Brachypodium pinnatum
Brachypodium pinnatum, le brachypode penné, est une espèce de plantes monocotylédones de la famille des Poaceae (graminées), sous-famille des Pooideae, originaire des régions tempérées de l'hémisphère nord.
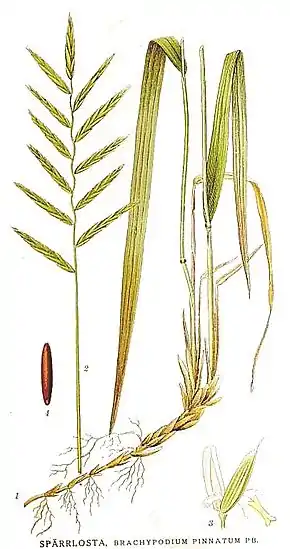
C'est une plante herbacée vivace, rhizomateuse, dont les tiges (chaumes) peuvent atteindre de 30 à 120 cm de long. L'inflorescence en grappe spiciforme regroupe des épillets longs pouvant compter plus d'une vingtaine de fleurons.

Depuis les années 1980 au moins, cette espèce se montre dans certaines circonstances (eutrophisation de pelouses calcicoles, et de plus en plus souvent, envahissante au détriment de la diversité biologique[1] de milieux auparavant parfois très riches en biodiversité (coteaux calcaires, certaines prairies...).
Description
Brachypodium pinnatum est une plante herbacée, vivace, cespiteuse, qui peut atteindre 70 cm à 1 mètre (voire jusqu'à 1,20 m) de haut.
Feuilles : elles sont de couleur verte (vert clair à jaunâtre parfois glaucescent).
Elles sont raides, dures, planes et larges de 4 à 6 mm. Leur extrémité peut être légèrement enroulée. Leurs nervures sont inégales, « quelques-unes saillantes et espacées, les autres fines et rapprochées »[2] ;
Tiges : elles sont raides, simples et dressées ; nues au sommet et portant 3 à 6 nœuds (le nœud supérieur étant situé au milieu ou au-dessus du milieu de la tige) ; ces nœuds sont pubescents, de même souvent que les feuilles, les gaines et les épillets ;
Épis : l'épi est distique ; formé de 6-12 épillets assez écartés ; les épillets (longs de 40 mm environ) sont dressés et légèrement arqués, ou plus ou moins étalés ;
L'inflorescence (de 5 à 25 cm de long) est dressée. Les fleurs sont bien visibles au moment de la floraison (juin à septembre) ainsi que les glumes (enveloppe des fleurs de graminées) et les glumelles (pubescentes ou non). Les glumelles inférieures sont un peu plus longues que les supérieures. Les arêtes ont la moitié ou le tiers de la longueur de leur glumelle.
Habitat
C'est une des herbacées typiques des ourlets préforestiers et des pelouses ou prairies de type mésotrophes et mésoxérophiles.
On la trouve sur sols plutôt calcaires et secs, mais parfois aussi en milieu légèrement acide.
Elle pousse en France jusqu'à 1 500 m d'altitude.
Distribution
Cette espèce serait originaire de zones steppiques et forestières (peu denses) du Sud de l'Eurasie[3].
Cette espèce, dont les semences ont pu être colportées par l'homme, est présente dans une grande partie de la zone tempérée de l'hémisphère nord, dont l'Amérique du Nord, et notamment les États-Unis, en particulier en Californie, dans le Massachusetts, et dans l'Oregon[4].
En France métropolitaine, cette espèce semble courante sur les friches et zones non cultivées dans tous les départements sauf peut-être dans le Massif central[5]. Elle est présente en Corse.
Deux cartes de répartition sont en cours de constitution en France, l'une par l'Inventaire national du patrimoine naturel (INPN)[6] et l'autre par Tela botanica[5].
Classification, synonymes
Cette espèce appartient au genre Brachypodium et à la famille des Poacées. Elle forme un groupe complexe avec une espèce très proche (Brachypodium sylvaticum[7] dont la définition taxonomique n'est sans doute pas stabilisée).
