Sistema endomembranoso
El sistema endomembranoso es el sistema de membranas internas existente en las células eucariotas, que divide la célula en compartimentos funcionales y estructurales, denominados orgánulos.
Los siguientes orgánulos son ejemplos de partes del sistema endomembranoso de las células eucariotas:
- La #Membrana plasmática o membrana celular funciona como barrera de protección que regula la entrada y salida de sustancias en la células.[1]
- La #Envoltura nuclear está formada por un complejo de dos membranas, que delimita los contenidos del núcleo mediante la membrana nuclear interna (INM) y al mismo tiempo forma una cisterna perinuclear, separada del citoplasma mediante la membrana nuclear externa (ONM).[2]
- El #Retículo endoplasmático es extenso distribuido en toda la célula, construido por sáculos (cisternas) y túbulos, que funciona como orgánulo de síntesis y transporte, como una extensión de la envoltura nuclear.[3]
- El #Aparato de Golgi un sistema multicisternal compacto y localizado, que funciona como el sistema de empaquetado y de entrega de moléculas.[4]
- Los #Lisosomas son las unidades “energéticas” de la célula. Utilizan enzimas que analizan las macromoléculas y también actúan como sistema de recogida de residuos.
- Las #Vacuolas actúan como unidades del almacenaje en algunas células.[5]
- Las #Vesículas son pequeñas unidades de transporte delimitadas por membranas que pueden transferir moléculas entre diversos compartimientos.[6]

Cabe mencionar que las membranas de cloroplastos y mitocondrias no están consideradas como parte del sistema, pero se piensa que evolucionaron a partir de este.
El sistema endomembranoso también proporciona un sistema de transporte para las moléculas móviles a través del interior de la célula, así como superficies interactivas para la síntesis de lípidos y de proteínas.
Los orgánulos del sistema endomembranoso pueden estar intercomunicados de dos maneras: mediante contacto directo o mediante la trasferencia de segmentos de membrana en forma de vesículas.[7] A pesar de esto, las membranas de los diferentes orgánulos no son idénticas; varían sus estructuras y funciones. El grosor, la composición molecular, y el comportamiento metabólico de una membrana pueden ser modificados a lo largo de la vida de la célula. Las membranas que componen el sistema endomembranoso se construyen a partir de una bicapa lipídica, con proteínas unidas a cada lado o atravesándolas y otras moléculas de reconocimiento.[8]
En células procariotas es raro encontrar endomembranas, sin embargo en diversas bacterias fotosintéticas la membrana plasmática se encuentra plegada hacia el interior de la célula de modo que su citoplasma se encuentra rodeado de capas de membrana capaces de captar luz.[9] Estas capas pueden incluso estar encapsuladas en estructuras llamadas clorosomas en bacterias tipo chlorobia.[10]
Desarrollo y formación
Este sistema endomembranoso parece comenzar en el retículo endoplasmático rugoso (RER) que es un conjunto de membranas plegadas que son una prolongación de la membrana nuclear. En él la principal función es la síntesis de proteínas. De ellas, algunas se almacenan para luego llevarlas a otros orgánulos; y otras se quedan adosadas al mismo para hacer crecer sus membranas.
Llegado a un punto variable de su longitud, el RE ya no tiene ribosomas y forma túbulos y cisternas membranosas donde principalmente se forman lípidos es el retículo endoplasmático liso. En él se generan unas estructuras que secretan vesículas de transición, que se van uniendo formando el dictiosoma. El dictiosoma es la unidad de funcionamiento del aparato de Golgi. El conjunto de dictiosomas de una célula forma el aparato de Golgi. Esta estructura finaliza con unas vesículas de síntesis que son secretadas al citoplasma y luego darán lugar a los lisosomas, peroxisomas y vacuolas.
Componentes del Sistema
Envoltura Nuclear

RE
perinuclear>
Membrana
Nuclear
Interna
Membrana
Nuclear
Externa
nuclear
La envoltura nuclear separa los contenidos de núcleo de aquellos del citoplasma. Está formada una membrana nuclear interna (INM en inglés) y una membrana nuclear externa (ONM en inglés).[11]
La membrana nuclear externa (ONM) presenta ribosomas sobre su superficie citoplásmica y se continúa con la membrana del retículo endoplasmático rugoso (RER). La membrana nuclear externa se continúa con la membrana nuclear interna (INM), en los llamados poros nucleares, pequeños orificios que tienen un diámetro de 120 nanómetros (nm).[12]
El espacio entre las dos membranas de la envoltura nuclear, se conoce como el espacio perinuclear (o cisterna perinuclear) y se encuentra unido y se continua con el lumen del RER.
