Membrana plasmática eucariota
La membrana plasmática eucariota, también llamada membrana celular, membrana citoplasmática o plasmalema de las células eucariotas, es una bicapa lipídica que delimita toda la célula.[1] Es una estructura formada por dos láminas de fosfolípidos, glucolípidos y proteínas[2] que rodean, limitan la forma y contribuyen a mantener el equilibrio entre el interior (medio intracelular) y el exterior (medio extracelular) de las células. Regula la entrada y salida de muchas sustancias entre el citoplasma y el medio extracelular.[3] Es similar a las membranas que delimitan a las células procariotas, en especial al de las bacterias, y es similar también a las membranas de los orgánulos del sistema endomembranoso.
| Membrana plasmática eucariota | ||
|---|---|---|
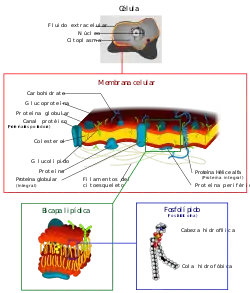 Ilustración de la membrana celular de una célula eucariota. | ||
| Nombre y clasificación | ||
| Sinónimos |
Plasmalema | |
| Latín | Plasmalemma; Membrana cellularis | |
| TH | H1.00.01.0.00011 H1.00.01.1.00001 | |
|
| ||
Está compuesta por dos láminas que sirven de "contenedor" para el citosol y los distintos compartimentos internos de la célula, así como también otorga protección mecánica. Está formada principalmente por fosfolípidos (fosfatidiletanolamina y fosfatidilcolina), colesterol, glúcidos y proteínas (integrales y periféricas).
La principal característica de esta barrera es su permeabilidad selectiva, lo que le permite seleccionar las moléculas que deben entrar y salir de la célula. De esta forma se mantiene estable el medio intracelular, regulando el paso de agua, iones y metabolitos, a la vez que mantiene el potencial electroquímico (haciendo que el medio interno esté cargado negativamente). La membrana plasmática es capaz de recibir señales que permiten el ingreso de partículas a su interior.
Cuando una molécula de gran tamaño atraviesa o es expulsada de la célula y se invagina parte de la membrana plasmática para recubrirlas cuando están en el interior ocurren respectivamente los procesos de endocitosis y exocitosis.
Tiene un grosor aproximado de 7,4 nm[4] (74 Å) y no es visible al microscopio óptico pero sí al microscopio electrónico, donde se pueden observar dos capas oscuras bilaterales y una central más clara. En las células eucariotas osmótrofas como plantas y hongos, se sitúa bajo otra capa exterior, denominada pared celular.
En la actualidad se ha descubierto que es posible que estas estructuras se formen sin la presencia de agua, a partir de metano líquido, lo que abre la posibilidad a encontrar vida fuera de la Tierra.[5]
Composición química
La membrana citoplasmática o plasmática es una estructura laminar que engloba a las células, define sus límites y contribuye a mantener el equilibrio entre el interior y el exterior. Es semejante además, a las membranas que delimitan los orgánulos de células eucariotas. Es una bicapa lipídica que sirve de "contenedor" para los contenidos de la célula, así como protección mecánica. Está formada principalmente por lípidos y proteínas. Esta barrera presenta una permeabilidad selectiva, lo cual le permite "seleccionar" las moléculas que entran y salen de la célula. Tiene un grosor aproximado de 75 Å. No es visible a microscopio óptico pero si a microscopio electrónico. Con un microloscopio todo maqueado se pueden observar tres láminas: dos láminas son osmiófilas (tiene más apetencia por el osmio y por ello se ven más oscuras) y una central osmiófoba (más clara). La lámina central tiene un grosor de entre 25 y 30 Å y las laterales de entre 20 y 25 Å. Las láminas osmiófilas se corresponden a las cabezas polares de los fosfolípidos y la lámina osmiófoba se corresponde a las colas de ácidos grasos. Presenta pliegues llamados interdigitaciones que amplían la zona de contacto entre células haciendo que se unan más fuerte entre ellas.
