Ajnabia
Ajnabia odysseus

Longrich et al., 2021
Longrich et al., 2021
Ajnabia est un genre éteint de dinosaures hadrosauridés, classé dans la sous-famille des Lambeosaurinae. Ses restes ont été retrouvés dans une carrière de phosphate du bassin d’Ouled Abdoun au Maroc dans des sédiments marins datant du Maastrichtien supérieur (Crétacé supérieur) il y a 67 à 66 millions d’années. L’espèce type et seule espèce connue, Ajnabia odysseus, a été décrite et nommée en 2021 par Nicholas R. Longrich, Xavier Pereda Suberbiola, R. Alexander Pyron, et Nour-Eddine Jalil[N 1]. Ajnabia est le premier hadrosauridé découvert en Afrique. Sa découverte sur ce continent a été une surprise pour les paléontologues car au Crétacé l'Afrique était isolée du reste du monde par des barrières marines, de sorte que les hadrosaures étaient supposés avoir été incapables d'atteindre ce continent. Une analyse phylogénétique place Ajnabia au sein d’un nouveau clade de lambeosaurinés, les Arenysaurini, dont tous les autres représentant connus proviennent d’Europe. Les données biogéographiques suggèrent que les lambeosaurinés se sont dispersés en Afrique depuis l’Asie via l’Europe. Cette dernière était un archipel durant le Crétacé supérieur et les ancêtres d’Ajnabia auraient atteint l’Afrique depuis l’île Ibero-Armoricaine par dispersion océanique, les hadrosaures nageant ou dérivant sur des radeaux naturels sur plusieurs centaines de kilomètres[1].
Étymologie
Le nom du genre vient de l’arabe ajnabi signifiant « étranger », une allusion au fait que cet hadrosaure appartient à une lignée de dinosaures ayant migrée en Afrique depuis l’Europe. L’épithète spécifique se réfère à Odysseus, le nom en Grec ancien d’Ulysse, héros grec et voyageur marin légendaire[1],[2],[3]
Description
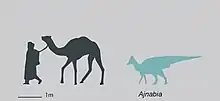
Ajnabia n’est connu que par des restes fragmentaires. L’holotype MHNM KHG 222 est composé d’un maxillaire gauche partiel ayant préservé de nombreuses dents, un fragment de maxillaire droit, et un fragment de dentaire droit. Le maxillaire le plus complet mesure 15,6 cm de long, mais il est cassé antérieurement et dorsalement. La diminution de la taille des alvéoles et le rétrécissement du maxillaire en avant suggèrent qu’il manque 2 à 3 cm d’os en longueur. L’animal était petit pour un hadrosauridé mais ses os ne présentent pas la texture osseuse d’un individu juvénile. S'il s'agit d'un adulte, il serait alors l'un des plus petits sinon le plus petit des Hadrosauridae connus. Bien que les vestiges de l’animal soient très incomplets, le maxillaire d’Ajnabia présente une combinaison unique de caractères. Le processus prémaxillaire du maxillaire est fortement incliné vers le bas dans des proportions comparables à ce qui est observé chez Canardia, Corythosaurus, Amurosaurus et Magnapaulia. Le maxillaire possède un foramen hypertrophié en dessous de l’articulation jugale et un foramen accessoire plus petit en dessous de celui-ci. La partie postérieure du maxillaire montre une expansion transversale extrême, formant un large plateau ectoptérygoïde qui fait saillie latéralement dans la même mesure que l’articulation jugale. Cette forte expansion transversale est absente chez les hadrosauroïdes basaux et les hadrosauridés à l’exception des lambeosaurinés européens comme Canardia et le lambeosauriné de Serrat del Rostiar. Latéralement, le plateau ectoptérygoïde porte une crête ectoptérygoïde massive, placée bas sur le maxillaire. En vue latérale, cette crête ectoptérygoïde est légèrement inclinée dorsalement par rapport au bord dentigère. Parmi les lambeosaurinés, cette condition est partagée avec Canardia et le lambeosauriné de Serrat del Rostiar. Comme chez ce dernier, la crête ectoptérygoïde est réduite antérieurement, ne touchant pas l’articulation jugale. La crête ectoptérygoïde est projetée loin et délimitée du corps du maxillaire par un surplomb, lui donnant une forme de lèvre épaisse.
Les couronnes des dents maxillaires présente un apex recourbé médialement. La carène médiane est décalée postérieurement, un caractère primitif perdu chez les hadrosauridés mais réacquis chez Ajnabia et chez les lambéosaurinés européens. Les denticules sont fortement réduits, un caractère dérivé présents chez les lambéosaurinés européens ainsi que chez Hypacrosaurus stebingeri et Edmontosaurus. Il n’y a pas de carènes accessoires. Les dents de la mâchoire inférieure ne sont pas encore connues[1].