Elle a autrefois aussi été nommée (ou certaines de ses variétés) :
- Agropyron gracile (Leyss.) Chevall., 1827 (CD_NOM = 80462)[8]
- Agropyron pinnatum (L.) Chevall., 1827 (CD_NOM = 80490)[8]
- Avena laeta Salisb., 1796 (CD_NOM = 85285)[8]
- Brachypodium cespitosum (Host) Roem. & Schult., 1817 (CD_NOM = 86258)[8]
- Brachypodium loliaceum (Ten.) Roem. & Schult., 1817 (CD_NOM = 86278)[8]
- Brachypodium pinnatum subsp. caespitosum (Host) Hack., 1914 (CD_NOM = 132179)[8]
- Brachypodium pinnatum subsp. pinnatum (CD_NOM = 618888)[8]
- Brachypodium pinnatum var. cespitosum (Host) W.D.J.Koch, 1838 (CD_NOM = 144336)[8]
- Brachypodium pinnatum var. genuense (DC.) J.Woods, 1850 (CD_NOM = 144337)[8]
- Brachypodium pinnatum var. gracile (Leyss.) Asch. & Graebn., 1901 (CD_NOM = 144343)[8]
- Brachypodium pinnatum var. pinnatum (CD_NOM = 618887)[8]
- Brachypodium pinnatum proles cespitosum (Host) Rouy, 1913 (CD_NOM = 152845)[8]
- Brachypodium rupestre subsp. cespitosum (Host) H.Scholz, 1968 (CD_NOM = 132185)[8]
- Bromus caespitosus Host, 1809 (CD_NOM = 614951)[8]
- Bromus cespitosus Host (CD_NOM = 86566)[8]
- Bromus corniculatus Lam., 1779 (CD_NOM = 86576)[8]
- Bromus loliaceus Ten., 1815 (CD_NOM = 86660)[8]
- Bromus pinnatus var. tigurinus Suter, 1802 (CD_NOM = 144662)[8]
- Bromus pinnatus L., 1753 (CD_NOM = 86712)[8]
- Festuca fragilis (Schrank) Borkh. (CD_NOM = 98237)[8]
- Festuca gracilis Bernh., 1800 (CD_NOM = 98264)[8]
- Festuca pinnata (L.) Huds., 1778 (CD_NOM = 98445)[8]
- Triticum bromoides Weber, 1780 (CD_NOM = 127708)[8]
- Triticum genuense DC. (CD_NOM = 618889)[8]
- Triticum pinnatum (L.) Moench, 1777 (CD_NOM = 127782)[8]
Liste des sous-espèces et variétés
Selon Tropicos (15 octobre 2016)[9] (Attention liste brute contenant possiblement des synonymes) :
- sous-espèce Brachypodium pinnatum subsp. cespitosum (Host) Hack.
- sous-espèce Brachypodium pinnatum subsp. gracile (Leyss.) Soó
- sous-espèce Brachypodium pinnatum subsp. ligulatum Buia
- sous-espèce Brachypodium pinnatum subsp. phoenicoides (L.) Nyman
- sous-espèce Brachypodium pinnatum subsp. pinnatum
- sous-espèce Brachypodium pinnatum subsp. rupestre (Host) Schübl. & G. Martens
- variété Brachypodium pinnatum var. abbreviatum (Dumort.) Asch. & Graebn.
- variété Brachypodium pinnatum var. angustifolium Geiran
- variété Brachypodium pinnatum var. australe Godr.
- variété Brachypodium pinnatum var. cespitosum (Host) W.D.J. Koch
- variété Brachypodium pinnatum var. glabrum Rchb.
- variété Brachypodium pinnatum var. glaucovirens St.-Yves
- variété Brachypodium pinnatum var. ligulatum (Buia) Anghel & Beldie
- variété Brachypodium pinnatum var. loliaceum (Asch. & Graebn.) Domin
- variété Brachypodium pinnatum var. longifolium (Schousb.) Asch. & Graebn.
- variété Brachypodium pinnatum var. minus Schur
- variété Brachypodium pinnatum var. mucronatum (Willk.) Pérez Lara
- variété Brachypodium pinnatum var. phoenicoides (L.) Fiori
- variété Brachypodium pinnatum var. pubescens Rchb.
- variété Brachypodium pinnatum var. ramosum (L.) Fiori
- variété Brachypodium pinnatum var. rupestre (Host) Posp.
- variété Brachypodium pinnatum var. villosissimum Domin
- variété Brachypodium pinnatum var. vulgare Koch
Génétique
Comme d'autres graminées vivaces, cette espèce se montre capable de rapidement diversifier sa population par reproduction sexuée en phase de colonisation d'un milieu[10]. Une fois le milieu bien colonisé, les populations semblent surtout caractérisées par des tapis constitués de touffes « clonales » (c'est-à-dire issues d'une reproduction végétative), changement qui pourrait être expliqué par l'exclusion compétitive des génotypes moins adaptés au milieu[10].