La estructura de la envoltura nuclear está determinada por proteínas fibrosas llamadas filamentos intermedios. Estos forman una malla denominada lámina nuclear, la cual se acopla a la cromatina, proteínas integrales de membrana y otros componentes nucleares que se encuentran alineados a la superficie interna del núcleo. Se cree que la lámina nuclear ayuda a que sustancias dentro del núcleo lleguen a los poros nucleares, así como en la desintegración de la envoltura nuclear durante la mitosis, así como su reconstrucción al finalizarla.
Los poros nucleares tienen una selectividad muy efectiva al momento de regular el paso de materiales a través de la envoltura nuclear, considerando el gran tráfico molecular en el que se encuentran. El RNA y las subunidades ribosomales están en continuo movimiento entre el citoplasma y el núcleo. Otras sustancias como histonas, proteínas reguladoras de genes, DNA y polimerasas de RNA deben ser importadas del citoplasma, por lo que deben atravesar los poros. La envoltura nuclear típica de una célula mamífera contiene entre 3000 y 4000 poros. Cuando la célula se encuentra en proceso de síntesis de DNA cada poro transporta alrededor de 100 moléculas de histona por minuto. En etapas de rápido crecimiento cada poro también transporta subunidades ribosomales del citosol al núcleo, a una tasa de seis por minuto, aproximadamente. Estas son recién armadas y de diferentes tamaños. Estas se utilizan para la síntesis de proteínas[13]
Retículo endoplasmático
El retículo endoplasmático (RE) es un orgánulo membranoso encargado de la síntesis y transporte de algunas biomoléculas. Es una continuación de la envoltura nuclear. En el RE se encuentra más de la mitad de la membrana total de células eucariotas. Este orgánulo está conformado por una bicapa lipídica plegada, que se ramifica en forma de túbulos. Estos están interconectados, de modo que la membrana del RE forma una sola manta que encapsula un solo espacio. Este espacio es conocido como el lumen o espacio cisternal del RE. El lumen del RE ocupa al menos el 10 % del volumen de la célula. La membrana del retículo endoplasmático permite la selección de moléculas para su transporte entre citoplasma y lumen. Puesto que se encuentra directamente conectada a la envoltura nuclear, el RE sirve igualmente como canal de conexión entre el núcleo y el citoplasma.[14]

El RE tiene un rol crucial en la producción, el procesamiento y el transporte de compuestos bioquímicos para su uso dentro y fuera de la célula. En su membrana se lleva a cabo la producción de las proteínas transmembranales, y lípidos que se encuentran en casi todos los orgánulos de la célula, incluyendo al aparato de Golgi, liososmas, endosomas, mitocondrias, peroxisomas, vesículas, membrana plasmática y el RE mismo. Además, casi todas las proteínas que serán transportadas al exterior de la células, así como las proteínas residentes del RE, aparato de Golgi o lisosomas son en primera instancia enviados al lumen del RE. Por esto una gran cantidad de proteínas que se pueden encontrar dentro del lumen del retículo endoplasmático permanecen en él por poco tiempo. Las proteínas que constantemente se pueden encontrar en el lumen se conocen como proteínas residentes del RE. La secuencia de aminoácidos de estas proteínas especiales sirve para la señalización de se retención en el orgánulo. Como ejemplo de estas se puede mencionar la proteína chaperona BiP (Proteína de Unión a Inmunoglobulina, por sus siglas en inglés), la cual identifica a otras proteínas que presentan defectos en su estructura y previene su salida del RE.[15]
El RE también está involucrado en la clasificación cotraduccional de las proteínas. Cuando un polipéptido contiene una señal secuencial del RE que pueda ser reconocida, la producción de la proteína puede detenerse. Una partícula de reconocimiento de señales transporta al polipéptido al la membrana del RE, donde es liberada y posteriormente su producción continua.[16]
En el RE se pueden identificar dos regiones que difieren en estructura y función: El RE liso (REL) y el RE rugoso (RER). El retículo endoplasmático rugoso recibe su nombre debido a que la cara citoplásmica de su membrana está recubierta por ribosomas, dándole un aspecto irregular visto desde un microscopio electrónico. El REL carece de ribosomas, por lo que tiene un aspecto liso.[17]
