Allosaurus
Allosaurus (gr. "lagarto extraño") es un género extinto con cuatro especies válidas y varias otras posibles de dinosaurios terópodos alosáuridos, que vivieron a finales del período Jurásico, hace aproximadamente 161,2 y 145 millones de años, entre el Oxfordiense y el Titoniense, en lo que hoy es Norteamérica y Europa.[1] Se han descrito varias especies, el número de especies válidas es dudoso y depende del autor. La más conocida y la especie tipo es Allosaurus fragilis de Estados Unidos descrita en el siglo XIX, luego de esta gran cantidad de especies e incluso géneros, que fueron propuestos por todo el globo sin que ninguna termine siendo totalmente aceptada. En 2006 en Portugal se describió Allosaurus europaeus, aunque solo fue separada tentativamente dada la localización geográfica y Allosaurus jimmadseni de EE. UU. y la tercera especie de este último país, Allosaurus lucasi, en 2014. Sin embargo, todas ellas han sido disputadas.[2]
| Allosaurus | ||
|---|---|---|
| Rango temporal: 161,2 Ma - 145 Ma Jurásico Superior | ||
 Reconstrucción de un esqueleto de Allosaurus en el Museo de Historia Natural de San Diego | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| Infraorden: | Carnosauria | |
| (sin rango): | Tetanurae | |
| Superfamilia: | Allosauroidea | |
| Familia: | Allosauridae | |
| Género: |
Allosaurus Marsh, 1877 | |
| Especie tipo | ||
|
Allosaurus fragilis Marsh, 1877 | ||
| Otras Especies | ||
| ||
| Sinonimia | ||
| ||
Los fósiles tipo fueron descritos en 1877 por el paleontólogo Othniel Charles Marsh. Su naturaleza pobre llevó a que en la primera monografía detallada sobre un espécimen relativamente completo de Allosaurus USNM 4734, este se describiera como Antrodemus valens,[3] hoy considerada inválida por el principio de prioridad de ICZN.[4] Esto ha llevado a peticiones para un nuevo ejemplar tipo que sustituya al original, como UUVP 6000 (=DINO 2560) o el paratipo USNM 4734. Como uno de los primeros dinosaurios terópodos conocidos, ha atraído mucho la atención fuera de los círculos paleontológicos. De hecho, ha sido representado en varias películas y documentales sobre la vida prehistórica.
A diferencia de otros terópodos anteriores, los Alosáuridos eran comparativamente más grandes, con fémures de más de 90 centímetros de largo para A.fragilis,[4] y hasta 113.5 cm para Allosaurus maximus, que sugieren longitudes máximas entre 8,5 y 10,5 metros.[5] Sin embargo comparten rasgos anatómicos con otros terópodos más basales. Era un carnívoro bípedo con garras y patas masivas, balanceadas por su pesada cola. Su cráneo curvo provisto de dientes aserrados poseía además unas crestas bajas irregulares, sobre y delante de los ojos. A pesar de haber sido un temible cazador relativamente grande no pesaba tanto, siendo comparable en masa a un moderno rinoceronte indio. Se lo conoció por más de la mitad del siglo XX como Antrodemus, pero el estudio de los abundantes restos de la cantera dinosaurio Cleveland-Lloyd llevó al nombre "Allosaurus" a volver a ser mayoritariamente usado, y se estableció como uno de los dinosaurios más conocidos.
Compartió el hábitat con varios saurópodos gigantescos tales como Apatosaurus, Brontosaurus, Barosaurus, Diplodocus, Camarasaurus, Brachiosaurus y Haplocanthosaurus así como con otros herbívoros tales como Stegosaurus, Dryosaurus y Camptosaurus, los cuales debieron haber sido presa potenciales. Fue el depredador grande más común en lo que hoy es Norteamérica, de hecho es famoso por ser el dinosaurio carnívoro más abundante y común en el registro fósil con cerca de 60 especímenes y unas cuantas docenas de ellos bien preservados. El gran número de Allosaurus encontrado en un mismo sitio hace pensar que tenían una cierta vida en grupo.
Descripción


Allosaurus es un gran terópodo típico, con un cráneo grande y un cuello corto, una cola larga y miembros superiores reducidos. Allosaurus fragilis, la especie mejor conocida, presenta un individuo adulto muy bien preservado, USNM 4734, con las escápulas firmemente fusionadas con sus coracoides, y todas las vértebras sacras fusionadas, ambos signos de adultez en dinosaurios. La longitud de este espécimen se calcula en unos 7.4 metros de longitud, con un fémur de 85 centímetros de largo.[3][6] Otro espécimen famoso, MOR 693, conocido como "Big Al", tiene longitud de 7.57 metros, medida a partir del esqueleto montado escaneado.[7] Un esqueleto exquisitamente preservado, SMA 005, llamado "Big Al" 2, presenta todos los huesos presentes excepto unos pocos huesos pequeños, midiendo 7,6 metros de largo con un cráneo de 79 centímetros. [8] El espécimen relativamente completo más grande descubierto en Cleveland LLoyd, UUVP 6000, tiene un cráneo de 845 milímetros de largo, un fémur de 88 centímetros y una longitud estimada desde 7,9 hasta 8,5 metros de longitud ,[4][6][5] a pesar de que la cola nunca fue encontrada, salvo la primera caudal. Este espécimen ha sido estimado con un peso de una a 1.7 toneladas[5] El ejemplar con el mayor fémur conocido de 973 milímetros de largo y 381 milímetros circunferencia, AMNH 680 podría haber superado una masa de 2.65 t[9] y haber alcanzado una longitud superior a 9 metros, hasta 9,7 metros.[10] El rango de peso propuesto para el género ha variado entre 1000 a 4000 kilogramos, teniendo en cuenta todos los especímenes, tanto Allosurus fragilis como la posible segunda especie Allosaurus (=Saurophaganax?) maximus y métodos tanto alométricos como volumétricos.[9][5] [11][12] A partir del espécimen apodado "Big Al", los investigadores utilizando modelos informáticos, llegaron a una mejor estimación de 1500 kilogramos para el individuo, pero variando los parámetros se encontraron con un rango de aproximadamente 1400 kilogramos a 2000 kilogramos.[13]
En su monografía de 1976 para Allosaurus, James Madsen menciona un tamaño máximo, según algunos huesos, de entre 12 a 13 metros.[4] Esto se debe a que varios especímenes gigantes han sido históricamente referidos a Allosaurus, pero en realidad pertenecen a otros géneros. El cercano Saurophaganax, OMNH 1708, habría alcanzado quizás los 10,9 metros de largo,[10] siendo este único espécimen incluido dentro de Allosaurus como Allosaurus maximus, aunque recientes estudios apoyan la hipótesis de dos géneros separados.[14] Existe otro potencial Allosaurus gigante, asignado al género Epanterias, AMNH 5767, que fue originalmente descrito como saurópodo por Cope, y asignado a los alosauroideos por Osborn y Mook. La vértebra dorsal 2 mide 115 mm de largo,[15] comparada con la vértebra dorsal de USNM 4734, de 88 mm de largo[16] y 7.4 m de largo total,[17] sugiere un individuo de 9,8 m de largo.[10] El descubrimiento más reciente es un esqueleto parcial de la mina de Peterson en las rocas de Morrison de Nuevo México, pero este alosauroide grande puede ser otro individuo de Saurophaganx,[18] de aprox. 9,9 metros de longitud.[10]
Cráneo


El cráneo y los dientes de Allosaurus tenían un tamaño modesto si lo comparamos con otros terópodos de su tamaño. A pesar de su tamaño, se ha encontrado una paradoja en su constitución: un cráneo fuerte con una mordida débil. Es por eso que los científicos creen que la forma de ataque de un Allosaurus era acechar a su presa y luego asestarle un fuerte golpe de mandíbulas moviendo su cabeza de arriba abajo como un hacha, aunque esto ha sido desmentido, ya que solo causaría daño con los dientes posteriores y no con los premaxilares, o los de las mejillas, por lo que se cree que este mordía y sacudía su cabeza hacia el lado opuesto de la presa y arrancar un gran pedazo de carne usando sus aserrados dientes. El paleontólogo Gregory S. Paul ha estimado un tamaño de 845 milímetros para el cráneo de un individuo de 7,9 metros de largo.[17] Cada premaxilar, los huesos que forman la punta del hocico, tenían cinco dientes con una sección al corte con forma de D y cada maxilar, los principales huesos con dientes en la mandíbula superior tenían entre 14 y 17 dientes, el número de dientes no se corresponde exactamente con el tamaño del hueso. Cada dentario, el hueso que soporta los dientes de la mandíbula inferior tenían entre 14 y 17 dientes, con un promedio de 16. Los dientes se volvieron más cortos, estrechos y curvados hacia la parte posterior del cráneo. Todos los dientes tenía bordes aserrados. Estos se perdían con facilidad, y fueron reemplazados continuamente, por lo que son los fósiles más comunes.[4] A diferencia de otros terópodos, los alósauridos tenían un par de abultamientos, similares a "cuernos" embotados, formados por extensiones del hueso lagrimal[4] y dos líneas paralelas de pequeñas protuberancias óseas sobre el hueso nasal, al tope de la cabeza, sobre y delante de los ojos, teniendo gran variedad de formas y tamaños.[4] Estas ornamentaciones probablemente estuvieran cubiertas de queratina lo que habría aumentado su tamaño. Se ha argumentado que poseían gran variedad de funciones, incluyendo que funcionaban para proteger del sol los ojos,[4] para exhibiciones, y para combates entre individuos de la misma especies[19][17] aunque eran muy frágiles.[4] El cráneo también poseía un largo hocico y amplias fenestras que reducían el peso de la cabeza proporcionando áreas para la atadura de músculos y órganos sensoriales. Sus mandíbulas contenían cerca de 60 dientes afilados con forma de D en borde transversal, los cuales le hubieran ayudado a cazar presas y devorar carroña. El reborde óseo que tenían en la parte posterior de la cabeza era fuerte y rugoso, sirviendo de inserción muscular, también ha sido visto dentro de los tiranosáuridos.[17]
Dentro de los huesos lacrimales tenía unas depresiones para contener una glándula, similar a la glándula de sal.[20] Con los senos maxilares mejor desarrollados que en terópodos basales como Ceratosaurus y Marshosaurus, probablemente estaban relacionados con el sentido del olfato, probablemente sosteniendo un órgano vomeronasal. El techo del cráneo era fino, quizás para mejorar la termorregulación del cerebro.[4] En las mandíbula, los huesos del frente y la mitad trasera se articularon libremente, permitiendo que las quijadas se arqueen hacia fuera y aumentando la apertura de la boca.[21] El techo del cráneo y los frontales también tenían ese tipo de unión.[4]
Esqueleto postcraneal

Allosaurus tenía nueve vértebras en el cuello, 14 en la espalda, y cinco en el sacro donde apoya la cadera.[22] El número de vértebras de la cola es desconocido y varía con el tamaño individual. James Madsen estima alrededor de 50,[4] mientras que Gregory S. Paul considera que este número es demasiado alto y sugirió 45 o menos.[17] En las vértebras del cuello y la porción anterior de la espalda había espacios huecos.[4] Estos espacios, que se encuentran también en los terópodos modernos, es decir, las aves, son interpretados como ocupados por los sacos aéreos que se usan en la respiración.[23]
La caja costal era amplia, dándole un pecho con forma de barril, especialmente con respecto a los menos derivados terópodos como Ceratosaurus.[24] El Allosaurus tenía gastralias (costillas abdominales), pero no son comunes de encontrar,[4] debido a que se fosilizaban muy pobremente.[17] En un caso publicado, las gastralias muestran lesiones durante la vida.[25] La fúrcula estaba presente pero recién fue reconocida en 1996, en algunos casos se la había confundido con gastralias.[26][25] El ilion, el hueso principal de la cadera, era grande y fuerte, y el pubis tenía un saliente prominente que se pudo haber utilizado para la inserción de músculos y como apoyo para reclinar el cuerpo en la tierra. Madsen observó que alrededor de la mitad de los individuos de la mina de dinosaurios de Cleveland Lloyd, independientemente del tamaño, tenían pubis que no se había fusionado al otro en los extremos de la saliente. Se ha sugerido que esto es una característica sexual, donde las hembra carecieron de la fusión para poder poner los huevos más fácilmente.[4] Esta propuesta no ha atraído la atención del medio científico.