Néanmoins, selon Wojciech Bąba et al., l'« ancienneté de l'habitat » (jusqu'à 300 ans dans certains cas étudiés) semble n'avoir qu'un effet marginal sur la diversité génétique (en termes de taux de loci polymorphes ou PPL)[10].
Étant donné leur importance écologique et pour le fourrage des herbivores, le genre Brachypodium fait depuis les années 2000 l'objet d'études cytomoléculaires. Ces études ont notamment permis de trouver chez B. pinnatum un génotype à 28 paires de chromosomes (au lieu de 18), mais il s'agit probablement d'un hybride entre B. distachyon (2n = 10) et B. pinnatum (2n = 18). Les caractéristiques génétiques de B pinnatum peuvent depuis 2009 notamment être comparées à celle d'une espèce modèle proche : Brachypodium distachyon[11],[12].
Écologie
Cette plante forme des touffes denses qui protègent le sol d'une insolation directe, dont par les UV solaires, ainsi que de l'érosion par les pluies. Mais certaines populations deviennent envahissantes, dans les contextes d'eutrophisation (tout comme deux autres graminées : Calamagrostis epigejos, Bromus inermis) ; par rapport à ces deux graminées et en condition expérimentale de monoculture, B. pinnatum profite beaucoup mieux des apports azotés (avec respectivement un gain de phytomasse de 1,1, 3,6 et 2,5 pour ces 3 espèces (sur un sol enrichi en azote, à raison de 50 kg/ha). Cependant une étude de l'agressivité respective de ces 3 plantes les unes par rapport aux autres quand elles sont en compétition (à trois ou deux par deux) montrent que B. pinnatum ne devient pas nécessairement dominant[13]. D'autres facteurs (notamment le microclimat et la pression d'herbivorie par les insectes) semblent également pouvoir contribuer à expliquer certaines invasions biologiques par cette espèce.
Elle sert potentiellement de nourriture à de nombreux mammifères herbivores, et c'est la plante hôte de certains lépidoptères ; c'est le cas par exemple du papillon Thymelicus lineola dont la chenille se nourrit de ce brachypode. Mais dans certaines circonstances les herbivores semblent l'éviter ; une explication à ce phénomène est que - comme quelques autres plantes (ex : prêles) ou graminées), certains phénotypes sont significativement moins appétents pour les herbivores (petits ou grands, tels le mouton) en raison d'une teneur accrue des feuilles et tiges en silice ;
une étude a exposé à des moutons cinq espèces de graminées plus ou moins enrichies en silice afin de mesurer l'impact de la silice en termes de préférences alimentaires des moutons[14]. Les préférences alimentaires des moutons ont peu changé pour un faible enrichissement, mais on a ensuite observé des différences significatives dans les préférences alimentaires, et les taux de broutage entre les espèces de graminées changeaient, d'autant plus que les herbacées avaient été exposées à des niveaux élevés de silice. L'impact était moindre pour Poa annua par rapport aux espèces moins appétentes (dont par exemple Brachypodium pinnatum et Festuca ovina)[14].
Les moutons nourris plus longtemps sur ces parcelles montrent une préférence pour l'espèce de graminée présentant les taux de silice foliaire les plus bas (à savoir P. annua)[14].
La plante se disperse via ses graines, notamment mangées par des oiseaux granivores et divers micro mammifères (rongeurs) ou transportées par des fourmis. Elle se reproduit aussi de manière végétative grâce à ses rhizomes (souche rampante).
Des expériences ont montré que cette espèce est douée d'une grande plasticité phénotypique et écologique[15], elle peut ainsi s'adapter à une vie à l'ombre ou en pleine lumière[15]. Certains phénotypes peuvent être localement très favorisés par les coupes rases ou désherbages, les incendies ou feux pratiqués dans certaines formes de sylviculture.
Caractère envahissant
Depuis quelques décennies, des pullulations de cette espèce sont constatées sur des pelouses calcaires dans tout l'hémisphère nord, avec alors une réduction importante de la biodiversité.
Ces pullulations peuvent apparaître avec les principaux modes de gestion (pâturage par ovins, gestion par brûlage dirigé, brûlage pastoral ou par la fauche).
Ce brachypode peut alors coloniser le milieu et constituer jusqu'à 80 % de la phytomasse du site.