Funciones del RE liso
En la gran mayoría de las células las regiones lisas en el RE son escasas, y frecuentemente se encuentran intercaladas con el partes rugosas. También se puede conocer como RE transicional, pues contienen sitios de brote de vesículas que transportan proteínas y lípidos recién sintetizados hacia el aparato de Golgi. En algunas células, sin embargo, el REL es abundante y tiene funciones adicionales. El REL de estas células especializadas funciona en diversos procesos metabólicos, incluyendo la síntesis de lípidos, el metabolismo de carbohidratos, y la desintoxicación de fármacos y sustancias tóxicas.[14][17]
Las enzimas de REL son cruciales en el proceso de síntesis de lípidos como aceites, fosfolípidos, y esteroides. En vertebrados, las hormonas sexuales y esteroideas secretadas por las glándulas suprarrenales son un ejemplos de los esteroides sintetizados en el REL. Las células de órganos como este son ricas en REL.[14][17]
Los hepatocitos son otro ejemplo de células especializadas con REL abundante. En estas células se puede ejemplificar el rol que juega el REL en el metabolismo de hidratos de carbono. En hepatocitos se almacena glucógeno, el cual sirve como reserva energética y puede fácilmente convertirse en glucosa. Esto es importante para la regulación de concentraciones de azúcares en sangre. No obstante, el producto primario de la glucogenólisis es la glucosa-1-fosfato. Esta es posteriormente convertida en glucosa-6-fosfato. Una enzima del REL de hepatocitos retira el fosfato de la glucosa de modo que pueda dejar la célula y entrar a sangre.[14][17]
Las enzimas del REL también ayudan a la desintoxicación de drogas, fármacos y sustancias tóxicas. La desintoxicación habitualmente consiste en la adición de un grupo hidroxilo a una droga, incrementando su solubilidad, facilitando así su desecho. Una reacción de desintoxicación que despierta mucho interés es llevada a cabo por la familia de enzimas citocromo P450, la cual cataliza a drogas o metabolitos hidrofóbicos que de otra manera se acumularían en niveles tóxicos en las membranas plasmáticas de las células.[14][17]
Otra función especializada del REL se puede encontrar en células musculares. La membrana del retículo endoplasmático bombea iones de calcio del citosol al espacio cisternal. Cuando una célula muscular es estimulada por impulsos nerviosos el calcio es devuelto a citosol, generando la contracción de la célula.[14][17]
Funciones del RE rugoso
Muchos tipos de células exportan proteínas que son producidas por los ribosomas adheridos al RER. Los ribosomas toman aminoácidos y los ensamblan en unidades proteicas, las cuales son después trasladadas al interior de RE para modificaciones posteriores. Estas pueden sen proteínas transmembranales, las cuales están incrustadas en la membrana del retículo endoplasmático, o bien proteínas hidrosolubles, las cuales son capaces de atravesar la membrana e ingresar al lumen. Las proteínas que ingresan al retículo endoplasmático son plegadas para obtener su estructura tridimensional correcta. Algunas sustancias químicas, como hidratos de carbono o azúcares, son añadidas y una vez terminado este proceso, el RE puede transportarlas a otras partes de la célula donde son necesitadas (proteínas secretoras) o al aparato del Golgi para continuar su modificación.[14][17]
Una vez que las proteínas secretoras son formadas, el RE selecciona las proteínas que deben permanecer en citosol. Las proteínas secretoras salen del RE encapsuladas en vesículas que brotan de la membrana del RE transicional. Estas vesículas se conocen como vesículas transportadoras.[14][17] Una vía alternativa de transporte de lípidos y proteínas al exterior del RE es mediante proteínas transportadoras de lípidos en regiones conocidas como sitios de contacto membranales, donde el RE establece una cercana conexión con las membranas de otros orgánulos, como la membrana plasmática, aparato de Golgi o lisosomas.[18]
Además de la síntesis de proteínas secretoras, el RER forma membranas mediante la adición de proteínas y fosfolípidos. Los polipéptidos que están destinados a ser proteínas membranales son ensamblado en ribosomas. Una vez ensambladas son insertadas a la membrana del RE mismo y son sostenidas por sus porciones hidrofóbicas. El RE rugoso también produce sus propios fosfolípidos membranales, los cuales son sintetizados por enzimas incrustadas en la membrana del RE. La membrana del RE es capaz de expandirse y transferirse a otros componentes del sistema endomembranoso mediante vesículas transportadoras.[14][17]