A pesar de que los miembros delanteros eran cortos en comparación a los miembros traseros, estos eran masivos y con garras parecidas a las del águila. El primer metacarpiano de cada "mano" es corto y robusto, y se encuentra girado lateralmente, lo que provocaba que su dedo correspondiente se dirigiese hacia los otros dos al cerrar la mano. El esqueleto del alosaurio, como otros terópodos, exhibía características de ave, así como la espoleta[27][28] y vértebras huecas del cuello con sacos aéreos, que usaba en la respiración.[23] Los brazos de los alosaurios eran cortos en comparación a sus patas traseras, solo el 35% del largo de estas en un ejemplar adulto.[29] Teniendo tres dedos por mano, terminados en unas garras, grandes y fuertes y curvadas.[4] Sus miembros superiores eran poderosos,[17] con el antebrazo más corto que el brazo (una relación 1:1.2; entre húmero y ulna).[30] Este tenía una versión del hueso semilunar en el carpo[31] cosa encontrada en los terópodos más derivados como en los maniraptores. Uno de esos dedos, el del medio, era más largo,[17] y se separaba de los otros.[30] Las patas no eran tan largas, o preparadas para la carrera como en los tiranosáuridos, y las garras de los dedos de los pies estaban menos desarrolladas y se parecían más a pezuñas que en los terópodos más antiguos.[17] Cada pie tenía tres fuertes dedos que contactaban el piso y uno, como un garrón que Madsen sugirió se habría podido utilizase para agarrar en los jóvenes.[4] También se ha interpretado que podía tener un remanente de un quinto metatarsal, que serviría como palanca entre el tendón de Aquiles y el pie.[32]
Descubrimiento e investigación
Primeros descubrimientos

El estudio del descubrimiento y los primeras descripciones de Allosaurus se complica por la multiplicidad de nombres acuñados durante la Guerra de los Huesos de finales del siglo XIX. En 1869 los nativos de Middle Park, cerca de Granby, Colorado, hallaron una vértebra caudal incompleta, que fue entregada al geólogo Ferdinand Vandiveer Hayden, quien pensó que era la pezuña fosilizada de un caballo prehistórico. En 1870, el paleontólogo Joseph Leidy la estudió debidamente y asignó la vértebra al género Poekilopleuron como Poicilopleuron [sic] valens, pero al notar la diferencia que esta vértebra tenía con las demás halladas hasta el momento, Leidy propuso que el fósil pertenecía a un nuevo género, al cual lo nombró Antrodemus,[33] sin embargo no fue un género formalmente descrito. Fue ya en 1877 que el célebre paleontólogo Othniel Charles Marsh le dio el nombre formal de Allosaurus al género, y A. fragilis a la especie tipo,[34] basado en un material mucho mejor, incluyendo un esqueleto parcial, descubiertos en Garden Park, al norte de Canyon City, Colorado. Marsh y Edward Drinker Cope, en su competencia científica, acuñaron varios géneros distintos basados en escaso material similar que posteriormente fueron colocados dentro de la taxonomía de Allosaurus. Estos incluyen los Creosaurus[35] y Labrosaurus[36] ambos por O. C. Marsh y Epanterias de E. D. Cope.[37] Allosaurus está basado en el holotipo YPM 1930, una pequeña colección de huesos que incluyen tres vértebras, fragmentos de costilla, un diente, un hueso del pie, a y lo más usado en posteriores discusiones, el eje del húmero derecho.[38][39] Con ansias de encontrar muchos fósiles y mantener los lugares de descubrimiento en secreto, Cope y Marsh no presentaban trabajos de sus descubrimientos, más comúnmente sucedió, esto se hizo por medio de sus subordinados. Por ejemplo, después del hallazgo de Benjamin Mudge del espécimen tipo de Allosaurus en Colorado, Marsh eligió concentrar sus esfuerzos en Wyoming;, cuando reasumió el trabajo en Garden Park en 1883, M. P. Felch encontró un ejemplar completo de Allosaurus más varios esqueletos parciales.[20] Además de esto, uno de los recolectores de Cope, H. F. Hubbell, encontró los especímenes de Como Bluff área de Wyoming en 1879, pero aparentemente no lo mencionó por completo, y Cope nunca los desenterró. Fueron recogidos en 1903, varios años después de la muerte de Cope, encontrándose uno de los esqueletos más completos conocidos hasta hoy, y en 1908 el esqueleto, hoy catalogado como AMNH 5753, fue expuesto al público.[40] Este es el montaje bien conocido inclinado sobre un esqueleto parcial de Apatosaurus como si estuviese alimentándose, ilustrado como Charles R. Knight Es el primer montaje libre de un dinosaurio terópodo, e ilustrado y fotografiado a menudo, aunque nunca fuese descrito científicamente.[41]

La multiplicidad de nombres tempranos ha complicado estudios posteriores, con la situación aún más complicada por las descripciones concisas que proporcionaron Marsh y Cope. Durante mucho tiempo autores como Samuel Wendell Williston sugirieron que existían muchos nombres para el mismo animal.[42] Por ejemplo, Williston apunta que en 1901 que Marsh nunca distinguió adecuadamente Allosaurus de Creosaurus.[43] La tentativa temprana más influyente de arreglar la situación fue producida por Charles W. Gilmore en 1920. Él llegó a la conclusión que la vértebra de la cola de ' ' Antrodemus' ' descrita por Leidy era indistinguible de las de Allosaurus, y así Antrodemus debe ser el nombre preferido porque como el más viejo nombre tenía prioridad.[30] Antrodemus se convirtió el nombre aceptado para este género por más de cincuenta años, hasta que James Madsen publicara los especímenes de Cleveland-Lloyd y concluyó que Allosaurus se debe utilizar ya que Antrodemus fue basado en el material con pobres características de diagnóstico y la pobre información del lugar de procedencia,(por ejemplo, formación geológica de Antrodemus es desconocida).[4] "Antrodemus" se ha utilizado informalmente para al distinguir entre el cráneo restaurado por Gilmore y el cráneo compuesto restaurado por Madsen. [44]
Significado del nombre
Allosaurus recibió su nombre en 1877 por Othniel Charles Marsh. El nombre del género está compuesto de las palabras griegas αλλος (allos, "extraño") y σαυρος (sauros, "lagarto"),[45] en referencia a su vértebra diferente a la de todos los dinosaurios conocidos hasta ese momento y a su naturaleza reptiliana.[38][39] El epíteto de la especie fragilis proviene del término en latín para "frágil", refiriéndose a la ligera constitución de la vértebra.

Descubrimientos de Cleveland-Lloyd
Una famosa "cama fósil" puede encontrarse en la Mina del Dinosaurio de Cleveland Lloyd, en Utah. Esta cama fósil contiene cerca de 10 000 huesos, sobre todo de Allosaurus, entrelezados con restos de otros dinosaurios, como el Stegosaurus y Ceratosaurus. Es aún un misterio como los fósiles de tantos animales pueden hallarse en un solo lugar. El cociente de fósiles de animales carnívoros sobre los de herbívoros es normalmente muy pequeño. Descubrimientos como estos pueden indicar que el alosaurio cazaba en grupo, sin embargo esto es muy difícil de probar. Algo verdaderamente más posible es que el sitio de Cleveland Lloyd haya formado una "trampa de depredadores" (por ejemplo arenas movedizas), similar a Rancho La Brea, lo cual causó que un gran número de predadores hayan sido atraídos por la carne de herbívoros y luego capturados en un sedimento ineludible.Cleveland-Lloyd Dinosaur Quarry, Utah - The Predator Trap - American West Travelogue
Aunque aparecía de forma esporádica en trabajos desde 1927 la Mina de Dinosaurio de Cleveland-Lloyd en Condado de Emery, Utah y el lugar en sí mismo fue estudiado por William J. Stokes en 1945,[46] las mayores operaciones no comenzaron hasta 1960. En un esfuerzo cooperativo de unas 40 instituciones, cientos de huesos fueron recobrados entre 1960 y 1965.[4] La mina es notable para el predominio de Allosaurus, con una excelente condición de los especímenes, y la carencia una explicación científica totalmente aceptada de cómo sucedió esta acumulación. La mayoría de los grandes terópodos son Allosaurus fragilis (por lo menos 46 A. fragilis, en un mínimo de 73 dinosaurios), y los fósiles se encontraron desarticulados y mezclados. Casi una docena de trabajos científicos se han escrito en tafonomía del sitio, presentando numerosas explicaciones contradictorias de su formación. Las sugerencias van desde que los animales habrían quedado atrapados en un pantano, en una trampa de fango, víctimas de una sequía muriendo alrededor de una charca que filtraba.[47] Sin importar la causa real, la gran cantidad de bien preservados Allosaurus han permitido que este género sea conocido detalladamente, haciéndolo entre los terópodos más conocidos. Los restos esqueléticos de la mina pertenecen a los individuos de casi todas las edades y tamaños, de 3 metros de largo[48] a 12 metros y lo disarticulado es una ventaja para describir huesos, generalmente encontrados fundidos.[4]
Descubrimiento de 1980 hasta el presente
El periodo desde el trabajo de Madsen ha sido marcado por la gran cantidad de trabajo que hicieron foco en la vida del Allosaurus (sobre la paleobiolgía y paleoecología). Tales estudios han cubierto asuntos incluyendo la variación esquelética ,[49] desarrollo,[50][51] conformación del cráneo,[52] métodos de caza,[53] el cerebro,[54] y la posibilidad de vida gregaria y del cuidado de las crías por los padres.[55] El nuevo análisis de material viejo (particularmente de los especímenes más grandes),[17][56] nuevos descubrimientos en Portugal,[57] varios nuevos ejemplares completos encontrados[58][25][59] cosas que hicieron aumentar el conocimiento de este dinosaurio.
"Big Al" y "Big Al 2"
Uno de los más significativos hallazgo de Allosaurus encontrados en 1991 fue el descubrimiento de "Big Al", MOR 693,[60] un espécimen completo en un 95% y parcialmente articulado, que mide alrededor de 7.57 metros de largo.[7] Tiene 19 huesos rotos con señales de infección, lo cual probablemente contribuyó a su muerte. [61] MOR 693 fue excavado cerca de Shell, Wyoming, por un equipo del Museo de las Rocallosas y la Universidad de Wyoming.[62] Este esqueleto fue descubierto por un equipo suizo, liderado por Kirby Siber. Este mismo equipo excavó un segundo espécimen deAllosaurus, "Big Al Dos", el cual es el mejor esqueleto preservado de su clase hasta la fecha, y pertenece a un adulto.[59] [8]
Lo completo, la preservación, y la importancia científica de este esqueleto dieron a "Big Al", su nombre, el individuo en sí mismo estaba debajo del tamaño medio para Allosaurus fragilis[62] y era un subadulto que habría alcanzado un 87% del tamaño adulto.[63] El espécimen lo describió Breithaupt en 1996.[58] Diecinueve de sus huesos estaban quebrados o mostraban signos de infección, que pudieron haber contribuido la muerte de "Big Al". Las patologías de los huesos incluían cinco costillas, cinco vértebras, y cuatro huesos de los pies; varios huesos dañados mostrando osteomielitis, una infección del hueso. Un problema particular para el animal vivo era infección y trauma al pie derecho que probablemente afectaba el movimiento y pudo también haber predispuesto el otro pie a lesión debido a un cambio en el paso.[63] También se describieron múltiples lesiones en "Big Al 2".[64]
Clasificación