Precedentes históricos
- En 1895, Overton asegura que la membrana tiene una estructura lipídica
- En 1932, Cole observa proteínas acompañando a los lípidos
- En 1935, Danielli y Davson descubren que la membrana plasmática presenta poros.
- En 1957, Robertson observó que la membrana plasmática estaba compuesta por las tres láminas.
- En 1972, Singer y Nicholson, proponen el modelo de mosaico fluido. Esto fue posible gracias a los avances en microscopía electrónica, el estudio de interacciones hidrófilas, al estudio de enlaces no covalentes como puentes de hidrógeno y el desarrollo de técnicas como criofractura y contraste negativo[6]
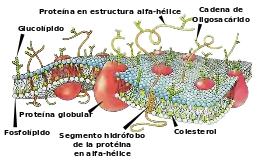
Antiguamente se creía que la membrana plasmática era un conjunto estático formado por la sucesión de capas proteínas-lípidos-lípidos-proteínas. Hoy en día se concibe como una estructura dinámica cuyo modelo se conoce como "mosaico fluido", término acuñado por S. J. Singer y G. L. Nicolson en 1972. Es una estructura líquida bidimensional de dos moléculas de grosor, en la que una porción de la misma puede fluir literalmente desde un punto de la membrana hasta otro. Esta estructura general —modelo unitario— se presenta también en todo el sistema de endomembranas (membranas de los diversos orgánulos del interior de la célula), como retículo endoplasmático, aparato de Golgi y envoltura nuclear, y los de otros orgánulos, como las mitocondrias y los plastos, que proceden de endosimbiosis.
La composición química de la membrana plasmática varía entre células dependiendo de la función o del tejido en la que se encuentren, pero se puede estudiar de forma general. La membrana plasmática está compuesta por una doble capa de fosfolípidos, por proteínas unidas no covalentemente a esa bicapa, y glúcidos unidos covalentemente a los lípidos o a las proteínas. Las moléculas más numerosas son los lípidos, ya que se calcula que por cada 50 lípidos hay una proteína. Sin embargo, las proteínas, debido a su mayor tamaño, representan aproximadamente el 50 % de la masa de la membrana.
Bicapa lipídica
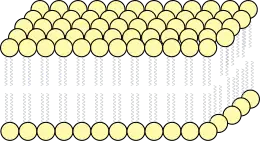
El orden de las llamadas cabezas hidrofílicas y las colas hidrofóbicas de la bicapa lipídica impide que solutos polares, como sales minerales, agua, carbohidratos y proteínas, difundan a través de la membrana, pero generalmente permite la difusión pasiva de las moléculas hidrofóbicas. Esto permite a la célula controlar el movimiento de estas sustancias vía complejos de proteína transmembranal tales como poros y caminos, que permiten el paso de iones específicos como el sodio y el potasio.
Las dos capas de moléculas fosfolípidas forman un "sándwich" con las colas de ácido graso dispuestos hacia el centro de la membrana plasmática y las cabezas de fosfolípidos hacia los medios acuosos que se encuentran dentro y fuera de la célula.
Componentes lipídicos
El 98 % de los lípidos presentes en las membranas celulares son los anfipáticos, es decir que presentan un extremo hidrófilo (que tiene afinidad e interacciona con el agua) y un extremo hidrofóbico (que repele el agua). Los tres principales tipos de lípidos en las membranas eucarióticas son los fosfolípidos, los esfingoglucolípidos y esteroides (sobre todo colesterol); cabe mencionar que los fosfoglicéridos y los esfingolípidos se encuentran en todas las células. Estos últimos, glucolípidos y esteroides, no existen o son escasos en las membranas plasmáticas de las células procariotas, hongos y algunos protistas. Existen también grasas neutras, que son lípidos no anfipáticos, pero solo representan un 2 % del total de lípidos de membrana.