Phylogénie
L’analyse phylogénétique conduite par Longrich et des collègues suggère qu’Ajnabia est étroitement apparenté aux Lambeosaurinae d’Europe avec lesquels il forme le clade des Arenysaurini. Au sein de ces derniers, Longrich et al. incluent des hadrosauridés Européens que d’autres auteurs ont rangé dans des clades distincts de lambéosaurinés (Aralosaurini[4], Tsintaosaurini[5],[6],[7], Parasaurolophini[8],[9], et Lambeosaurini[4] ,[9]).
Ci-dessous le cladogramme des lambéosaurinés publié par Longrich et al.[1].
| Hadrosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paléoécologie

Le spécimen holotype, MHNM KHG 222, a été trouvé dans les phosphates du bassin d'Ouled Abdoun à Sidi Chennane (région de Béni Mellal-Khénifra, Province de Khouribga) dans le centre-nord du Maroc. Les phosphates comprennent des sables phosphatés, des marnes et des calcaires déposés dans un milieu marin proche du rivage, une baie qui était ouverte vers l’Atlantique. Leur âge s’étend du Maastrichtien à l’Éocène moyen. Ajnabia provient d’un niveau appelé couche supérieure III, daté du Maastrichtien terminal par la faune d’Elasmobranchii associée. La chimiostratigraphie isotopique du carbone et de l’oxygène indique que les organismes fossiles de ce niveau ont vécu durant le dernier million d’années du Crétacé[1]. La faune marine comprends de nombreuses espèces de requins et de raies, des poissons osseux parmi lesquels un coelacanthe Mawsoniidae[10], le lézard marin Pachyvaranus[11], de nombreux mosasaures (Mosasaurus[12],[13], Prognathodon[14], Halisaurus[15], Eremiasaurus[16], Globidens[14],[17], Carinodens[18], Gavialimimus[19], Pluridens[20], Xenodens[21], et Thalassotitan[22]), l’élasmosaure Zarafasaura[23], les tortues marines géantes Ocepechelon[24] et Alienochelys[25], et le crocodylien gavialoïde Ocepesuchus[26]. Plusieurs ptérosaures ont également été identifiés (Phosphatodraco, Simurghia, Alcione, Barbaridactylus, et Tethydraco)[27],[28]. Bien que rares, des fossiles d’animaux vivant sur le continent sont régulièrement découverts dans ces couches marines, notamment l’abélisauridé Chenanisaurus barbaricus[29] et un titanosaure non nommé[30]. Ils correspondent, tout comme les fossiles d’Ajnabia, à des restes de cadavres transportés par les fleuves jusqu’à la mer. Ils donnent un aperçu de la diversité des dinosaures d’Afrique juste avant l’événement d’extinction Crétacé-Paléogène[1].
Paléobiogéographie
Sur la base des relations de parenté d'Ajnabia avec des dinosaures d'Europe et des reconstitutions des continents et des mers du Crétacé supérieur, Longrich et des collègues ont proposé que la dispersion des Lambeosaurinae en Afrique du Nord s'est probablement produite via la dispersion océanique, les hadrosaures nageant ou dérivant entre l'Europe (qui était alors un archipel) et l'Afrique du Nord. La dispersion des dinosaures entre les différentes îles européennes a pu se faire soit à travers des ponts terrestres, soit en franchissant des barrières marines, ou les deux. Les mers peu profondes qui séparaient les îles européennes auraient pu être comblées par des liaisons terrestres intermittentes, en particulier lors d’événements de régression marine. Le niveau de la mer au Crétacé supérieur était d’environ 100 m au-dessus des niveaux actuels, mais d’importantes baisses du niveau marin (>25 m) se sont produites durant de brèves périodes (≤1 Ma) au Santonien moyen, au Campanien moyen, et près de la limite Campanien-Maastrichtien, entraînés par la glaciation de l’Antarctique. Si la dispersion des dinosaures entre les différentes îles européennes peut s’expliquer par de multiples possibilités, pour Longrich et des collègues, les reconstructions paléogéographiques montrent que la dispersion des lambéosaurinés en Afrique ne peut pas s’expliquer par l’hypothèse des ponts terrestres. La mer de Téthys aurait isolé l’Afrique de l’Europe tout au long du Crétacé. Les événements de bas niveau marin auraient évidemment diminués les barrières marines séparant l’Afrique des îles européennes en rétrécissant les voies maritimes, en étendant les péninsules, et en exposant les récifs et les hauts-fonds, ce qui aurait facilité la dispersion. Mais ils n’auraient pas créé de ponts terrestres étant donné la présence de passages océaniques profonds entourant l’Afrique. De plus, les ponts terrestres suppriment les barrières géographiques à la dispersion et se caractérisent par des échanges fauniques importants et bidirectionnels. Ainsi, au Crétacé supérieur, l’existence d’une liaison terrestre entre l’Asie et l’Amérique du Nord a permis la migration de nombreux clades de dinosaures d’un continent vers l’autre et vice versa. Au Néogène, la formation de l’isthme de Panama a permis le Grand échange faunique interaméricain durant lequel les mammifères nord-américains se sont répandus en Amérique du Sud tandis que de nombreux taxons sud-américains ont migré en Amérique du Nord. Rien de tel n’est observé entre l’Europe et l’Afrique au Crétacé supérieur où les échanges semblent limités et unidirectionnels, suggérant l’existence de barrières géographiques[1].