Allosaurus es un miembro de la familia Allosauridae de grandes terópodos dentro del gran grupo Carnosauria. El nombre de la familia fue creado a partir de este género en 1878 por Othniel Charles Marsh,[35] pero el término cayó en desuso hasta los años de 1970 en favor de Megalosauridae, otra familia de grandes terópodos que se convertiría en un taxón cajón de sastre. Esto junto con el uso de Antrodemus por Allosaurus durante el mismo periodo, es un punto que es necesario recordar al revisar la información sobre Allosaurus en publicaciones anteriores al trabajo de James Madsen en 1976. La mayor parte de la publicaciones que usaron el nombre de Megalosauridae en vez de Allosauridae incluye trabajos de Gilmore, 1920,[30] von Huene, 1926,[65] Romer, 1956 y 1966,[66][67] Steel, 1970,[68] y Walker, 1964.[69]
Luego del influyente trabajo de Madsen, Allosauridae se convirtió en la familia preferida para clasificarlo, pero esta no se encontraba fuertemente definida. Durante ese periodo se usó una gran variedad de grandes terópodos para definir Allosauridae, usualmente los más grandes y mejor conocidos que los megalosáuridos. Los típicos terópodos con los que se relacionó a Allosaurus incluyen a Indosaurus (un abelisáurido), Piatnitzkysaurus (un tetanuro basal), Piveteausaurus (un megalosáurido), Yangchuanosaurus (un metriacantosáurido),[70] Acrocanthosaurus (un carcarodontosáurido), Chilantaisaurus (un espinosáurido), Compsosuchus (un abelisáurido), Stokesosaurus (un tiranosauroide basal), y Szechuanosaurus (un metriacantosáurido).[71] Con el moderno conocimiento de la diversidad de los terópodos y las ventajas de la cladística aplicada en el estudio de las relaciones entre las especies, ninguno de esos terópodos está considerado como un alosáurido, aunque varios de estos como Acrocanthosaurus y Yangchuanosaurus, son miembros de familias cercanas.[23]

Allosaurus es el género tipo de la familia Allosauridae, la cual fue también nombrada por Marsh en 1878. En la taxonomía filogenética, el grupo Allosauridae es usualmente definido como "todos los carnosaurios más cercanamente relacionados con Allosaurus que con el Sinraptor o Carcharodontosaurus". Cuatro géneros han sido formalmente descritos los cuales son el Allosaurus y sus parientes más cercanos, otros dos géneros, "Madsenius"[72] y "Wyomingraptor",[73] no han sido nombrados formalmente y tampoco se conocen por muchos restos.
Allosauridae fue una de las cuatro familias de Carnosauria, las otras tres son Carcharodontosauridae, Metriacanthosauridae[23] y Neovenatoridae.[74] Allosauridae fue en un tiempo propuesto como ancestro de la familia Tyrannosauridae, lo cual los haría parafiléticos, con un ejemplo reciente en Gregory S. Paul's Predatory Dinosaurs of the World,[75] pero esto ha sido rechazado ya que los tiranosauroideos han sido identificados como miembros de Coelurosauria, un grupo totalmente separado de terópodos.[76] Allosauridae fue la más pequeña familia dentro de Carnosauria, solo con Saurophaganax y un alosauroide francés sin describir aceptados como géneros válidos aparte de Allosaurus en las revisiones más recientes.[23] El otro género, Epanterias, es potencialmente válido, el cual junto con Saurophaganax es considerado a veces como ejemplares gigantes de Allosaurus.[17] Las últimas revisiones aceptan mantener a Saurophaganax como válido e incluyen a Epanterias en Allosaurus.[77][23]
A la fecha abril de 2018, Carnosaurina no se considera válida, y Allosaurus forma parte de la superfamilia allosauroidea, que contiene a su vez a Metriacanthosauridae y Carcharodontosauridae.[78]
Filogenia
Cladograma según Benson et al. de 2010.[79]
| Allosauroidea |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Especies de Allosaurus
Todavía no está claro el número de especies conocidas de Allosaurus. Al menos ocho especies han sido consideradas potenciales válidas desde 1988 (A. amplexus,[17] A. atrox,[17] A. europaeus,[80] la especie tipo A. fragilis,[23] la todavía no descrita formalmente "A. jimmadseni",[14] A. lucasi[81] A. maximus,[49] y A. tendagurensis[23]), aunque generalmente solamente una fracción de estas sea considera válida en un momento dado. Además, hay por lo menos diez especies dudosas o sin describir que se han asignado a Allosaurus, junto con la especie que pertenece a los géneros hoy inválidos de Allosaurus. En la revisión más reciente de tetanuros basales, solamente A. fragilis (' incluyendo; ' A. amplexus' ' y ' ' A. atrox' ' como sinónimos), A "Jimmadseni".; (como especie sin nombre), y A. tendagurensis fueron aceptados como especies potencialmente válidas, con A. europaeus no todavía propuesta y A. maximus asignado a Saurophaganax.[23] al igual que A.jimmanedseni, al que consideró "nomen nudum", y por tanto, sinónima de fragilis.
Allosaurus se usó como sinónimo probable de los géneros Antrodemus, Creosaurus, Epanterias y Labrosaurus. La mayor parte de las especies que se ven como sinónimos de A. fragilis, o han sido apartados del género, debido a que están basadas en materiales y escaso. Una excepción es Labrosaurus ferox, nombrado en 1884 por Marsh para una mandíbula inferior parcial de formada extraña, con un boquete prominente en la fila del diente en el extremo, y una sección posterior ampliada y girada grandemente abajo.[82] Investigaciones posteriores sugirieron que es una patología del hueso, una lesión que sufrió el animal en vida, y esa forma inusual de la parte posterior del hueso era debido a la reconstrucción del yeso.[83] Hoy en día se considera un ejemplar de A. fragilis.[23]

Se han identificado varias especies de Allosaurus a lo largo de la historia, sin embargo muchas de ellas han resultado ser sinónimos de la especie tipo Allosaurus fragilis o trasladadas a otros géneros, con la posible excepción del espécimen de Epanterias amplexusO el género cercanamente relacionado Saurophaganax (OMNH 1708) , que es a veces incluido en el género Allosaurus como A. maximus, pero recientes estudios apoyan la idea de que pertenece a un género separado.[84] Otras especies que se creyeron pertenecer al género Allosaurus también fueron desplazadas a otros géneros.
La especie más extendida fue A. fragilis que habitó en EE. UU. y en Portugal había una especie muy relacionada, A. europeus, ya que en aquel entonces estaban conectados.[85] Sin embargo, esta especie fue sugerida mera y tentatv por las distintas localizaciones geográficas, y los autores reconocen que ML415 es indistinguible de cranéos de Allosaurus fragilis de Norteamérica. Carrano 2012 y Malafaia et al 2013 y 2014 consideró A.europaeous sinónimo de A.fragilis, basado en que es virtualmente indistinguible de esta especie usando tanto caracteres craniales como postcraniales.[86][2]
A. amplexus, A. atrox, A. fragilis, A. lucasi, "A. jimmadseni" y A. maximus son todas conocidas del intervalo Kimmeridgiense–Titoniense en el Jurásico superior de la Formación Morrison de los Estados Unidos, apareciendo a través de los estados de Colorado, Montana, Nuevo México, Oklahoma, Dakota del Sur, Utah y Wyoming. A. fragilis es sin duda la más común, con alrededor de sesenta especímenes conocidos.[23] La discusión ocurre desde los años 80 con respecto a la posibilidad que haya dos especies comunes en la Formación Morrison de Allosaurus, con la segunda conocida como ' ' A. atrox' ';[17][87] Un trabajo reciente ha seguido una interpretación de "un especie",[23] con las diferencias vistas en el material de la Formación Morrison atribuido a la variación individual.[88][89] En 2014 una nueva especie, A. lucasi fue descrita a partir de YPM VP 57589, estos fósiles fueron recogidos por Joseph T. Gregory y David Techter en 1953 en el yacimiento Cañón de McElmo, en el Condado de Montezuma, Colorado, Estados Unidos y se conservan en el Museo Peabody de Historia Natural de la Universidad de Yale.[81]
Se ha hallado a A. europaeus en sedimentos del Kimeridgiano del Miembro Porto Novo de la Formación Lourinhã,[80] pero podría ser un A. fragilis.[90] A. tendagurensis fue encontrado en rocas del Kimeridgiano de Tendaguru, en Mtwara, Tanzania.[91] Aunque la revisión más reciente lo aceptara tentativo como especie válida de Allosaurus, puede ser un tetanuro más básico,[92] o un simple terópodo dudóso.[93] Aunque sea oscuro, fue un terópodo grande, posiblemente alrededor de 10 metros de largo y 2,5 toneladas de peso,[10] del cual solo se ha encontrado una tibia parcial.[94] También se incluye al "A. robustus" de Australia,[95][96] el cual actualmente se concluyó que era un carnosaurio basal por su astrágalo (hueso del tobillo) muy similar al del fukuirráptor. Este pudo haber pertenecido a una especie similar a Australovenator si es que no era el mismo,[97] o también podría representar a un abelisaurio.[98] Una vértebra caudal fragmentaria de otra posible especie ha sido hallada en la provincia de Shanxi, China,[99] y seis dientes que datan del Jurásico medio y superior han sido recobrados de la Formación de Djaskoian, Rusia.,[100][101] y en Suiza,[93]
Especies válidas de Allosaurus
- A. fragilis — oeste de Estados Unidos
Posibles especies de Allosaurus
- A. europaeus - Portugal
- A. lucasi - oeste de Estados Unidos[81]
- A. tendagurensis - Tanzania
- A. atrox- Estados Unidos
- A. sp. indet.? — noreste central de China
- A. sp. indet.? — Rusia (noreste de Siberia)
- A. trihedrodon? — oeste de Estados Unidos
- A. maximus — trasladado a Saurophaganax maximus
- A. amplexus — trasladado dudosamente a E. amplexus, últimos estudios aportan a la idea de que es el sinónimo de A. fragilis.
Especies inválidas de Allosaurus
- A. lucaris — sinónimo de A. fragilis
- A. jimmadseni— oeste de Estados Unidos[102]
- A. ferox — sinónimo de A. fragilis
- "A. whitei" — sinónimo de A. fragilis
- "A. carnegeii" — sinónimo de A. fragilis
- "A. meriani" — sinónimo de Ceratosaurus meriani
- "A. robustus" — ahora considerado sinónimo de Australovenator
- "A. medius" — ahora considerado neoterópodo basal
- A. sibiricus — trasladado a Chilantaisaurus sibiricus[103]
- A. stechowi — trasladado a C. stechowi[104]
- A. valens — trasladado a Antrodemus valens
Paleobiología
Postura
Al igual que otros grandes carnívoros, el allosaurio era representado con una postura incorrecta, apoyando la cola al suelo permaneciendo como "trípode" al caminar, parecido a un canguro. Pinturas como las de Charles R. Knight y películas como El mundo perdido de Arthur Conan Doyle lo recrearon de esa manera. Aunque ya por 1970, científicos comprobaron que esta posición era errónea y si el animal vivo la hubiera adoptado resultaría en dislocación o debilitamiento de varias articulaciones, incluyendo la cadera y la unión entre la cabeza y la columna espinal.[105]
Alimentación