Aparato de Golgi
-es.svg.png.webp)
El Aparato de Golgi está compuesto por varios compartimentos separados denominados cisternas. Su forma es similar a la de una torre de panqueques. La cantidad de cisternas que un aparato de Golgi puede tener depende de la función específica de la célula que lo contiene. El aparato de Golgi es útil en la modificación postraduccional de proteínas. El lado del aparato Golgi que recibe vesículas del RE es conocido como la cara cis. Esta usualmente se encuentra cercano al RE. La cara opuesta del aparato de Golgi se conoce como la cara trans. Los compuestos modificados dejan al aparato de Golgi por este lado. La mayoría de los compuestos modificados por el aparato son enviados a la membrana plasmática, por lo que la cara trans se encuentra usualmente viendo hacia esta.[19]
Las vesículas liberadas por el RE que contienen proteínas son modificadas en el aparato de Golgi y son preparadas para su secreción hacia el exterior de la célula o hacia otras partes de la misma. Cuando una proteína atraviesa el espacio del aparato de Golgi que está cubierto por enzimas existen muchas posibilidades en cuando a las modificaciones que las proteínas pueden sufrir. Un proceso común es la síntesis o modificación de las porciones de hidrato de carbono en glicoproteínas. El aparato de Golgi es capaz de remover y sustituir monómeros de azúcares, produciendo una gran variedad de oligosacáridos. Además de modificar proteínas, el aparato de Golgi es capaz de manufacturar macromoléculas por sí solo. En células vegetales, el aparato puede producir pectinas y otros polisacáridos que la planta necesita.[20]
Una vez que la modificación termina, el aparato de Golgi clasifica los productos y los envía a varias partes de la célula. Las enzimas del Golgi añaden pequeñas etiquetas para ayudar al reconocimiento de las moléculas. Una vez organizados los productos, el aparato de Golgi los libera brotando vesículas en su cara trans.[21]
Vacuolas
Las vacuolas, al igual que las vesículas, son segmentos de membrana que encierran un espacio dentro de la célula. Son de mayor tamaño que las vesículas y su función específica varía. Las funciones de la vacuolas es diferente para células animales y vegetales.
En células vegetales, las vacuolas ocupan entre 30% y 90% del volumen total de la célula.[22] La mayoría de las células vegetales maduras contienen un vaucola central grande comprendida por una membrana llamada tonoplasto. Las vacuolas de células vegetales funcionan como unidades de almacenamiento para nutrientes y desechos de una célula. Estas moléculas se encuentran disueltas en una sustancia llamada jugo celular. Algunos pigmentos que dan color a las células se pueden encontrar en el jugo celular. Las vacuolas también pueden incrementar el tamaño de una célula mediante la adición de agua. Esto controla la presión de turgencia (la presión osmótica que evita que la pared celular se colapse). Similar a los lisosomas de células animales, las vacuolas tienen un pH acído y contienen enzimas hidrolíticas. El pH de las vacuolas les permite realizar procedimientos de homeóstasis dentro de la célula. Por ejemplo, cuando el pH alrededor de la célula disminuye, los iones de H+ que penetran al citosol pueden ser transferidos a una vacuola para mantener el pH del citosol constante.[23]
En animales, las vacuolas tienen funciones en procesos de exocitosis y endocitosis. La endocitosis se refiere al proceso de adquisición de sustancias al interior de la célula, mientras que en la exocitosis las sustancias son transportadas del interior de la célula al espacio extracelular. El material a importar es encerrado por la membrana plasmática y posteriormente transferido a una vacuola. Existen dos tipos de endocitosis, fagocitosis y pinocitosis. En la fagocitosis las células pueden ingerir partículas grandes como bacterias. La pinocitosis es el mismo proceso, con la diferencia de que las sustancias ingeridas se encuentran en forma de fluido.[24]
Vesículas