Dans la plupart des cas (pays industrialisés, régions agricoles), cette pullulation semble pouvoir être expliquée par des apports anormaux et excessifs d'azote aéroporté (introduit dans le milieu par le mouvement des masses d'air et par le lessivage de cet air par les pluies[16].
Une étude suisse ayant porté sur les effets de 22 années consécutives de différents modes de gestion a montré qu'il est très favorisé par le brûlage, même « dirigé » dans le cadre d'un plan de gestion[17].
Facteurs d'explication
Plusieurs facteurs ou conditions ont été identifiés par les chercheurs, mais sont encore difficiles à hiérarchiser ; ils pourraient d'ailleurs aussi cumuler leurs effets. Ce sont notamment :
- des taux élevés de nitrates (et non d'autres éléments) semblent toujours ou très souvent associés aux phénomènes de pullulation de cette espèce, notamment sur sols "abandonnés"[18] ;
- Certains modes de gestion favorisant une ouverture du milieu avec faible concurrence d'autres plantes : Brachypodium pinnatum semble avoir les qualités d'une très bonne espèce pionnière, qui se montre volontiers dominante (ou co-dominante, par exemple avec Bromus erectus[19]. Le désherbage[20] et la gestion par le feu[17] peuvent fortement la favoriser, notamment en contexte d'eutrophisation (c'est-à-dire presque partout dans l'hémisphère nord où les pluies charrient des nitrates issus des épandages agricoles et issus des émissions de véhicules fonctionnant à l'essence, au fuel ou au gaz, via les NOx ;
- pression anormalement faible d'herbivorie par les insectes. C'est une autre hypothèse. Elle concerne notamment
- la pression par les chenilles des papillons de prairies (en forte régression) est aussi en cause, au moins dans certains cas. Dans les pays industrialisés et dans les régions agricoles, tous les papillons ont fortement régressé à partir des années 1960, probablement en raison de la régression de leurs habitats, mais aussi de la pollution diffuse et généralisée par les insecticides agricoles. La régression des chenilles et des orthoptères pourrait aussi contribuer à favoriser Brachypodium pinnatum[20]. Là où les orthoptères (sauterelles notamment) régressent, des espèces proches ont mieux et anormalement prospéré.
Une expérience conduite dans les Alpes françaises a montré que les brachypodes étaient effectivement limités par les insectes herbivores (sauterelles dans ce cas), mais seulement quand la végétation voisine a aussi été supprimée (par désherbage) ce qui laisse penser que la concurrence avec d'autres végétaux est aussi un facteur naturel de limitation du développement de cette espèce, même si elle se montre très compétitive. On a par ailleurs montré que de manière générale, l'herbivorie par les orthoptères varie fortement selon le microclimat, la luminosité et la fauche quand elle est pratiquée ; l'herbivorie augmente quand la température augmente et quand l'hygrométrie (vapeur saturante) diminue[21]. - l'herbivorie par les mammifères ; qui peut sélectionner des phénotypes de brachypodes plus riches en silice, moins appétents pour ces mammifères.
- la pression par les chenilles des papillons de prairies (en forte régression) est aussi en cause, au moins dans certains cas. Dans les pays industrialisés et dans les régions agricoles, tous les papillons ont fortement régressé à partir des années 1960, probablement en raison de la régression de leurs habitats, mais aussi de la pollution diffuse et généralisée par les insecticides agricoles. La régression des chenilles et des orthoptères pourrait aussi contribuer à favoriser Brachypodium pinnatum[20]. Là où les orthoptères (sauterelles notamment) régressent, des espèces proches ont mieux et anormalement prospéré.
Lutte contre les pullulations de B. pinnatum
- Le glyphosate (herbicide le plus vendu au monde, qui inhibe la germination des graines de B. pinnatum) a été utilisé pour tenter de maîtriser ce phénomène, sans résultats probants. Au Royaume-Uni, une étude a montré[22] que le recul n'est que provisoire ; B. pinnatum redevenant assez rapidement dominant, et d'autres espèces ubiquistes risquant d'occuper la place laissée libre par le désherbage chimique. Les auteurs de cette étude ont conclu qu'il ne faudrait pas utiliser le désherbage chimique pour contrôler B. pinnatum, sauf éventuellement dans le cadre d'un programme de lutte intégré dans un plan de gestion.