Los paleontólogos aceptan que Allosaurus era un carnívoro que depredaba sobre una amplia gama de dinosaurios herbívoros con los más variados tamaños. Sus presas podían ser pequeñas, como el driosaurio, de tamaño medio como el camptosaurio o auténticos pesos pesados como el estegosaurio y varios saurópodos. En estos últimos casos, un alosaurio que actuase en solitario se encontraba en desventaja, por lo que es probable que solo atacara a los individuos más débiles de estas especies, crías, ancianos o enfermos. No obstante, no se puede descartar que actuara en pequeños grupos, en cuyo caso solo los saurópodos más grandes y fuertes, como los diplodoco, apatosaurio y braquiosaurio estarían a salvo de sus ataques. Se ha encontrado de hecho, en los huesos de la cola de un espécimen de apatosaurio, marcas de dentellada de alosaurio, aunque podría ser resultado de un acto de carroña.[106][107] Existe dramática evidencia de que los alosaurios atacaban a los estegosaurios, incluyendo una vértebra de la cola de un alosaurio que se encuentra parcialmente curada de un lesión que concuerda con las espinas de la cola del un estegosaurio, y una placa de cuello de un estegosaurio que tiene una mordida en forma de U que concuerda con el hocico de un alosaurio.[108]
Al igual que cualquier otro carnívoro, los alosaurios también consumían carroña cuando se les presentaba la oportunidad, a veces con nefastas consecuencias: el famoso yacimiento de Cleveland Lloyd Quarry, en Utah (Estados Unidos), alberga los restos de decenas de alosaurios, en muchos casos jóvenes, que probablemente se vieron atraídos hasta una ciénaga por los cadáveres de dinosaurios herbívoros atrapados previamente allí, y luego quedaron atrapados uno tras otro. Usaban el olfato como principal medio para detectar su alimento, a juzgar por el gran desarrollo de los lóbulos olfativos en comparación con el resto del cerebro.
En cuanto al método de caza, es muy probable que prefiriesen la emboscada.[109] Aunque podían alcanzar grandes velocidades, no podían mantener éstas durante una larga carrera, por lo que preferían esperar agazapados en el bosque a que la presa llegara a su alcance. Con sus tres garras prensiles en cada mano, se servían para aferrarse a su presa, mientras que utilizaban sus potentes mandíbulas, capaces de ejercer más presión que las de un cocodrilo, para matarla por asfixia.
El alosaurio es el carnívoro de gran tamaño más abundante en los yacimientos donde aparece registrado, por lo que es posible que fuera el depredador dominante de su área de distribución. Otros grandes terópodos que convivieron con el alosaurio son el ceratosaurio, de menor tamaño y dotado también de protuberancias craneales similares a cuernos, y el gigantesco aunque escaso torvosaurio.
Volviendo a la predación de los saurópodos, Gregory Paul noto en 1988, que Allosaurus no fuese un predador de individuos completamente crecidos, a menos que cazara en manadas, pues tenía un cráneo de tamaño modesto y dientes relativamente pequeños, y fue sobrepasado en tamaño por los saurópodos contemporáneos.[17] Otra posibilidad es que prefirió cazar a jóvenes en vez de adultos completamente crecidos.[87][110] Investigaciones en los años 90 y el 2000s dieron otras soluciones posibles a esta pregunta. Robert Bakker, comparando Allosaurus con los Tigre diente de sable del cenozoico, encontró adaptaciones similares, tales como una reducción de los músculos de la quijada y aumento en músculos del cuello, y la capacidad de gran apertura de las fauces. Aunque Allosaurus no tenía dientes del sable, Bakker sugirió otro modo de ataque que habría utilizado tales adaptaciones del cuello y de la quijada. Los dientes cortos en efecto se convirtieron en pequeñas dientes de sierra, produciendo grandes mordidas muy sangrantes, en la que la mandíbula superior se clava en la víctima y con los fuertes músculos del cuello desgarran la carne de la víctima. Este tipo de estrategia permitiría ataques contra una presa mucho más grande, con la meta de debilitar a la víctima.[53]

Otros aspectos de la alimentación incluyen los ojos, los brazos, y las piernas. La forma del cráneo de Allosaurus le brinda un campo de visión binocular de 20°, levemente menos que el cocodrilo moderno. Como en los cocodriloideos, pudo haber sido lo bastante bueno como para evaluar adecuadamente la distancia y tiempo de ataque a la presa.[111] Los brazos, comparados con los de otros terópodos, estaban adaptados para agarrar la presa en una distancia corta,[31] y la articulación de las garras sugiere que habrían podido ser utilizados para enganchar cosas. [30] Finalmente, la máxima velocidad de Allosaurus se ha estimado en 8 a 15 metros por segundo, alrededor de 30 a 55 kilómetros por hora.[112]
Conclusiones similares fueron encontradas por otro estudio utilizando el análisis de elementos finitos en un cráneo Allosaurus. De acuerdo con su análisis biomecánico, el cráneo era muy fuerte, pero tenía una fuerza de mordida relativamente pequeña. Mediante el uso de los músculos de la mandíbula solamente, podría producir una fuerza de mordida de 805 a 2148 N, inferior a los valores de caimanes, 13000 N, leones, 4167 N y leopardos, 2268 N, pero el cráneo podría soportar casi 55.500 N de fuerza vertical contra la hilera de dientes. Los autores sugirieron que Allosaurus utilizó su cráneo como un hacha de guerra contra su presa, atacando con la boca abierta, cortando carne con sus dientes, no entrando muy profundo para no astillar el hueso, a diferencia de Tyrannosaurus, que se cree que han sido capaces de romper los huesos. También sugirieron que la arquitectura del cráneo podría haber permitido el uso de diferentes estrategias en contra de presas diferentes, el cráneo era lo suficientemente ligero como para permitir los ataques a ornitópodos pequeños y más ágiles, y lo suficientemente fuerte como para soportar el alto impacto en emboscadas contra presas más grandes como estegosauridos y saurópodos.[52] Sus interpretaciones fueron criticadas por otros investigadores, quienes no encontraron análogos modernos a un ataque de hacha y consideraron más probable que el cráneo era fuerte para compensar su construcción abierta al absorber las tensiones de luchar con la presa.[113] Los autores del primer estudio hicieron notar que Allosaurus en sí no tiene equivalente moderno y que la hilera de dientes que se adapta bien a este tipo de ataque, y que las articulaciones del cráneo citadas por sus detractores como un problema realmente ayudan a proteger la boca y reducir la tensión.[114] Otro posibilidad para el manejo de grandes presas es que los terópodos como el Allosaurus eran "herbívoros de carne" que podría tomar bocados de carne de saurópodos vivos que eran suficientes para sostener el depredador, por lo que no habría sido necesario hacer el esfuerzo para matar a la presa por completo. Esta estrategia potencialmente también habría permitido a la presa recuperarse y ser alimentado de manera similar más adelante.[23] Se observa como sugerencia adicional que los ornitópodos eran las más comunes dinosaurios presas disponibles, y que los alosaurios los dominaban mediante el uso de un ataque similar a la de los grandes felinos modernos: agarrar la presa con sus patas delanteras, y luego hacer las mordidas múltiples en la garganta para aplastar la tráquea,[110] Esto es compatible con otras pruebas de que las patas delanteras son fuertes y capaces de contener presa.[31]
Reproducción y crecimiento
El crecimiento se conoce realmente bien en esta especie gracias al hallazgo de individuos de ambos sexos y todas las edades. El yacimiento portugués de Lourinhã, donde se han encontrado fósiles de crías pequeñas, nidos y unos 100 huevos (varios con restos de embriones fosilizados en su interior) han permitido averiguar bastante sobre cómo nacían y se reproducían estos animales. Cada nido albergaba decenas de huevos que se incubaban solos, enterrados en el suelo como los de los cocodrilos. Los recién nacidos, similares a adultos en miniatura, ya podían caminar y tenían pequeños dientes aptos para comer insectos y pequeños invertebrados, pero no se podían valer realmente por sí mismos. Es probable que en sus primeras horas tras la salida del huevo fuesen tutelados por su madre de forma similar a como ocurre actualmente entre los cocodrilos y aligátores. Posteriormente, crecían rápidamente como las aves y al contrario que los cocodrilos, hasta alcanzar el gran tamaño de los adultos entre los 6 y 8 años.

La abundancia de fósiles de Allosaurus, de individuos de casi todas las edades, permite que los científicos estudien cómo el animal creció y cuánto tiempo pudo haber vivido. Los restos pueden ser rastreados a las más tempranas etapas de la vida, incluyendo huevos, cáscaras de huevos machacados de Colorado que se han encontrado pertenecientes a Allosaurus.[93] Basado en técnicas de análisis histológico de los huesos del miembro, el límite de edad superior para Allosaurus se estima en 22 a 28 años, que es comparable al de otros terópodos grandes como Tyrannosaurus. Del mismo análisis, su crecimiento máximo aparece haber sucedido a la edad de 15 años, con un índice de crecimiento estimado alrededor de 150 kilogramos por año.[50]
El tejido medular del hueso, que también encontró en dinosaurios tan diversos como Tyrannosaurus y Tenontosaurus, se ha encontrado en por lo menos un espécimen Allosaurus, una hueso de espinilla de la mina de Cleveland-Lloyd. Hoy, este tejido óseo se forma solamente en hembras de aves que están poniendo los huevos y se utilizan para proveer calcio a las cáscaras. Su presencia en el individuo Allosaurus establece el sexo y demuestra que ella habría alcanzado edad reproductiva. Contando líneas del crecimiento, fue demostrado que tenía 10 años a su muerte, estableciendo que la madurez sexual en Allosaurus fue lograda mucho antes de finalizar el crecimiento y alcanzar el tamaño máximos.[115]
El descubrimiento de un espécimen juvenil con una pata trasera casi completa demuestra que estas eran relativamente más largas en jóvenes, y los segmentos más bajos de la pierna, espinilla y pie. eran relativamente más largos que el muslo. Estas diferencias sugieren que un Allosaurus joven era más rápido y tenía diferentes estrategias de la caza que los adultos, quizás persiguiendo la pequeña presa como jóvenes, después se convertían en cazadores de emboscada a la edad adulta.[51] El fémur se volvió más denso y más ancho durante el crecimiento, y de la sección transversal menos circular, con cambios en los puntos e inserción muscular, los músculos se acortaron, y el crecimiento de la pierna se hizo más lento. Estos cambios implican que las piernas juveniles tienen tensiones menos fiables comparadas con los adultos, que se habrían movido con una progresión delantera más regular.[116]
Comportamiento social
Se ha especulado desde 1970 que Allosaurus se alimentaban de saurópodos y otros dinosaurios grandes que los cazaba en grupos.[117] Se lo ha mostrado en la literatura semitécnica y popular como animal que cazó a saurópodos y otros dinosaurios grandes en grupos.[70][20][87] Robert T. Bakker ha ampliado el comportamiento social al cuidado parental, y ha interpretado los dientes del alosaurio rompían y masticaban los huesos de los animales grandes como evidencia que los alosaurios adultos traían el alimento a la guarida para que sus crías comieran hasta que estuviesen crecidos, y evitó que otros carnívoros limpiaran en el alimento.[55] Sin embargo, hay realmente poca evidencia del comportamiento gregario en terópodos,[23] y las interacciones sociales con los miembros de la misma especie habrían incluido encuentros antagónicos, como se muestra por lesiones de las gastralia[25] y heridas de mordedura en los cráneos, la quijada inferior patológica nombrada Labrosaurus ferox es un posible ejemplo. El morder la cabeza pudo haber sido una manera de establecer la dominación en una manada de cazadores o durante conflictos territoriales.[118]