- Fosfoglicéridos. Tienen una molécula de glicerol con la que se esterifica un ácido fosfórico y dos ácidos grasos de cadena larga; los principales fosfoglicéridos de membrana son la fosfatidiletanolamina o cefalina, la fosfatidilcolina o lecitina, el fosfatidilinositol y la fosfatidilserina.Este tipo de glicéridos son diglicéridos, es decir que solo dos grupos hidróxilo del glicerol están esterificados con ácidos grasos y el tercero lo está con un grupo fosfato, hidrófilo, por un enlace fosfoéster.[7]
- Esfingolípidos. Son lípidos de membrana constituidos por ceramida (esfingosina + ácido graso); solo la familia de la esfingomielina posee fósforo; el resto poseen glúcidos y se denominan por ello glucoesfingolípidos o, simplemente glucolípidos. Los cerebrósidos poseen principalmente glucosa, galactosa y sus derivados (como N-acetilglucosamina y N-acetilgalactosamina). Los gangliósidos contienen una o más unidades de ácido N-acetilneuramínico (ácido siálico).
- Colesterol. El colesterol representa un 23 % de los lípidos de membrana. Sus moléculas son pequeñas y más anfipáticas en comparación con otros lípidos. Se dispone con el grupo hidroxilo hacia el exterior de la célula (ya que ese hidroxilo interactúa con el agua). El colesterol es un factor importante en la fluidez y permeabilidad de la membrana ya que se hace hueco, a modo de cuña, entre las otras moléculas. A mayor cantidad de colesterol, menos permeable y más dura es la membrana. Se ha postulado que los lípidos de membrana se podrían encontrar en dos formas: como un líquido bidimensional, y de una forma más estructurada, en particular cuando están unidos a algunas proteínas formando las llamadas balsas lipídicas. Se cree que el colesterol podría tener un papel importante en la organización de estas últimas. Su función en la membrana plasmática es evitar que se adhieran las colas de ácido graso de la bicapa, mejorando la fluidez de la membrana. En las membranas de las células vegetales son más abundantes los fitoesteroles.
Componentes proteicos
El porcentaje de proteínas oscila entre un 20 % en la mielina de las neuronas y un 70 % en la membrana interna mitocondrial;[8] el 80 % son intrínsecas, mientras que el 20 % restantes son extrínsecas. Las proteínas son responsables de las funciones dinámicas de la membrana, por lo que cada membrana tienen una dotación muy específica de proteínas; las membranas intracelulares tienen una elevada proporción de proteínas debido al elevado número de actividades enzimáticas que albergan. En la membrana las proteínas desempeñan diversas funciones: transportadoras, conectoras (conectan la membrana con la matriz extracelular o con el interior), receptoras (encargadas del reconocimiento celular, adhesión) y enzimas.
Las proteínas de la membrana plasmática se pueden clasificar según cómo se dispongan en la bicapa lipídica:[9][10][11]
- Proteínas integrales. Embebidas en la bicapa lipídica, atraviesan la membrana una o varias veces, asomando por una o las dos caras (proteínas transmembrana); o bien mediante enlaces covalentes con un lípido o un glúcido de la membrana. Su aislamiento requiere la ruptura de la bicapa. Muchas de estas proteínas forman canales iónicos estructurales a través de los cuales difunden de forma selectiva moléculas de agua y sustancias hidrosolubles, en especial los iones, entre los líquidos intracelular y extracelular. Otras actúan como proteínas transportadoras, y otras como enzimas.[12]
- Proteínas periféricas. Se encuentran a un lado u otro de la bicapa lipídica aunque principalmente en el interior, y pueden estar unidas débilmente por enlaces no covalentes, a menudo a las proteínas integrales. Fácilmente separables de la bicapa sin provocar su ruptura, ya que no la penetran. Funcionan casi exclusivamente como enzimas o como reguladores de la función intracelular.[12]
- Proteína de membrana fijada a lípidos. Se localiza fuera de la bicapa lipídica, ya sea en la superficie extracelular o intracelular, conectada a los lípidos mediante enlaces covalentes.