- Les retours d'expérience de cinq ans d'expérimentations de gestion des pelouses calcaires concernées par des pullulations de B. pinnatum aux Pays-Bas a montré qu'une fauche tardive d'automne n'empêchait pas une forte augmentation de cette espèce, et entrainait une diminution drastique de la diversité des espèces[23]. Alors que trois ans consécutifs de fauche à la mi-été ont diminué la phytomasse relative de cette espèce (elle passe de 80 % de la phytomasse totale à moins de 35 % en 5 ans), avec une forte augmentation de la diversité mesurée par l'indice de Shannon)[23]. En particulier, les plantes typiques des pelouses calcaires, les plus petites notamment réapparaissant avec ce régime de coupe[23].
Des analyses d'N et P faites dans la matière végétale (au-dessus du sol ou en dessous) n'ont pas mis en évidence de carences nutritionnelles dues à la coupe d'été ; et on n'a pas trouvé de lien entre les opérations d'exportation d'azote et la dominance de Brachypodium[23]. Il a été suggéré que la diminution de la quantité d'hydrates de carbone non structuraux dans les rhizomes de Brachypodium pourrait être un facteur important de cette réduction de leur croissance[23].
Statut, pressions, réponses
Même si les prairies qui l'abritaient ont beaucoup régressé en raison de l'intensification de l'agriculture (élevage hors-sol) et de l'urbanisation ou la périurbanisation, cette espèce n'est pas considérée comme menacée ou vulnérable, et elle ne fait pas l'objet de mesures de protection particulières.
Notes et références
- R Bobbink, JH Willems (1987), Increasing dominance of Brachypodium pinnatum (L.) beauv. in chalk grasslands: A threat to a species-rich ecosystem, Biological Conservation ; Volume 40, Issue 4, 1987, pages 301–314 ; Elsevier
- Fiche Tela botanica
- Fiche Brachypodium pinnatum Boratela
- « Brachypodium pinnatum (L.) P. Beauv. Heath false brome », département de l'Agriculture des États-Unis (consulté le ).
- Tela botanica, Brachypodium pinnatum (L.) P.Beauv. Poaceae ; chorologie départementale
- Carte de répartition, INPN / Muséum national d'Histoire naturelle [Ed]. 2003-2013., site Web : http://inpn.mnhn.fr. consulté 2013-09-07
- Beata Paszko (2007), The differing characteristics of Brachypodium pinnatum (L.) P. Beauv. and B. sylvaticum (Huds.) P. Beauv. ; Biodiv. Res. Conserv. 5-8: 11-16
- Citation : Muséum national d'Histoire naturelle [Ed]. 2003-2013. Inventaire national du patrimoine naturel, site Web : http://inpn.mnhn.fr. Le 7 septembre 2013.
- Tropicos.org. Missouri Botanical Garden., consulté le 15 octobre 2016
- Wojciech Bąba, Marzena Kurowska, Agnieszka Kompała-Bąba, Adam Wilczek, Joanna Długosz, Iwona Szarejko (2012), Genetic diversity of the expansive grass Brachypodium pinnatum in a changing landscape: Effect of habitat age ; Flora - Morphology, Distribution, Functional Ecology of Plants, Volume 207, Issue 5, mai 2012, pages 346-353 (résumé)
- Wolny E, Hasterok R (2009, Comparative cytogenetic analysis of the genomes of the model grass Brachypodium distachyon and its close relatives ; Ann Bot. Oct 2009 ;104(5):873-81. doi: 10.1093/aob/mcp179. Epub 2009-07-25 (résumé)
- Breda E, Wolny E, Hasterok R. (2012), Intraspecific polymorphism of ribosomal DNA loci number and morphology in Brachypodium pinnatum and Brachypodium sylvaticum ; Cell Mol Biol Lett. 2012 Dec;17(4):526-41. Epub 2012-08-15 (résumé).