Las vesículas son pequeñas unidades encapsuladas por segmentos de membrana que son capaces de transferir moléculas entre diferentes compartimentos. La mayoría de las vesículas transfieren las membranas ensambladas en el retículo endoplasmático hacia el aparato de Golgi, el cual las transporta a diferentes lugares.[25]
Existen diferentes tipos de vesículas, cada uno con una configuración diferente de proteínas. La mayoría de las vesículas se encuentran formados por regiones específicas de membranas. Cuando una vesícula brota de una membrana, esta contiene proteínas específicas en su cara citosólica. Cada membrana que recibe una vesícula tiene igualmente moléculas de reconocimiento en su cara citosólica. Estas moléculas son correspondientes a las proteínas de las vesículas que viajan hacia una membrana. Una vez que la vesícula llega a su destino se fusiona con la membrana.[26]
Existen tres tipos de vesículas conocidas: cubiertas por clatrina, por el complejo proteico COP I, o por el complejo proteico COP II. Cada tipo tiene funciones específicas en la célula. Por ejemplo, las vesículas cubiertas por clatrina transportan sustancias entre el aparato de Golgi y la membrana plasmática. Las vesículas cubiertas por COP I y COP II frecuentemente se utilizan para el transporte entre el RE y el aparato de Golgi.[26]
Lisosomas
Los lisosomas son orgánulos que contienen enzimas hidrolíticas utilizadas en la digestión intracelular. Las funciones principales de un lisosoma son el procesamiento de moléculas ingeridas por la célula, así como el reciclaje de partes celulares obsoletas. Las enzimas en el interior de los lisosomas son hidrolasas ácidas, las cuales requieren un ambiente ácido para su funcionamiento óptimo. Los lisosomas conservan un ambiente de estas características al mantener un pH de 5.0 en su interior.[27] En caso de ruptura de un lisosoma, las enzimas liberada no funcionarían correctametne, pues el pH del citosol es neutro. No obstante, si grandes cantidades de lisosomas se rompen la célula puede ser destruida por autodigestión.
Los lisosomas llevan a cabo la digestión intracelular en un proceso llamado fagocitosis (del griego phagein, comer y kytos, vesícula, en este caso la célula), en el cual se fusiona con una vacuola, liberando sus enzimas dentro de ella. Mediante este proceso, las azúcares, aminoácidos y otros monómeros ingresan al citosol y sirven de nutrientes para la célula. Los lisosomas también ocupan sus enzimas hidrolíticas para reciclar a los orgánulos obsoletos de la célula en un proceso denominado autofagia. El lisosoma ingiere a otro orgánulo y utiliza sus enzimas para degradar el material ingerido. Los monómeros orgánicos resultantes son transferidos a citosol para ser utilizados nuevamente. La última función de un lisosoma es la digestión de la célula misma a través del proceso de autólisis.[28]
Spitzenkörper
El spitzenkörper es un componente del sistema endomembranoso encontrado únicamente en hongos, y está asociado con el crecimiento de punta de hifa. Es un cuerpo de fase oscura compuesto por una agregación de vesículas rodeadas por membrana que contienen componentes de la pared celular. Sirve como punto de ensamblaje y liberación de estos componentes entre el aparato de Golgi y la membrana celular. El spitzenkörper es capaz de moverse y su desplazamiento genera nuevo crecimiento de puntas de hifa.
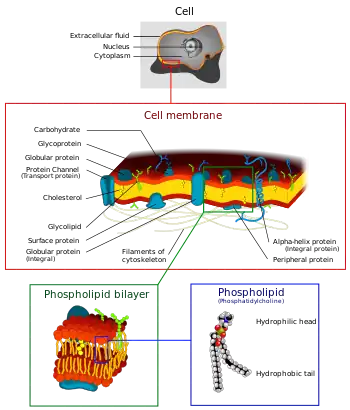
Membrana plasmática
La membrana plasmática es una membrana formada por una bicapa de fosfolípidos que separa al contenido celular del ambiente que la rodea, y regula el transporte de moléculas y señales hacia el interior y exterior de la célula. Tiene incrustadas proteínas que realizan diversas de sus funciones. La membrana plasmática no tiene una estructura fija ni rígida, las moléculas que la componen son capaces de realizar movimientos laterales. Estos movimientos, junto con los múltiples componentes que la conforman son la razón por la que se refiere a ella como "mosaico fluido". Moléculas pequeñas como dióxido de carbono, agua y oxígeno pueden atravesar la membrana plasmática libremente mediante difusión u ósmosis. Las moléculas de mayor tamaño que la célula necesita son ingresadas con la ayuda de proteínas mediante el transporte activo.[29]