En el componente proteico reside la mayor parte de la funcionalidad de la membrana; las diferentes proteínas realizan funciones específicas:
- Proteínas estructurales o de anclaje: estas proteínas hacen de "eslabón clave" uniéndose al citoesqueleto y la matriz extracelular.
- Proteínas receptoras: que se encargan de la recepción y transducción de señales químicas.
- Proteínas de transporte: mantienen un gradiente electroquímico mediante el transporte de membrana de diversos iones.
- Estas a su vez pueden ser:
- Proteínas transportadoras: Son enzimas con centros de reacción que sufren cambios conformacionales.
- Proteínas de canal: Dejan un canal hidrofóbico por donde pasan los iones hidrofílicos.
Componentes glucídicos
Se encuentran dirigidos hacia el exterior de la membrana unidos covalentemente a las proteínas o a los lípidos, formando glucoproteínas y glucolípidos, donde las porciones glucídicas quedan suspendidas fuera de la pared celular. De hecho la mayoría de las proteínas integrales son glupoproteínas, y alrededor de un 10% de los lípidos son glucolípidos. Pueden ser polisacáridos u oligosacáridos. Representan el 8 % del peso seco de la membrana plasmática.
Se encuentran en el exterior de la membrana formando parte del glicocalix, junto con alguna cadena proteica.
Sus principales funciones son dar soporte a la membrana y el reconocimiento celular (colaboran en la identificación de las señales químicas de la célula).
Funciones
La membrana celular cumple varias funciones: [13]
- Delimita y protege las células. La función principal de la membrana plasmática es mantener el medio interno separado de la capa fosfolipídica y a las funciones de transporte que desempeñan las proteínas. La combinación de transporte activo y transporte pasivo hacen de la membrana endoplásmica una barrera selectiva que permite a la célula diferenciarse del medio.
- Es una barrera selectivamente permeable, ya que impide el libre intercambio de materiales de un lado a otro, pero al mismo tiempo proporcionan el medio para comunicar un espacio con otro.
- Permite el paso o transporte de solutos de un lado a otro de la célula, pues regula el intercambio de sustancias entre el interior y el exterior de la célula siguiendo un gradiente de concentración.
- Poseen receptores químicos que se combinan con moléculas específicas que permiten a la membrana recibir señales y responder de manera específica, por ejemplo, inhibiendo o estimulando actividades internas como el inicio de la división celular, la elaboración de más glucógeno, movimiento celular, liberación de calcio de las reservas internas.
- Permite a la célula dividir en secciones los distintos orgánulos y así proteger las reacciones químicas que ocurren en cada uno.
- Transporta sustancias de un lugar de la membrana a otro, por ejemplo, acumulando sustancias en lugares específicos de la célula que le puedan servir para su metabolismo.
- Percibe y reacciona ante estímulos provocados por sustancias externas (ligando).
- Mide las interacciones que ocurren entre células internas y externas.
Diferenciaciones de la membrana
Van dirigidas al desempeño de funciones concretas y consistentes en algún tipo de alteración morfológica del contorno de la célula en cualquiera de sus superficies:
- Superficie apical (que da hacia la luz del conducto): son típicas las microvellosidades de algunas células epiteliales. Se tratan de evaginaciones con forma de dedo de guante que aumentan la superficie de absorción intestinal.[14]
- Superficie basal (lado opuesto a la luz del conducto): también destacan las células epiteliales, concretamente las que en el riñón presentan invaginaciones, que aumentan la superficie de reabsorción de agua en el tubo contorneado proximal de las nefronas.