Aunque Allosaurus puede haber cazado en manadas,[119] la investigación reciente sugiere que Allosaurus y otros terópodos eran como otros diapsidos y tendían a comportamiento agresivo en vez de interacciones cooperativas con otros miembros de sus propias especies. Un estudio observó la caza cooperativa de la presa mucho más grande que un depredador individual, como se deduce comúnmente para los dinosaurios terópodos, es raro entre vertebrados y generalmente los modernos diapsidos (incluyendo lagartos, cocodrilos, y pájaros) cooperan muy raramente para cazar de esta manera. Muchos depredadores modernos diapsidos son territoriales y matarán y se comerán a los intrusos de la misma especie, y también harán lo mismo con individuos más pequeños que intenten comer antes de ellos cuando están juntos en los sitios de alimentación. Esto sugiere que, por ejemplo, que en sitios como Cleveland-Lloyd se ve que Allosaurus acudieron juntos a alimentarse de otros alosaurios inmovilizados o muertos, y fueron muertos en el proceso, así acumulándose. Esto podría explicar la elevada presencia de alosaurios jóvenes y subadultos presentes, como matan a los jóvenes y en los sitios de alimentación de grupos moderno como los de cocodrilos y dragones de komodo, la tasa de no adultos muertos es mayor. La misma interpretación se aplica al sitio de Bakker.[120] Hay una cierta evidencia del canibalismo entre alosaurios, incluyendo marcas de dientes de alosaurios encontrados entre los fragmentos de la costilla y posibles marcas en un omóplato, [121] y huesos canibalizados recuperados en el sitio de Bakker.[122]
Órganos sensoriales
El cerebro de Allosaurus, según lo interpretado de espiral Exploración del CT del molde endocraneal, era más similar con un cocodrilo que con los de otros arcosaurios vivos, las aves. La estructura del aparato vestibular indica que el cráneo se desarrolló casi horizontal, en comparación con inclinado fuertemente hacia arriba o hacia abajo. La estructura del oído interno era como la de un cocodriloideo, y por lo que Allosaurus habría probablemente podido oír mejor frecuencias más bajas y habría tenido problemas con los sonidos sutiles. El bulbo olfatorio era grande y parece haber estado bien adaptado para detectar olores, aunque el área para la evaluación de olores fuera relativamente pequeña.[54]
Paleopatología
En 2001, Bruce Rothschild et al, publicaron un estudio que examina la evidencia de fracturas por estrés y avulsiones de tendones en dinosaurios terópodos y las consecuencias en su comportamiento. Dado que las fracturas de estrés son causadas por un trauma repetidos en lugar de un acontecimientos singular, son más probable que sean causadas por el comportamiento del animal que otros tipos de lesiones. Las fracturas por estrés y avulsiones de tendones que se producen en los miembros anteriores tienen un significado especial en el comportamiento mientras que las lesiones en los pies pueden ser causadas por correr o migrar, la resistencia de las presas son la fuente más probable de las lesiones de la mano. Allosaurus fue uno de los dos únicos terópodos examinados en el estudio en exhibir una avulsión de tendón, y en ambos casos la avulsión se produjo en la extremidad anterior. Cuando los investigadores analizaron las fracturas de estrés, se encontraron con que Allosaurus tenía un número significativamente mayor de fracturas por estrés que Albertosaurus, Ornithomimus o Archaeornithomimus. De los 47 huesos de la mano que se estudiaron, 3 fueron encontrados con fracturas por estrés. De los pies, los huesos que se estudiaron fueron 281 y en 17 se encontró fracturas por estrés. Las fracturas por estrés en los huesos del pie "fueron distribuidas en las falanges proximales" y se produjeron en los tres dedos del pie que soportan peso siendo "estadísticamente indistinguibles" entre ellos. Dado que el extremo inferior del tercer metatarsiano habría hecho contacto con el suelo primero mientras que el animal corría, se llevaba la mayor parte de la tensión. Si las fracturas por estrés en los alosaurios fueron causadas por la acumulación de daño al caminar o correr este hueso debería tener más fracturas por estrés que los otros. La falta de tal sesgo en los fósiles estudiados de Allosaurus indica un origen para las fracturas de tensión tienen una fuente distinta que al correr. Los autores concluyen que estas fracturas se produjeron durante la interacción con la presa, como un alosaurio tratando de sostener la presa que lucha con sus pies. La abundancia de las fracturas por estrés y lesiones por avulsión en Allosaurus proporcionan evidencia de "muy activa predación" en vez de carroñero.[123]

La escápula izquierda y el peroné de un espécimen de A. fragilis catalogado como USNM 4734 son patológicos, ambas probablemente debido a las fracturas curadas. El ejemplar USNM 8367 conserva varios gastralias patológicas que conservan evidencias de fracturas curadas cerca de su centro. Algunas de las fracturas estaban mal sanadas y se había formado una "pseudoartrosis". Un ejemplar con una costilla fracturada fue recuperado de la cantera de Cleveland-Lloyd. Otro espécimen tenía costillas fracturadas y vértebras fusionadas cerca del final de la cola. Un aparente macho subadulto de A. fragilis se informó a tener patologías extensas, con un total de catorce lesiones separadas. El ejemplar MOR 693 tenían patologías en cinco costillas, en la sexta vértebra del cuello , la tercera octava y decimotercera vértebras de la espalda, la segunda vértebra de su cola y su cheuron, una gastralia, escápula derecha, falange manual del dedo 1, ilion izquierdo, metatarsianos III y V, la primera falange del tercer dedo del pie y la tercera falange del segundo. El ilion tenía "un gran agujero causado por un golpe desde arriba". El extremo próximo de la primera falange del tercer dedo fue afectado por hueso neoformado.[124] Otras patologías reportados en Allosaurus incluyen, fracturas en rama verde en dos costillas. Fracturas consolidadas en el húmero y el radio. La distorsión de las superficies articulares del pie, posiblemente debido a problemas de artrosis o de desarrollo. Las distorsiones de las superficies articulares de las vértebras cola posiblemente debido a osteoartritis o problemas de desarrollo. Una gran anquilosis neoplasica de caudales, posiblemente debido a un traumatismo físico, así como la fusión de cheurones al centro. Osificación de centros vertebrales cerca del final de la cola. Amputación de un hueso cheuron y el pie, tanto posiblemente como consecuencia de mordeduras. Exostosis extensiva en la primera falange del tercer dedo. Lesiones similares a las provocadas por osteomielitis en dos escápulas. Espolones óseos en un premaxilar, ungueal y dos metacarpianos. Exostosis en una falange pedal posiblemente atribuible a una enfermedad infecciosa. Un metacarpiano con una fractura por compresión.[124]
Paleoecología
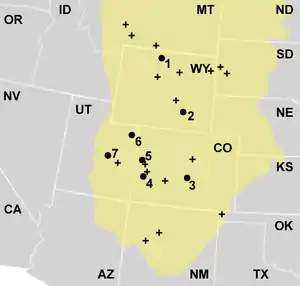
El alosaurio es el terópodo más común en la zona extensa de rocas fósiles en el Suroeste Americano conocida como la Formación de Morrison. Sus hallazgos son el 70 a 75% de los especímenes hallados en dicha formación,[110] y por mucho es la cúspide de la pirámide alimenticia de dicha formación.[125] Otros restos han sido recolectados en Montana, Wyoming, Dakota del Sur, Colorado, Oklahoma, Nuevo México y Utah, en los Estados Unidos. También ha habido hallazgos en Portugal. La Formación Morrison es interpretada como un ambiente semiárido con temporada húmeda con llanuras inundables.[126] La vegetación variaba de bosques de coníferas a los lados de los ríos, Helechos arborescentes y helechos, pasando a sabanas de helechos con algunos árboles similares a Araucarias del género Brachyphyllum.[127] La Formación Morrison es un área muy rica en fósiles, en ella se encuentran clorofitos, hongos, musgos, equisetos, helechos, cycadales, ginkgos, y varias familias de coníferas. Otros fósiles incluyen bivalvos, caracoles, Actinopterigios, ranas, salamandras, tortugas, esfenodontos, lagartos, cocodrilomorfos terrestres y acuáticos, varias especies de pterosaurios, numerosas especies de dinosaurios, y mamíferos primitivos docodontes, multituberculados, symmetrodontes, y triconodontes. Algunos de los dinosaurios terópodos fueron Ceratosaurus, Ornitholestes, y Torvosaurus, los saurópodos Apatosaurus, Brachiosaurus, Camarasaurus, y Diplodocus, y los ornitisquios Camptosaurus, Dryosaurus, y Stegosaurus son conocidos en Morrison.[102] Las formaciones de finales del Jurásico en Portugal donde Allosaurus está presente son similares a la de Morrison pero con mayor influencia marina. Muchos de los géneros de dinosaurios de la Formación de Morrison están presentes, como Allosaurus, Ceratosaurus, Torvosaurus, y Apatosaurus, o tienen una cercana contraparte como Brachiosaurus y Lusotitan, Camptosaurus y Draconyx.[128]
Allosaurus coexistió con otros grandes teropódos como Ceratosaurus y Torvosaurus, ambos en Portugal y en Estados Unidos,[128] Los tres parecieron tener distintos nichos ecológicos, basado en la localización y morfología de los fósiles. Ceratosaurios y torvosaurios pueden haber preferido ser activos alrededor de los canales de agua, y tenían cuerpos más finos y más bajos, que les habrían dado una ventaja en terrenos boscosos y maleza, mientras que los alosaurios eran más compactos, con piernas más largas, más rápidos y maniobrables, y parecen haber preferido las llanuras inundadas.[122] Ceratosaurus, más conocido que Torvosaurus, se diferenciaba perceptiblemente de Allosaurus en su anatomía funcional teniendo un cráneo más alto, más estrecho con dientes grandes y amplios.[44] Se sabe que pedúnculo púbico de un Allosaurus muestra marcas de dientes de probablemente un Ceratosaurus o un Torvosaurus. La localización de este hueso, en lo profundo del cuerpo indica que dicho Allosaurus debió sufrir la acción de carroñeros.[129]
En la cultura popular