- Petr Holub, Ivan Tůma, Karel Fiala (2012), The effect of nitrogen addition on biomass production and competition in three expansive tall grasses ; Environmental Pollution, Volume 170, Nov. 2012, Pages 211-216 (résumé)
- Fergus P. Massey, Kate Massey, A. Roland Ennos, Sue E.Hartley (2009), Impacts of silica-based defences in grasses on the feeding preferences of sheep ; Basic and Applied Ecology, Volume 10, Issue 7, October 2009, Pages 622-630 (résumé)
- Andrea Mojzes, Tibor Kalapos, Klára Virágh (2003), Plasticity of leaf and shoot morphology and leaf photochemistry for Brachypodium pinnatum (L.) Beauv. growing in contrasting microenvironments in a semiarid loess forest-steppe vegetation mosaic ; Flora - Morphology, Distribution, Functional Ecology of Plants, Volume 198, Issue 4, Pages 304-320 (résumé)
- Audra Hurst, Elizabeth John (1999), The biotic and abiotic changes associated with Brachypodium pinnatum dominance in chalk grassland in south-east England ; Biological Conservation, Volume 88, Issue 1, avril 1999, Pages 75-84
- Barbara Köhler, Andreas Gigon, Peter J. Edwards, Bertil Krüsi, Regula Langenauer, André Lüscher, Peter Ryser (2005), Changes in the species composition and conservation value of limestone grasslands in Northern Switzerland after 22 years of contrasting managements ; Perspectives in Plant Ecology, Evolution and Systematics, Volume 7, Issue 1, 2005-03-31, Pages 51-67 (résumé)
- Bonanomi, G., Caporaso, S., & Allegrezza, M. (2006), Short-term effects of nitrogen enrichment, litter removal and cutting on a Mediterranean grassland. Acta Oecologica, 30(3), 419-425.
- Grime, J. P., Shacklock, J. M. L., & Band, S. R. (1985). Nuclear DNA contents, shoot phenology and species co‐existence in a limestone grassland community. New Phytologist, 100(3), 435-445.
- Emmanuel Corcket, Ragan M. Callaway, Richard Michalet (2003) Insect herbivory and grass competition in a calcareous grassland: results from a plant removal experiment ; Acta Oecologica, Volume 24, Issue 3, July 2003, Pages 139-146 (résumé)
- Emmanuel Corcket, Ioana Chintauan-Marquier, Ragan M Callaway, Richard Michalet (2002), Sélectivité et variations environnementales de l'herbivorie par les Orthoptères (= Species-dependence and environmental changes in grasshoppers herbivory) ; Comptes Rendus Biologies, Volume 325, Issue 2, Février 2002, Pages 155-164 (résumé)
- Audra Hurst, Elizabeth John (1999), The effectiveness of glyphosate for controlling Brachypodium pinnatum in chalk grassland ; Biological Conservation, Volume 89, Issue 3, aout 1999, Pages 261-265 (résumé)
- R. Bobbink, J.H. Willems (1991), Impact of different cutting regimes on the performance of Brachypodium pinnatum in Dutch chalk grassland ; Biological Conservation, Volume 56, Issue 1, Pages 1-21 (résumé)
Voir aussi
Articles connexes
Bibliographie
- (en) Audra Hurst, Elizabeth John (1999), The biotic and abiotic changes associated with Brachypodium pinnatum dominance in chalk grassland in south-east England ; Biological Conservation, Volume 88, Issue 1, avril 1999, Pages 75–84
- (en) Reinhard Bornkamm (2006), Fifty years vegetation development of a xerothermic calcareous grassland in Central Europe after heavy disturbance ; Flora - Morphology, Distribution, Functional Ecology of Plants, Volume 201, Issue 4, 16 June 2006, Pages 249-267 (résumé)
Liens externes
- (en) Référence JSTOR Plants : Brachypodium pinnatum (consulté le )
- (en) Référence Catalogue of Life : Brachypodium pinnatum (L.) P.Beauv. (consulté le )
- (en) Référence Flora of China : Brachypodium pinnatum (consulté le )
- (en) Référence Kew Garden World Checklist : Brachypodium pinnatum (L.) P.Beauv., Ess. Agrostogr.: 101, 155 (1812) (consulté le )
- (en) Référence The Plant List : Brachypodium pinnatum (L.) P.Beauv. (source : KewGarden WCSP) (consulté le )
- (en) Référence Tropicos : Brachypodium pinnatum (L.) P. Beauv. (+ liste sous-taxons) (consulté le )
- (en) Référence uBio : Brachypodium pinnatum (L.) P. Beauv., 1812 (consulté le )
- (en) Référence World Register of Marine Species : espèce Brachypodium pinnatum (L.) Beauv. (consulté le )
- (fr) Boratela, site consacré aux Poacées, avec clé d'identification
- (fr) Fiche INPN (Muséum national d'histoire naturelle)
- (fr) Fiche Tela botanica
- (en) Tor Grass page from the Flora of Northern Ireland site.
- (en) Brachypodium.org
- (en) The « International Brachypodium Initiative »
 Portail de la botanique
Portail de la botanique  Portail de la protection des cultures
Portail de la protection des cultures  Portail des Poaceae
Portail des Poaceae