- Superficie lateral: son las denominadas uniones intercelulares que posibilitan las interacciones entre células vecinas. Son de varios tipos: estrechas o impermeables, que no dejan espacio intercelular alguno, comunicantes o en hendidura, que dejan un reducido espacio intercelular, y adherentes o desmosomas, que, aunque con un espacio intercelular mayor,[15] implican una fuerte unión mecánica entre las neuronas.
Permeabilidad
La permeabilidad de las membranas es la facilidad de las moléculas para atravesarla. Esto depende principalmente de la carga eléctrica y, en menor medida, de la masa molar de la molécula. Moléculas pequeñas o con carga eléctrica neutra pasan la membrana más fácilmente que elementos cargados eléctricamente y moléculas grandes. Además, la membrana es selectiva, lo que significa que permite la entrada de unas moléculas y restringe la de otras.
La bicapa lipídica, debido a su interior hidrofóbico, actúa como una barrera altamente impermeable a la mayoría de moléculas polares, impidiendo que la mayor parte del contenido hidrosoluble de la célula salga de ella. Pero por esta misma razón, las células han tenido que desarrollar sistemas especiales para transporte las moléculas polares a través de sus membranas.
Con el tiempo suficiente, esencialmente cualquier molécula difundirá a través de una bicapa lipídica libre de proteínas, a favor de su gradiente de concentración. Sin embargo la velocidad a la que una molécula difunde a través de una bicapa lipídica varía enormemente, dependiendo en gran parte del tamaño de la molécula y de su solubilidad relativa al aceite (es decir, cuanto más hidrofóbica o no polar), tanto más rápidamente difundirá a través de una bicapa.
Las moléculas pequeñas no polares se disuelven fácilmente en las bicapas lipídicas y por lo tanto difunden con rapidez a través de ellas. Las moléculas polares sin carga si su tamaño es suficientemente reducido también difunden rápidamente a través de una bicapa. Ejemplos de estas sustancias no polares son los solventes orgánicos, que presentan una polaridad alta o baja. Por ejemplo: el metanol, la acetona, el etanol, la urea, etc.
La reacción que provocan en la membrana plasmática, dichos solventes, al no ser capaces de atravesar dicha membrana, es de degradación, al ser moléculas muy polares provocan que la bicapa lipídica se degrade, que sufra un desgaste. Hay que tomar en cuenta que la permeabilidad de cada soluto se expresa como su penetración relativa. Los alcoholes, como ejemplo de ellos el metanol, etanol, butanol, octanol, etc., pueden actuar en las membranas biológicas fundamentalmente de 3 formas:
- alterando la fluidez de las membranas, lo que indirectamente afectaría el funcionamiento de las proteínas como enzimas y canales;
- produciendo una deshidratación a nivel de las membranas;
- interactuando directamente con las proteínas de la membrana.
La membrana plasmática puede sufrir un proceso llamado lisis, que hace referencia al rompimiento de la membrana, ya sea mecánicamente, químicamente o por alguna combinación de los dos. Para realizar la lisis química, las células se suspenden en una solución que contiene detergentes y otros reactivos que interfieren con los enlaces químicos que sostienen las proteínas de las membranas juntas. Esto resulta en la rotura de la membrana y la liberación de los componentes intracelulares.
Existen dos tipos de lisis: la lisis tradicional (mecánica) y la lisis por medio de detergentes (química) haciendo referencia al párrafo anterior:
Dentro de la tradicional se encuentran tres ejemplos; homogeneización líquida, donde las células se rompen al ser forzadas a pasar por espacios muy pequeños; sonificación, aplicada a ondas de alta frecuencia rompen las células y congelamiento, lo cual son ciclos de congelación continuos que rompen la célula induciendo la formación de cristales. De igual manera está la lisis por medio de detergentes (química), donde los detergentes rompen la barrera lipídica de una manera suave, solubilizando las proteínas e interrumpiendo la interacción lípido-lípido, lípido-proteína y proteína-proteína. Los detergentes, al igual que los lípidos, se asocian entre ellos y se unen a superficies hidrofóbicas. Se componen de una cabeza polar hidrofílica y una cola no polar hidrofóbica.