Junto con Tyrannosaurus, Allosaurus es el terópodo grande más representado en la cultura popular. Es un dinosaurio común en los museos, en particular después de las excavaciones de la mina de dinosaurios de Cleveland Lloyd en 1976, como resultado de un esfuerzo conjunto, 38 museos de 8 países de 3 continentes tienen material de Cleveland-Lloyd o copias de estos.[4] Allosaurus es el dinosaurio oficial del estado de Utah.[130]
Allosaurus ha aparecido en los medios de comunicación desde principios del siglo XX. Es el principal predador en la novela de 1912 de Arthur Conan Doyle, El mundo perdido, y en la adaptación cinematográfica de 1925, el primer largometraje con dinosaurios y las versiones de 1960, 1992 y 1998 con una secuela del film de 1992 y miniserie por la cadena de televisión BBC llamada The Lost World (2001) y la serie de televisión basada en el libro con el nombre Sir Arthur Conan Doyle's The Lost World.[131] Más tarde aparece en el filme de 1956 La bestia de la Montaña Hollow,[132] y en el filme de 1969 El valle de Gwangi, una combinación del género de terror con el western. En El valle de Gwangi, Gwangi es creado como Allosaurus, aunque Ray Harryhausen basa su modelo para la criatura en la pintura Charles R. Knight' pintura de un Tyrannosaurus. Harryhausen confunde a veces a los dos, indicando en una entrevista de DVD «Ambos son comedores de la carne, ambos son tiranos… uno era apenas un poco más grande que el otro».[133]
En apariciones documentales, Allosaurus aparece en el segundo y el quinto episodio de la serie de televisión de la BBC Walking with Dinosaurs, y en el especial de Walking with Dinosaurs "La Balada del Gran Al" mostrando la crónica de la vida del espécimen de Allosaurus llamado "Big Al". También aparece en dos episodios de Jurassic Fight Club.[134] Allosaurus también hace su aparición en la serie de Discovery Channel, Dinosaur Revolution. Su representación en esta serie se basó en una muestra con una mandíbula rota inferior que fue descubierta por el paleontólogo Thomas Holtz.[135]
En la serie animada de los años 1980 Dinosaucers, un grupo de dinosaurios con mentalidad desarrollada a nivel humano, llega al planeta Tierra desde el ficticio universo de Reptilión. Su líder Allo, está basado en el Allosaurus. Él y su equipo llegan y pactan una alianza con un grupo de adolescentes terrícolas, con el objetivo de defender el planeta de la amenaza de los Tyrannus, un grupo rival que también arribó a la Tierra, pero con intenciones de conquista. Su líder Gengis Rex, estaba basado en el Tyrannosaurus Rex.
Referencias
- Holtz, Thomas (2012). Genus List for Holtz (2007) Dinosaurs:The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. Consultado el 26 de septiembre de 2015.
- E.Malafaia, P.Dantas, F.Ortega1 y F.Escaso ;Nuevos restos de Allosaurus fragilis (Theropoda: Carnosauria) del yacimiento de Andrés (Jurásico Superior; Centro-Oeste de Portugal)
- C.W. Gilmore (1920) OSTEOLOGY OF THE CARNIVOROUS DINOSAURIA IN THE UNITED STATES NATIONAL MUSEUM, WITH SPECIAL REFERENCE TO THE GENERA ANTRODEMUS (ALLOSAURUS) AND CERATOSAURUS. SMITHSONIAN INSTITUTION UNITED STATES NATIONAL MUSEUM Bulletin 110
- Madsen, James H., Jr. (1993) [1976]. Allosaurus fragilis: A Revised Osteology. Utah Geological Survey Bulletin 109 (2ª ed. edición). Salt Lake City: Utah Geological Survey.
- Gregory S.Paul: The priceton field guide to dinosaurs (2016) second edition ISBN 978-0-691-16766-4
- http://theropoddatabase.com/Carnosauria.htm#Allosaurusfragilis
- Karl T. Bates, Roger B. J. Benson, and Peter L. Falkingham (2012): A computational analysis of locomotor anatomy and body mass evolution in Allosauroidea. (Dinosauria: Theropoda) Paleobiology, 38(3), 2012, pp. 486–507
- «Copia archivada». Archivado desde el original el 1 de diciembre de 2020. Consultado el 31 de diciembre de 2017.
- N.C Campione, D.C Evans,C.M Brown, M.T Carrano.(2014). Body mass estimation in non-avian bipeds using a theoretical conversion to quadruped stylopodial proportions. Methods in Ecology and Evolution 2014, 5, 913–923.
- Mortimer, Mickey (21 de julio). «And the Largest Theropod Is....». The Dinosaur Mailing List. Archivado desde el original el 25 de marzo de 2010. Consultado el 8 de setiembre de 2007.
- Paul, G. S. Predatory Dinosaurs of the World (Simon & Schuster, New York, 1988).
- Benson RBJ, Campione NE, Carrano MT, Mannion PD, Sullivan C, Upchurch P, et al. (2014) Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage. PLoS Biol 12(5): e1001853. https://doi.org/10.1371/journal.pbio.1001853
- Bates, Karl T.; Falkingham, Peter L.; Breithaupt, Brent H.; Hodgetts, David; Sellers, William I.; Manning, Phillip L. (2009). «How big was 'Big Al'? Quantifying the effect of soft tissue and osteological unknowns on mass predictions for Allosaurus (Dinosauria:Theropoda)». Palaeontología Electrónica 12 (3). Archivado desde el original el 25 de diciembre de 2009. Consultado el 13 de diciembre de 2009.
- Chure, Daniel J. (2000). A new species of Allosaurus from the Morrison Formation of Dinosaur National Monument (Utah–Colorado) and a revision of the theropod family Allosauridae. Ph.D. dissertation. Columbia University.
- H.F.Osborn, C.C.Mook : Camarasaurus, Amhicoelias, and other sauropods of Cope
- C.W. Gilmore (1920) OSTEOLOGY OF THE CARNIVOROUS DINOSAURIA IN THE UNITED STATES NATIONAL MUSEUM, WITH SPECIAL REFERENCE TO THE GENERA ANTRODEMUS (ALLOSAURUS) AND CERATOSAURUS. SMITHSONIAN INSTITUTION UNITED STATES NATIONAL MUSEUM Bulletin 110
- Paul, Gregory S. (1988). «Genus Allosaurus». Predatory Dinosaurs of the World. Nueva York: Simon & Schuster. pp. 307-313. ISBN 0-671-61946-2.
- Foster, John. 2007. Jurassic West: the Dinosaurs of the Morrison Formation and Their World. Bloomington, Indiana:Indiana University Press. p. 117.
- Molnar, Ralph E. (1977). «Analogies in the evolution of combat and display structures in ornithopods and ungulates». Evolutionary Theory 3: 165-190.
- Norman, David B. (1985). «'Carnosaurs'». The Illustrated Encyclopedia of Dinosaurs: An Original and Compelling Insight into Life in the Dinosaur Kingdom. Nueva York: Crescent Books. pp. 62-67. ISBN 0-517-468905.
- Paul, Gregory S. (1988). Predatory Dinosaurs of the World. 91 and Figure 4–5 (93).
- Madsen, 1976; note that not everyone agrees on where the neck ends and the back begins, and some authors such as Gregory S. Paul interpret the count as ten neck and thirteen back vertebrae.
- Holtz, Thomas R., Jr.; Molnar, Ralph E.; Currie, Philip J. (2004). «Basal Tetanurae». En Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.), ed. The Dinosauria (2ª edición). Berkeley: University of California Press. pp. 71-110. ISBN 978-0-520-24209-8.
- Paul, Gregory S. (1988). Predatory Dinosaurs of the World. 277.
- Chure, Daniel J. (2000). «Observations on the morphology and pathology of the gastral basket of Allosaurus, based on a new specimen from Dinosaur National Monument». Oryctos 3: 39-37. ISSN 1290-4805.
- Chure, Daniel J.; and Madsen, James H. (1996). «On the presence of furculae in some non-maniraptoran theropods». Journal of Vertebrate Paleontology 16 (3): 573-577.
- Chure and Madsen, 1996. On the presence of furculae in some non-maniraptoran theropods. Journal of Vertebrate Paleontology. 16(3): 573-577.
- Chure and Madsen, 1996. The furcula in allosaurid theropods and its implication for determining bird origins. Society of Vertebrate Paleontology. P. 28A.
- Middleton, Kevin M.; y Gatesy, Stephen M. (2000). «Theropod forelimb design and evolution». Zoological Journal of the Linnean Society 128: 149-187. doi:10.1006/zjls.1998.0193. Archivado desde el original el 25 de octubre de 2007. Consultado el 25 de octubre de 2007.
- Gilmore, Charles W. (1920). «Osteology of the carnivorous dinosauria in the United States National Museum, with special reference to the genera Antrodemus (Allosaurus) and Ceratosaurus''». Bulletin of the United States National Museum 110: 1-159.
- Carpenter, Kenneth (2002). «Forelimb biomechanics of nonavian theropod dinosaurs in predation» (pdf). Senckebergiana lethaea 82 (1): 59-76. Archivado desde el original el 27 de septiembre de 2007.
- Paul, Gregory S. (1988). Predatory Dinosaurs of the World. 113; ver ilustraciones del Allosaurus en 310 y 311; Madsen (1976) interpretó esos huesos como posibles porciones superiores del metatarsal interior .
- Leidy, Joseph (1870). «Remarks on Poicilopleuron valens, Clidastes intermedius, Leiodon proriger, Baptemys wyomingensis, and Emys stevensonianus''». Proceedings of the Academy of Natural Sciences, Philadelphia 22: 3-4.
- Marsh OC. (1877). Notice of new dinosaurian reptiles from the Jurassic formation. American Journal of Science and Arts 14:514-516
- Marsh, Othniel Charles (1878). «Notice of new dinosaurian reptiles». American Journal of Science and Arts 15: 241-244.
- Marsh, Othniel Charles (1879). «Principal characters of American Jurassic dinosaurs. Part II». American Journal of Science, Series 3 17: 86-92.
- Cope, Edward Drinker (1878). «A new opisthocoelous dinosaur». American Naturalist 12 (6): 406. doi:10.1086/272127.
- Marsh, Othniel Charles (1877). «Notice of new dinosaurian reptiles from the Jurassic formation». American Journal of Science and Arts 14: 514-516.
- Creisler, Ben (7 de julio de 2003). «Dinosauria Translation and Pronunciation Guide A». Dinosauria On-Line. Archivado desde el original el 30 de septiembre de 2007. Consultado el 11 de septiembre de 2007.
- Norell, Mark A.; Gaffney, Eric S.; Dingus, Lowell (1995). Discovering Dinosaurs in the American Museum of Natural History. Nueva York: Knopf. pp. 112-113. ISBN 0-679-43386-4.
- Breithaupt, Brent H.; Chure, Daniel J.; Southwell, Elizabeth H. (1999). «AMNH 5753: The world's first free-standing theropod skeleton». Journal of Vertebrate Paleontology 19 (3, Suppl.): 33A.
- Williston, Samuel Wendell (1878). «American Jurassic dinosaurs». Transactions of the Kansas Academy of Science 6: 42-46.
- Williston, Samuel Wendell (1901). «The dinosaurian genus Creosaurus, Marsh». American Journal of Science, series 4 11 (11): 111-114.