La membrana plasmática de una célula tiene múltiples funciones. Estas incluyen el transporte de nutrientes a interior de la célula, el desecho de residuos metabólicos, la prevención del ingreso de materiales no deseados, la prevención de la salida de materiales que la célula necesita, la conservación del pH y presión osmótica del citosol. Las proteínas transportadoras se utilizan para cumplir estas funciones, pues permiten el paso selectivo de materiales hacia el interior de la célula. Estas proteínas utilizan la hidrólisis de ATP para bombear materiales en contra de su gradiente de concentración.[29]
Además de estas funciones universales, la membrana plasmática tiene un rol específico en organismos multicelulares. Las membranas contienen glucoproteínas que ayudan al reconocimiento de otras celulares, de modo que puedan intercambiar metabolitos y formar tejidos. Otras proteínas de la membrana plasmática permiten la unión con el citoesqueleto y la matriz extracelular. Estas funciones ayudan a mantener la forma de la célula y fijan la ubicación de las proteínas membranales. También se pueden encontrar enzimas ctalizadoras de reacciones en la membrana. Las proteínas receptoras en la membrana tiene una forma específica que depende del mensajero químico que lo active. Esto resulta en diferentes respuestas celulares.[30]
Evolución
El origen del sistema endomembranoso está relacionado con el origen de las células eucariotas mismas y este a su vez con el origen endosimbiótico de las mitocondrias. Se han propuesto muchos modelos para la explicación de su origen (vistos en [31]). El concepto más reciente sugiere que el sistema endomembranoso evolucionó de vesículas secretadas por la mitocondria endosimbiótica.[32] Este modelo, basado en vesículas de membrana externa, es actualmente el que requiere menos modificaciones a la teoría del origen de células eucariotas y explica las muchas conexiones que tienen las mitocondrias con otros compartimentos celulares.[33]
Véase también
Referencias
- Davidson, Michael (2005). «Plasma Membrane». Molecular Expressions. Florida State University. Consultado el 9 de diciembre de 2008.
- Davidson, Michael (2005). «The Nuclear Envelope». Molecular Expressions. Florida State University. Consultado el 9 de diciembre de 2008.
- Davidson, Michael (2005). «The Endoplasmic Reticulum». Molecular Expressions. Florida State University. Consultado el 9 de diciembre de 2008.
- Graham, Todd R. (2000). Eurekah Bioscience Collection Cell Biology. University of New South Wales and Landes Bioscience. ISBN 0-7334-2108-3.
- Lodish, Harvey (2000). «Section 5.4 Organelles of the Eukaryotic Cell». Molecular Cell Biology. W. H. Freeman and Company. Consultado el 9 de diciembre de 2008.
- Cooper, Geoffrey (2000). «The Mechanism of Vesicular Transport». The Cell: A Molecular Approach. Sinauer Associates, Inc. Consultado el 9 de diciembre de 2008.
- Smith, A. L. (1997). Oxford dictionary of biochemistry and molecular biology. Oxford [Oxfordshire]: Oxford University Press. pp. 206. ISBN 0-19-854768-4.
- Campbell, Neil A.; Reece, Jane B. (2002). Biology (6th edición). Benjamin Cummings. ISBN 0-8053-6624-5.
- Bryant DA, Frigaard NU (2006). «Prokaryotic photosynthesis and phototrophy illuminated». Trends Microbiol. 14 (11): 488-96. PMID 16997562. doi:10.1016/j.tim.2006.09.001.
- Psencík J, Ikonen TP, Laurinmäki P (August 2004). «Lamellar Organization of Pigments in Chlorosomes, the Light Harvesting Complexes of Green Photosynthetic Bacteria». Biophys. J. 87 (2): 1165-72. PMC 1304455. PMID 15298919. doi:10.1529/biophysj.104.040956. Archivado desde el original el 10 de mayo de 2020. Consultado el 2 de marzo de 2018.
- Childs, Gwen V. (2003). «Nuclear Envelope». UTMB. Archivado desde el original el 20 de junio de 2006. Consultado el 28 de septiembre de 2008.
- Cooper, Geoffrey (2000). «The Nuclear Envelope and Traffic between the Nucleus and Cytoplasm». The Cell: A Molecular Approach. Sinauer Associates, Inc. Consultado el 9 de diciembre de 2008.
- Alberts, Walter (2002). «Nuclear Pore Complexes Perforate the Nuclear Envelope». Molecular Biology of the Cell 4th edition. Garland Science. Consultado el 9 de diciembre de 2008.
- Cooper, Geoffrey (2000). «9:Protein Sorting and Transport - The Endoplasmic Reticulum, Golgi Apparatus, and Lysosomes». The Cell: A Molecular Approach (en inglés). Sinauer Associates, Inc. Consultado el 9 de diciembre de 2008.