La permeabilidad depende de los siguientes factores:
- Solubilidad en los lípidos: Las sustancias que se disuelven en los lípidos (moléculas hidrófobas, no polares) penetran con facilidad en la membrana dado que está compuesta en su mayor parte por fosfolípidos.
- Tamaño: la más grande parte de las moléculas de gran tamaño no pasan a través de la membrana. Solo un pequeño número de moléculas polares de pequeño tamaño pueden atravesar la capa de fosfolípidos.
- Carga: Las moléculas cargadas y los iones no pueden pasar, en condiciones normales, a través de la membrana. Sin embargo, algunas sustancias cargadas pueden pasar por los canales proteicos o con la ayuda de una proteína transportadora.
También depende de las proteínas de membrana de tipo:
- Canales: algunas proteínas forman canales llenos de agua por donde pueden pasar sustancias polares o cargadas eléctricamente que no atraviesan la capa de fosfolípidos.
- Transportadoras: otras proteínas se unen a la sustancia de un lado de la membrana y la llevan al otro lado donde la liberan.
Véase también
Referencias
- Tortora, Gerard J.; Funke, Berdell R.; Case, Christine L. (2007). Introducción a la microbiología. Ed. Médica Panamericana. ISBN 9789500607407. Consultado el 22 de febrero de 2018.
- Pocock, Gillian; Richards, Christopher D. (2005). Pocock, G., Fisiología Humana, 2a ed. ©2005. Elsevier España. ISBN 9788445814796. Consultado el 24 de septiembre de 2019.
- Curtis, Helena; Schnek, Adriana (30 de junio de 2008). Curtis. Biología. Ed. Médica Panamericana. ISBN 9789500603348. Consultado el 22 de febrero de 2018.
- Gartner, Leslie P. (24 de febrero de 2017). Texto de histología + StudentConsult: Atlas a color. Elsevier España. ISBN 9788491131908. Consultado el 22 de febrero de 2018.
- «¿Pueden existir las membranas celulares y, por tanto, la vida, si no hay agua?». Investigación y Ciencia. 2015.
- Datos de la historia de las investigaciones de la membrana plasmática en la UNAM
- Karp, G. 2014. "Biología celular y molecular". 7a edición, Ed. McGrawHill
- Devlin, T. M. 2004. Bioquímica, 4.ª edición. Reverté, Barcelona. ISBN 84-291-7208-4
- Alberts et al, Introducción a la Biología Celular, pág. 375-376, 2ª edición, Ed. Médica Panamericana
- Alberts et al, Biología Molecular de la célula, pág. 595, 4ª edición, Ed. Omega
- Cooper, La célula, pág 470-471, 2ª edición, Ed. Marbán
- Guyton, Arthur C. (2001). Tratado de fisiología médica (10. ed. en español edición). McGraw-Hill Interamericana. p. 14. ISBN 970-10-3599-2. OCLC 49608187. Consultado el 10 de junio de 2021.
- Campos, Patricia (2002). Biologia/ Biology. Editorial Limusa. ISBN 9789681860783. Consultado el 22 de febrero de 2018.
- Brusco, Herminia Alicia; López Costa, Juan José; Loidl, César Fabián (23 de julio de 2014). Histología médico-práctica + StudentConsult en español. Elsevier España. ISBN 9788490227572. Consultado el 22 de febrero de 2018.
- García Poblete, Eduardo; Fernández García, Héctor; Rodríguez, José Ernesto Moro; Ocio, José Antonio Uranga; Bona, María Paz Nieto; Heras, Mª Soledad García Gómez de las; Mora, Nora Sánchez (20 de noviembre de 2006). Histología humana práctica: Enfermería. Universitaria Ramon Areces. ISBN 9788480047906. Consultado el 22 de febrero de 2018.
| Control de autoridades |
|
|---|
 Datos: Q88869758
Datos: Q88869758