- Henderson, Donald M. (1998). «Skull and tooth morphology as indicators of niche partitioning in sympatric Morrison Formation theropods» (pdf). Gaia 15: 219-266. ISSN 0871-5424. Archivado desde el original el 27 de noviembre de 2007.
- Liddell & Scott (1980). Greek-English Lexicon, Abridged Edition. Oxford University Press, Oxford, UK. ISBN 0-19-910207-4.
- Stokes, William J. (1945). «A new quarry for Jurassic dinosaurs». Science 101 (2614): 115-117. PMID 17799203. doi:10.1126/science.101.2614.115-a.
- Hunt, Adrian P; Lucas, Spencer G.; Krainer, Karl; Spielmann, Justin (2006). «The taphonomy of the Cleveland-Lloyd Dinosaur Quarry, Upper Jurassic Morrison Formation, Utah: a re-evaluation». En Foster, John R.; y Lucas, Spencer G. (eds.), ed. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, Nuevo México: New Mexico Museum of Natural History and Science. pp. 57-65.
- Loewen, Mark A.; Sampson, Scott D.; Carrano, Matthew T.; Chure, Daniel J. (2003). «Morphology, taxonomy, and stratigraphy of Allosaurus from the Upper Jurassic Morrison Formation». Journal of Vertebrate Paleontology 23 (3, Suppl.): 72A.
- Smith, David K. (1998). «A morphometric analysis of Allosaurus''». Journal of Vertebrate Paleontology 18 (1): 126-142.
- Bybee, Paul J.; Lee, A.H.; Lamm, E.-T. (2006). «Sizing the Jurassic theropod dinosaur Allosaurus: Assessing growth strategy and evolution of ontogenetic scaling of limbs». Journal of Morphology 267 (3): 347-359. doi:10.1002/jmor.10406.
- Foster, John R.; y Chure, Daniel J. (2006). «Hindlimb allometry in the Late Jurassic theropod dinosaur Allosaurus, with comments on its abundance and distribution». En Foster, John R.; y Lucas, Spencer G. (eds.), ed. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, Nuevo México: New Mexico Museum of Natural History and Science. pp. 119-122.
- Rayfield, Emily J.; Norman, David B.; Horner, Celeste C.; Horner, John R.; Smith, Paula May; Thomason, Jeffrey J.; Upchurch, Paul (2001). «Cranial design and function in a large theropod dinosaur». Nature 409: 1033-1037. doi:10.1038/35059070.
- Bakker, Robert T. (1998). «Brontosaur killers: Late Jurassic allosaurids as sabre-tooth cat analogues» (pdf). Gaia 15: 145-158. ISSN 0871-5424. Archivado desde el original el 31 de octubre de 2007.
- Rogers, Scott W. (1999). «Allosaurus, crocodiles, and birds: Evolutionary clues from spiral computed tomography of an endocast» (pdf). The Anatomical Record 257 (5): 163-173. doi:10.1002/(SICI)1097-0185(19991015)257:5<162::AID-AR5>3.0.CO;2-W. Archivado desde el original el 1 de abril de 2020. Consultado el 15 de octubre de 2008.
- Bakker, Robert T. (1997). «Raptor Family values: Allosaur parents brought giant carcasses into their lair to feed their young». En Wolberg, Donald L.; Sump, Edmund; and Rosenberg, Gary D. (eds.), ed. Dinofest International, Proceedings of a Symposium Held at Arizona State University. Filadelfia: Academy of Natural Sciences. pp. 51-63. ISBN 0-935868-94-1.
- Chure, Daniel J. (1995). «A reassessment of the gigantic theropod Saurophagus maximus from the Morrison Formation (Upper Jurassic) of Oklahoma, USA». En Ailing Sun y Yuangqing Wang (eds.), ed. Sixth Symposium on Mesozoic Terrestrial Ecosystems and Biota, Short Papers. Beijing: China Ocean Press. pp. 103-106. ISBN 7-5027-3898-3.
- Pérez-Moreno, B.P.; Chure, D.J.; Pires, C.; Silva, C.M.; Santos, V.; Dantas, P.; Póvoas, L.; Cachão, M.; Sanz, J.L.; Galopim De Carvalho, A.M. (1999). «On the presence of Allosaurus fragilis (Theropoda: Carnosauria) in the Upper Jurassic of Portugal: First evidence of an intercontinental dinosaur species» (pdf). Journal of the Geological Society 156 (3): 449-452. doi:10.1144/gsjgs.156.3.0449. Archivado desde el original el 25 de octubre de 2007.
- Breithaupt, Brent (1996). «The discovery of a nearly complete Allosaurus from the Jurassic Morrison Formation, eastern Bighorn Basin, Wyoming». En Brown, C.E.; Kirkwood, S.C.; y Miller, T.S. (eds)., ed. Forty-Seventh Annual Field Conference Guidebook. Casper, Wyoming: Wyoming Geological Association. pp. 309-313. OCLC 36004754.
- «Howe Dinosaur Quarry – Wyoming’s Jurassic Treasure». GeoScience Adventures. 24 de julio de 2007. Consultado el 27 de septiembre de 2007.
- Chure and Fiorillo, 1997. One Big Al to go and hold the mayo: Evidence of scavenging of a specimen of Allosaurus from the Morrison Formation (Late Jurassic) of Wyoming:: Journal of Vertebrate Paleontology, v. 17, Supplement to no. 3, p. 38A.
- Hanna, 2002. Multiple injury and infection in a sub-adult theropod dinosaur Allosaurus fragilis with comparisons to allosaur pathology in the Cleveland-Lloyd Dinosaur Quarry collection: Journal of Vertebrate Paleontology, v. 22, n. 1, p. 76-90.
- Breithaupt, Brent H. «The case of "Big Al" the Allosaurus: a study in paleodetective partnerships». Archivado desde el original el 17 de enero de 2010. Consultado el 3 de octubre de 2007.
- Hanna, Rebecca R. (2002). «Multiple injury and infection in a sub-adult theropod dinosaur (Allosaurus fragilis) with comparisons to allosaur pathology in the Cleveland-Lloyd dinosaur quarry collection». Journal of Vertebrate Paleontology 22 (1): 76-90. doi:10.1671/0272-4634(2002)022[0076:MIAIIA]2.0.CO;2.
- Foth, C.; Evers, S.; Pabst, B.; Mateus, O.; Flisch, A.; Patthey, M.; Rauhut, O. W. M. (2015). «New insights into the lifestyle of Allosaurus (Dinosauria: Theropoda) based on another specimen with multiple pathologies». PeerJ PrePrints 3: e824v1. doi:10.7717/peerj.940.
- von Huene, Friedrich (1926). «The carnivorous Saurischia in the Jura and Cretaceous formations, principally in Europe». Revista del Museo de La Plata 29: 35-167.
- Romer, Alfred S. (1956). Osteology of the Reptiles. Chicago: University of Chicago Press. pp. 772pp. ISBN 0-89464985-X.
- Romer, Alfred S. (1966). Vertebrate Paleontology (Tercera Edición edición). Chicago: University of Chicago Press. pp. 468 pp. ISBN 0-7167-1822-7.
- Steel, R. (1970). «Part 14. Saurischia. Handbuch der Paläoherpetologie/Encyclopedia of Paleoherpetology». Gustav Fischer Verlag, Stuttgart (en alemán): 1-87.
- Walker, Alick D. (1964). «Triassic reptiles from the Elgin area: Ornithosuchus and the origin of carnosaurs». Philosophical Transactions of the Royal Society of London, Series B, Biological Sciences 248 (744): 53-134. doi:10.1098/rstb.1964.0009.
- Lambert, David; and the Diagram Group (1983). «Allosaurids». A Field Guide to Dinosaurs. Nueva York: Avon Books. pp. 80-81. ISBN 0-380-83519-3.
- Lambert, David; and the Diagram Group (1990). «Allosaurids». The Dinosaur Data Book. Nueva York: Avon Books. p. 130. ISBN 0-380-75896-2.
- Lambert, 1990. The Dinosaur Data Book.
- "Bakker, 1997. Dr. Bob's Dinofacts. Tate Geological Times 5(2) Marzo/Abril 1997: p. 3.
- Benson, R. B. J.; Carrano, M. T.; Brusatte, S. L. (2009). "A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic". Naturwissenschaften 97 (1): 71–78. Bibcode 2010NW.....97...71B. doi:10.1007/s00114-009-0614-x. PMID 19826771. edit
- Paul, Gregory S. (1988). "The allosaur-tyrannosaur group", Predatory Dinosaurs of the World. 301–347.
- Holtz, Thomas R., Jr. (1994). «The phylogenetic position of the Tyrannosauridae: Implications for theropod systematics». Journal of Paleontology 68 (5): 1100-1117.
- Foster, John R. (2003). Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A. New Mexico Museum of Natural History and Science Bulletin 23 (en inglés). Albuquerque: New Mexico Museum of Natural History and Science. p. 37.
- Matthew T. Carrano, Roger B. J. Benson & Scott D. Sampson (2012): The phylogeny of Tetanurae (Dinosauria: Theropoda), Journal of Systematic Palaeontology, 10:2, 211-300
- Benson, R.B.J.; Carrano, M.T.; Brusatte, S.L. (2010). «A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic». Naturwissenschaften 97 (1): 71-78. Bibcode:2010NW.....97...71B. PMID 19826771. doi:10.1007/s00114-009-0614-x.
- Mateus, Octávio; Walen, Aart; Antunes, Miguel Telles (2006). «The large theropod fauna of the Lourinha Formation (Portugal) and its similarity to that of the Morrison Formation, with a description of a new species of Allosaurus». En Foster, John R.; y Lucas, Spencer G. (eds.), ed. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, Nuevo México: New Mexico Museum of Natural History and Science. pp. 123-129.
- Dalman, S.G. 2014. Osteology of a large allosauroid theropod from the Upper Jurassic (Tithonian) Morrison Formation of Colorado, USA. Volumina Jurassica, 12(2): 159-180.
- Marsh, Othniel Charles (1884). «Principal characters of American Jurassic dinosaurs. Part VIII». American Journal of Science, Series 3 27: 329-340.
- Madsen, James H.; and Welles, Samuel P. Ceratosaurus (Dinosauria, Theropoda), a Revised Osteology. Miscellaneous Publication, 00-2. Utah Geological Survey. ISBN 1-55791-380-3.
- Chure D. J. (2000). "A new species of Allosaurus from the Morrison Formation of Dinosaur National Monument (Utah-Colorado) and a revision of the theropod family Allosauridae." Ph.D. dissertation, Columbia University, 1-964.
- Mateus, O., Walen, A., and Antunes, M.T. (2006). "The large theropod fauna of the Lourinha Formation (Portugal) and its similarity to that of the Morrison Formation, with a description of a new species of Allosaurus." New Mexico Museum of Natural History and Science Bulletin, 36.
- Matthew T. Carrano, Roger B. J. Benson & Scott D. Sampson (2012): The phylogeny of Tetanurae (Dinosauria: Theropoda), Journal of Systematic Palaeontology, 10:2, 211-300
- Lessem, Don; and Glut, Donald F. (1993). «Allosaurus». The Dinosaur Society's Dinosaur Encyclopedia. Random House. pp. 19-20. ISBN 0-679-41770-2. OCLC 30361459.
- Smith, David K. (1996). «A discriminant analysis of Allosaurus population using quarries as the operational units». Museum of Northern Arizona Bulletin 60: 69-72.
- Smith, David K. (1999). «Patterns of size-related variation within Allosaurus''». Journal of Vertebrate Paleontology 19 (2): 402-403.
- Malafaia, Elisabete; Dantas, Pedro; Ortega, Francisco; Escaso, Fernando (2007). «Nuevos restos de Allosaurus fragilis (Theropoda: Carnosauria) del yacimiento de Andrés (Jurásico Superior; centro-oeste de Portugal)» (pdf). Cantera Paleontológica: 255-271.
