size relationship between avian wing and avian body"]. Biologia Generalis 19: 403-443 [Article in German].
- Mlíkovský, Jirí (1996): Tertiary avian localities of the United Kingdom. Acta Universitatis Carolinae: Geologica 39: 759–771. PDF fulltext
- Mlíkovský, Jirí (2002): Cenozoic Birds of the World, Part 1: Europe. Ninox Press, Prague. ISBN 80-901105-3-8 PDF fulltext
- Mlíkovský, Jirí (2003): Early Miocene birds of Djebel Zelten, Libya. Časopis Národního muzea, Řada přírodovědná (J. Nat. Mus., Nat. Hist. Ser.) 172: 114–120. PDF fulltext
- Mlíkovský, Jirí (2009): Evolution of the Cenozoic marine avifaunas of Europe. Annalen des Naturhistorischen Museums Wien A 111: 357–374 PDF fulltext
- Olson, Storrs L. (1985): The Fossil Record of Birds. In: Farner, D.S.; King, J.R. & Parkes, Kenneth C. (eds.): Avian Biology 8: 79-252. PDF fulltext Archivado el 18 de julio de 2011 en Wayback Machine.
- Olson, Storrs L. & Rasmussen, Pamela C. (2001): Miocene and Pliocene Birds from the Lee Creek Mine, North Carolina. In: Ray, Clayton E. & Bohaska, David J. (eds.): Geology and Paleontology of the Lee Creek Mine, North Carolina, III. Smithsonian Contributions to Paleobiology 90: 233-307. PDF fulltext
- Ono, Keiichi (1980): Pliocene Tubinare Bird from Kakegawa, Shizuoka Prefecture, Japan. Memoirs of the National Science Museum 13(1): 29-34, plate 2 [Japanese with English abstract]. [http://ci.nii.ac.jp/lognavi?name=nels&lang=en&type=pdf&id=ART0006481420 (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última). PDF fulltext]
- Ono, Keiichi (1989): A Bony-Toothed Bird from the Middle Miocene, Chichibu Basin, Japan. Bulletin of the National Science Museum Series C: Geology & Paleontology 15(1): 33-38. [http://ci.nii.ac.jp/lognavi?name=nels&lang=en&type=pdf&id=ART0006480358 (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última). PDF fulltext]
- Paleobiology Database (PD) [2009]: Taxonomic name search form. Retrieved on 2009-AUG-11.
- Palmer, D. (1999): The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals. Marshall Editions, London. ISBN 1-84028-152-9
- Price, Jordan P. & Clague, David A. (2002): How old is the Hawaiian biota? Geology and phylogeny suggest recent divergence. Proc. R. Soc. B 269(1508): 2429-2435. doi 10.1098/rspb.2002.2175 PDF fulltext
- Purdy, Robert W.; Schneider, Vincent P.; Applegate, Shelton P.; McLellan, Jack H.; Meyer, Robert L. & Slaughter, Bob H. (2001): The Neogene Sharks, Rays, and Bony Fishes from Lee Creek Mine, Aurora, North Carolina. In: Ray, Clayton E. & Bohaska, David J. (eds.): Geology and Paleontology of the Lee Creek Mine, North Carolina, III. Smithsonian Contributions to Paleobiology 90: 71-202. PDF fulltext
- Rasmussen, Pamela C. (1998): Early Miocene Avifauna from the Pollack Farm Site, Delaware. Delaware Geological Survey Special Publication 21: 149-151. PDF fulltext (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Rincón R., Ascanio D. & Stucchi, Marcelo (2003): Primer registro de la familia Pelagornithidae (Aves: Pelecaniformes) para Venezuela [First record of Pelagornithidae family from Venezuela]. Boletín de la Sociedad Venezolana de Espeleología 37: 27-30 [Spanish with English abstract]. PDF fulltext
- Scarlett, R.J. (1972): Bone of a presumed odontopterygian bird from the Miocene of New Zealand. New Zealand Journal of Geology and Geophysics 15(2): 269-274. Fulltext at Google Books