- Bertolotti, Anne; Zhang, Yuhong; Hendershot, Linda M.; Harding, Heather P.; Ron, David (2000). «Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response». Nature Cell Biology 2 (6): 326-333. PMID 10854322. doi:10.1038/35014014. Consultado el 3 de octubre de 2008.
- Mason K.A.; Losos J.B.; Singer S.R.; Raven P.H. (2011). Biology. McGraw Hill education. pp. Fluid mosaic 89.
- Alberts, Walter (2002). «Membrane-bound Ribosomes Define the Rough ER». Molecular Biology of the Cell 4th edition. Garland Science. Consultado el 9 de diciembre de 2008.
- Levine T, Loewen C (August 2006). «Inter-organelle membrane contact sites: through a glass, darkly». Curr. Opin. Cell Biol. 18 (4): 371-8. PMID 16806880. doi:10.1016/j.ceb.2006.06.011.
- Rothman, J. (1981). «The golgi apparatus: two organelles in tandem». Science 213 (4513): 1212-1219. PMID 7268428. doi:10.1126/science.7268428. Consultado el 4 de octubre de 2008.
- Alberts, Walter (2002). «Transport from the ER through the Golgi Apparatus». Molecular Biology of the Cell 4th edition. Garland Science. Consultado el 9 de diciembre de 2008.
- Cooper, Geoffrey (2000). «The Golgi Apparatus». The Cell: A Molecular Approach. Sinauer Associates, Inc. Consultado el 9 de diciembre de 2008.
- Alberts, Walter (2002). «Plant and Fungal Vacuoles Are Remarkably Versatile Lysosomes». Molecular Biology of the Cell 4th edition. Garland Science. Consultado el 9 de diciembre de 2008.
- Lodish, Harvey (2000). «Plant Vacuoles Store Small Molecules and Enable the Cell to Elongate Rapidly». Molecular Cell Biology. W. H. Freeman and Company. Consultado el 9 de diciembre de 2008.
- Cooper, Geoffrey (2000). «Endocytosis». The Cell: A Molecular Approach. Sinauer Associates, Inc. Consultado el 9 de diciembre de 2008.
- Lodish, Harvey (2000). «Section 17.10 Molecular Mechanisms of Vesicular Traffic». Molecular Cell Biology. W. H. Freeman and Company. Consultado el 9 de diciembre de 2008.
- Alberts, Walter (2002). «The Molecular Mechanisms of Membrane Transport and the Maintenance of Compartmental Diversity». Molecular Biology of the Cell 4th edition. Garland Science. Consultado el 9 de diciembre de 2008.
- Alberts, Walter (2002). «Transport from the Trans Golgi Network to Lysosomes». Molecular Biology of the Cell 4th edition. Garland Science. Consultado el 9 de diciembre de 2008.
- Cooper, Geoffrey (2000). «Lysosomes». The Cell: A Molecular Approach. Sinauer Associates, Inc. Consultado el 9 de diciembre de 2008.
- Cooper, Geoffrey (2000). «Structure of the Plasma Membrane». The Cell: A Molecular Approach. Sinauer Associates, Inc. Consultado el 9 de diciembre de 2008.
- Lodish, Harvey (2000). «Section 5.3. Biomembranes: Structural Organization and Basic Functions». Molecular Cell Biology. W. H. Freeman and Company. Consultado el 9 de diciembre de 2008.
- Martin WF (6 de mayo de 2014). «Endosymbiotic theories for eukaryote origin». Philos. Trans. R. Soc. B 370: 20140330-18. PMC 4571569. PMID 26323761. doi:10.1098/rstb.2014.0330.
- Gould SB (31 de marzo de 2016). «Bacterial Vesicle Secretion and the Evolutionary Origin of the Eukaryotic Endomembrane System». Trends Microbiol. 24: 525-34. PMID 27040918. doi:10.1016/j.tim.2016.03.005.
- Murley, A.; Nunnari, J. (3 de marzo de 2016). «The emerging network of mitochondria-organelle contacts». Mol. Cell 61: 648-653. PMID 26942669. doi:10.1016/j.molcel.2016.01.031.
| Control de autoridades |
|
|---|
 Datos: Q899827
Datos: Q899827 Multimedia: Endomembrane system / Q899827
Multimedia: Endomembrane system / Q899827