- Janensch, Werner (1925). «Die Coelurosaurier und Theropoden der Tendaguru-Schichten Deutsch-Ostafrikas». Palaeontographica, Supplement 7 (en alemán) 1: 1-99.
- Rauhut, Oliver W.M. (2005). «Post-cranial remains of 'coelurosaurs' (Dinosauria, Theropoda) from the Late Jurassic of Tanzania». Geological Magazine 142 (1): 97-107. doi:10.1017/S0016756804000330.
- Glut, Donald F. (1997). «Allosaurus». Dinosaurs: The Encyclopedia (en inglés). Jefferson, North Carolina: McFarland & Co. pp. 105–117. ISBN 978-0-89950-917-4.
- Janensch, 1925. Die Coelurosaurier und Theropoden der Tendaguru-Schichten Deutsch-Ostafrikas. Palaeontographica (Suppl. 7)1:1-99.
- Molnar, Flannery and Rich, 1981. An allosaurid theropod dinosaur from the early Cretaceous of Victoria, Australia. Alcheringa 5 p. 141-146.
- Chure, 1998. A reassessment of the Australian Allosaurus and its implications for the Australian refugium concept. Journal of Vertebrate Paleontology 18(suppl. 3): 34A.
- Hocknull, Scott A.; White, Matt A.; Tischler, Travis R.; Cook, Alex G.; Calleja, Naomi D.; Sloan, Trish; Elliott, David A. (2009). «New Mid-Cretaceous (Latest Albian) Dinosaurs from Winton, Queensland, Australia». En Sereno, Paul, ed. PLoS ONE 4 (7): e6190. PMC 2703565. PMID 19584929. doi:10.1371/journal.pone.0006190.
- Agnolin, F. L.; Ezcurra, M. D.; Pais, D. F.; Salisbury, S. W. (2010). «A reappraisal of the Cretaceous non-avian dinosaur faunas from Australia and New Zealand: Evidence for their Gondwanan affinities». Journal of Systematic Palaeontology 8 (2): 257-300. doi:10.1080/14772011003594870.
- Lü and Hu, 1998. Dinosaur remains from Datong Suburb, Shanxi Province. Vertebrata PalAsiatica 36(3):252-256.
- Riabinin, Anatoly Nikolaenvich (1914). «Zamtka o dinozavry ise Zabaykalya». Trudy Geologichyeskago Muszeyah Imeni Petra Velikago Imperatorskoy Academiy Nauk (en ruso) 8 (5): 133-140.
- Kurzanov, Efimov, y Gubin, 2003. New archosaurs from the Jurassic of Siberia and Mongolia. Paleontological Journal 37(1):53-57.
- Chure, Daniel J.; Litwin, Ron; Hasiotis, Stephen T.; Evanoff, Emmett; Carpenter, Kenneth (2006). «The fauna and flora of the Morrison Formation: 2006». En Foster, John R.; and Lucas, Spencer G. (eds.), ed. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, Nuevo México: New Mexico Museum of Natural History and Science. pp. 233-248.
- Molnar, R.E., Kurzanov, S.M. y Dong, Z. (1990). Carnosauria, In Weishampel, D.B., Dodson, P., and Osmolska, H., eds., The Dinosauria, Berkeley: University of California Press, p. 169-209.
- Madsen and Welles, 2000. Ceratosaurus (Dinosauria, Theropoda) a revised osteology. Miscellaneous Publication 00-2 Utah Geological Survey, 80 págs.
- Newman, B.H. 1970. Stance and gait in the flesh-eating Tyrannosaurus. Biological Journal of the Linnean Society. 2: 119-123.
- Planeta - De Agostini, 1993. Dinosaurios, descubre los gigantes del mundo preshistórico Vol.1, VV.AA. ed.
- Fastovsky, David E.; y Smith, Joshua B. (2004). "Dinosaur Paleoecology", en The Dinosauria (2º). 614–626.
- Kenneth, Carpenter; Sanders, Frank; McWhinney, Lorrie A.; Wood, Lowell (2005). «Evidence for predator-prey relationships: Examples for Allosaurus and Stegosaurus» (pdf). En Carpenter, Kenneth (ed.), ed. The Carnivorous Dinosaurs. Bloomington e Indianapolis: Indiana University Press. pp. 325-350. ISBN 0-253-34539-1. Archivado desde el original el 17 de diciembre de 2008. Consultado el 16 de octubre de 2008.
- Tim Haines, ed. BBC Worldwide Ltd, 1999. Walking with Dinosaurs.
- Foster, John (2007). «Allosaurus fragilis». Jurassic West: The Dinosaurs of the Morrison Formation and Their World (en inglés). Bloomington, Indiana: Indiana University Press. pp. 170-176. ISBN 978-0-253-34870-8. OCLC 77830875.
- Stevens, Kent A. (2006). «Binocular vision in theropod dinosaurs». Journal of Vertebrate Paleontology 26 (2): 321-330. doi:10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2.
- Christiansen, Per (1998). «Strength indicator values of theropod long bones, with comments on limb proportions and cursorial potential» (pdf). Gaia 15: 241-255. ISSN 0871-5424. Archivado desde el original el 31 de octubre de 2008.
- Frazzetta, T.H.; Kardong, KV (2002). «Prey attack by a large theropod dinosaur». Nature 416 (6879): 387-388. PMID 11919619. doi:10.1038/416387a.
- Rayfield, Emily J.; Norman, D. B.; Upchurch, P. (2002). «Prey attack by a large theropod dinosaur: Response to Frazzetta and Kardong, 2002». Nature 416 (6879): 388. doi:10.1038/416388a.
- Lee, Andrew H.; and Werning, Sarah (2008). «Sexual maturity in growing dinosaurs does not fit reptilian growth models». Proceedings of the National Academy of Sciences 105 (2): 582-587. PMID 18195356. doi:10.1073/pnas.0708903105.
- Loewen, Mark A.; Carrano, Matthew T.; Sampson, Scott D. (2002). «Ontogenetic changes in hindlimb musculature and function in the Late Jurassic theropod Allosaurus''». Journal of Vertebrate Paleontology 22 (3, Suppl.): 80A.
- Farlow, James O. (1976). «Speculations about the diet and foraging behavior of large carnivorous dinosaurs». American Midland Naturalist 95 (1): 186-191. doi:10.2307/2424244.
- Tanke, Darren H.; and Currie, Philip J. (1998). «Head-biting behavior in theropod dinosaurs: Paleopathological evidence» (pdf). Gaia (15): 167-184. ISSN 0871-5424. Archivado desde el original el 11 de enero de 2006.
- Currie, Philip J. (1999). «Theropods». En Farlow, James; y Brett-Surman, M.K. (eds.), ed. The Complete Dinosaur. Indiana: Indiana University Press. p. 228. ISBN 0253213134.
- Roach, Brian T.; y Brinkman, Daniel L. (2007). «A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs». Bulletin of the Peabody Museum of Natural History 48 (1): 103-138. doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2.
- Goodchild Drake, Brandon (2004). «A new specimen of Allosaurus from north-central Wyoming». Journal of Vertebrate Paleontology 24 (3, Suppl.): 65A.
- Bakker, Robert T.; and Bir, Gary (2004). «Dinosaur crime scene investigations: theropod behavior at Como Bluff, Wyoming, and the evolution of birdness». En Currie, Philip J.; Koppelhus, Eva B.; Shugar, Martin A.; y Wright, Joanna L. (eds.), ed. Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Bloomington e Indianapolis: Indiana University Press. pp. 301-342. ISBN 0-253-34373-9.
- Rothschild, B., Tanke, D. H., and Ford, T. L., 2001, Theropod stress fractures and tendon avulsions as a clue to activity: en: Mesozoic Vertebrate Life, editado por Tanke, D. H., y Carpenter, K., Indiana University Press, p. 331-336.
- Molnar, R. E., 2001, Theropod paleopathology: a literature survey: En: Mesozoic Vertebrate Life, editado por Tanke, D. H., y Carpenter, K., Indiana University Press, p. 337-363.
- Foster, John R. (2003). Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A. New Mexico Museum of Natural History and Science Bulletin, 23. Albuquerque, Nuevo México: New Mexico Museum of Natural History and Science. p. 29.
- Russell, Dale A. (1989). An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press. pp. 175-176. ISBN 1-55971-038-1.
- Carpenter, Kenneth (2006). «Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus» (pdf). En Foster, John R.; and Lucas, Spencer G. (eds.), ed. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, Nuevo México: New Mexico Museum of Natural History and Science. pp. 131-138. Archivado desde el original el 2 de diciembre de 2007.
- Mateus, Octávio (2006). «Jurassic dinosaurs from the Morrison Formation (USA), the Lourinhã and Alcobaça Formations (Portugal), and the Tendaguru Beds (Tanzania): A comparison». En Foster, John R.; y Lucas, Spencer G. (eds.), ed. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, Nuevo México: New Mexico Museum of Natural History and Science. pp. 223-231.
- Chure, Daniel J.; Fiorillo, Anthony R.; Jacobsen, Aase (2000). «Prey bone utilization by predatory dinosaurs in the Late Jurassic of North America, with comments on prey bone use by dinosaurs throughout the Mesozoic» (pdf). Gaia 15: 227-232. ISSN 0871-5424. Archivado desde el original el 19 de julio de 2011.
- «Utah Symbols – State Fossil». State of Utah. Archivado desde el original el 22 de octubre de 2007. Consultado el 30 de setiembre de 2007.
- Glut, Donald F.; Brett-Surman, Michael K. (1997). «Dinosaurs and the media». En Farlow, James; and Brett-Surman, Michael K. (eds.), ed. The Complete Dinosaur. Bloomington e Indianapolis: Indiana University Press. pp. 675-706. ISBN 0-253-33349-0.
- Lambert, Dinosaur Data Book, p. 299.
- Return to the Valley, a documentary on the Region 1 Valley of Gwangi DVD.
- Canal History. «Jurassic Fight Club: Most Lethal Dinosaur». Consultado el 14 de julio de 2010. (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Marcot, Molly (8 de septiembre de 2011). «Dr. Dinosaur». The Diamondback (The University of Maryland). Archivado desde el original el 18 de enero de 2012. Consultado el 20 de octubre de 2011.
Véase también
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikimedia Commons alberga una galería multimedia sobre Allosaurus.
Wikimedia Commons alberga una galería multimedia sobre Allosaurus. Wikispecies tiene un artículo sobre Allosaurus.
Wikispecies tiene un artículo sobre Allosaurus.- Datos del Allosaurus en Duiops.net
- Allosaurus information sheet at Enchanted Learning (en inglés)
- Allosaurus brief fact sheet at Enchanted Learning (en inglés)
- Dinosaur bites back Noticia en la BBC (en inglés)
- Sobre La Balada del Gran Al, un especial de la serie de la BBC Walking with Dinosaurs que recrea la vida de un joven Allosaurus (en inglés)
- Lista de nombres asignados en algún momento al género Allosaurus Archivado el 24 de marzo de 2005 en Wayback Machine. (en inglés)
- Lusodinos, dinosaurios en Portugal (en portugués)
- Allosaurus en Dinodata (en inglés, se requiere registrarse en forma gratuita)
- Allosaurus dentro de Sauropodomorpha en The Thescelsosaurus (en inglés)
- Significado y pronunciación del nombre (en inglés)
- Allosaurus en The Theropod Database (en inglés)
| Control de autoridades |
|
|---|
 Datos: Q14400
Datos: Q14400- Multimedia: Allosaurus / Q14400
- Especies: Allosaurus