- Sluijs, Appy; Schouten, Stefan; Pagani, Mark; Woltering, Martijn; Brinkhuis, Henk; Damsté, Jaap S. Sinninghe; Dickens, Gerald R.; Huber, Matthew; Reichart, Gert-Jan; Stein, Rüdiger; Matthiessen, Jens; Lourens, Lucas J.; Pedentchouk, Nikolai; Backman, Jan; Moran, Kathryn & the Expedition 302 Scientists (2006): Subtropical Arctic Ocean temperatures during the Palaeocene/Eocene thermal maximum. Nature 441(7093): 610–613. doi 10.1038/nature04668 PDF fulltext Archivado el 10 de abril de 2013 en Wayback Machine. Supplementary Notes
- Stilwell, Jeffrey D.; Jones, Craig M.; Levy, Richard H. & Harwood, David M. (1998): First fossil bird from East Antarctica. Antarctic Journal of the United States 33(1): 12-16. PDF fulltext
- Stone, Witmer (1918): Recent Literature – Wetmore on Palæochenoides mioceanus Shufeldt. Auk 35(2): 251. DjVu fulltext PDF fulltext
- Tetrapod Zoology (TZ) [2009]: 2007: a good year for terror birds and mega-ducks. Version of March 14-MAR-2009. Retrieved 2009-AUG-11.
- Tonni, Eduardo Pedro (1980) Un pseudodontornitido [Pelecaniformes, Odontopterygia] de gran tamaño, del Terciario temprano de Antártida. ["A large-sized pseudotooth bird from the Early Tertiary of Antarctica"]. Ameghiniana 17(3): 273-276 [Spanish with English abstract]. HTML abstract
- Tonni, Eduardo Pedro & Tambussi, Claudia Patricia (1985): Nuevos restos de Odontopterygia (Aves: Pelecaniformes) del Terciario temprano de Antártida ["New pseudotooth bird remains from the Early Tertiary of Antarctica"]. Ameghiniana 21(2-4): 121-124 [Spanish with English abstract]. HTML abstract
- uBio (2005): Digital Nomenclator Zoologicus, version 0.86 3: 387. PDF fulltext
- Walsh, Stig A. (2000): Big-chested birds – exciting new avian material from the Neogene of Chile. Talk held at the 48th Annual Symposium of Vertebrate Palaeontology and Comparative Anatomy, 1 September 2000, Portsmouth, UK. HTML abstract
- Walsh, Stig A. & Hume, Julian P. (2001): A new Neogene marine avian assemblage from north-central Chile. J. Vertebr. Paleontol. 21(3): 484-491. DOI:10.1671/0272-4634(2001)021[0484:ANNMAA]2.0.CO;2 PDF fulltext (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Warheit, Kenneth I. (1992): A Review of the Fossil Seabirds from the Tertiary of the North Pacific: Plate Tectonics, Paleoceanography, and Faunal Change. Paleobiology 18(4): 401-424. Abstract and first page text
- Warheit, Kenneth I. (2001): The seabird fossil record and the role of paleontology in understanding seabird community structure. In: Schreiber, E.A. & Burger, J. (eds.): Biology of marine birds: 17-55. CRC Press, Boca Raton, Florida, USA. ISBN 0-8493-9882-7 Excerpt at Google Books
- Wetmore, Alexander (1918): The Relationships of the Fossil Bird Palaeochenoides mioceanus. J. Geol. 25(6): 555-557. First page image
- Wetmore, Alexander (1956): A check-list of the fossil and prehistoric birds of North America and the West Indies. Smithsonian Miscellaneous Collections 131(5): 1-105. Fulltext at the Internet Archive
- Woodward, Arthur Smith (ed.) (1909): A Guide to the Fossil Mammals and Birds in the Department of Geology and Palaeontology of the British Museum (Natural History) (9th ed.). William Clowes and Sons Ltd., London. Fulltext at the Internet Archive
| Control de autoridades |
|
|---|
 Datos: Q143473
Datos: Q143473- Multimedia: Pelagornithidae / Q143473
 Especies: Pelagornithidae
Especies: Pelagornithidae