Environ 500 km de pleine mer séparait les côtes du Maroc, où vivait Ajnabia, de l’île Ibero-Armoricaine (péninsule Ibérique + France), où vivaient les autres Arenysaurini. Selon Longrich et al., la dispersion de ces lambéosaurinés depuis cette île européenne vers l’Afrique du Nord se serait produite par voie maritime, les hadrosaures nageant ou dérivant sur des radeaux naturels (ces derniers, en fonction de leur taille, ne pouvant transporter que des œufs, de très jeunes individus, ou des adultes de tailles modestes) sur de grandes distances. Longrich et al. soulignent le fait que de nombreux animaux terrestres savent nager sur de longues distances. Les cerfs peuvent nager jusqu'à 5 km dans l’océan et 25 km dans les lacs pour se nourrir sur des îles. Les éléphants peuvent nager sur des distances approchant les 50 km. Les ours polaires peuvent nager sur plusieurs centaines de kilomètres, le record étant une nage de 9 jours et 687 km. Au Pléistocène, les éléphants ont colonisé des îles dont la Crète, Chypre, la Sardaigne et les Channel Islands de Californie, des cerfs se sont dispersés en Crète et en Sardaigne. Les hadrosaures étaient probablement mieux adaptés que les mammifères pour des traversées maritimes. Leurs grands pieds et leur queue en faisaient probablement de puissants nageurs. Des courants favorables ont également pu aidé à la dispersion (les hippopotames, qui sont de mauvais nageurs, ont colonisé des îles comme la Crète et Chypre, et ont traversé le canal du Mozambique jusqu’à Madagascar sur une distance de 300 km, peut être en dérivant). D’autres dinosaures communs à l’Afrique et à l’archipel européen du Crétacé supérieur, les titanosaures, étaient peut être particulièrement bien adaptés à la dérive. Bien que non spécialisés pour la vie dans l’eau, le cou et le dos pneumatisés des titanosaures les rendaient probablement flottants. De plus, une combinaison de vertèbres légères et d’os des membres lourds les aurait fait flotter debout, gardant la tête hors de l’eau même si l’animal était épuisé ou endormi. Ainsi, selon Longrich et des collègues, les échanges d’hadrosauridés et de titanosaures entre le Gondwana et la Laurasie suggèrent que la dispersion océanique était plus courante qu’on ne le pensait au Crétacé, et a joué un rôle important dans la structuration des faunes terrestres de cette époque[1].
Notes et références
Notes
- Une première description fut publiée en ligne en Novembre 2020, mais la publication officielle date d’Avril 2021.
Références
- (en) Nicholas R. Longrich, Xabier Pereda Suberbiola, R. Alexander Pyron et Nour-Eddine Jalil, « The first duckbill dinosaur (Hadrosauridae: Lambeosaurinae) from Africa and the role of oceanic dispersal in dinosaur biogeography », Cretaceous Research, vol. 120, , p. 104678 (ISSN 0195-6671, DOI 10.1016/j.cretres.2020.104678, lire en ligne, consulté le )
- Southworth, « Dinosaurs swam across oceans, landmark study reveals », sur www.telegraph.co.uk,
- « New fossil discovery suggests dinosaurs traveled across oceans », www.msn.com
- (en) A. Prieto-Marquez, F.M. Dalla Vecchia, R. Gaete et A. Galobart, « Diversity, relationships, and biogeography of the lambeosaurine dinosaurs from the European Archipelago, with description of the new aralosaurin Canardia garonnensis », PLOS ONE, vol. 8(7), , e69835 (PMID 23922815, PMCID 3724916, DOI 10.1371/journal.pone.0069835)
- (en) A. Prieto-Marquez et J.R. Wagner, « Pararhabdodon isonensis and Tsintaosaurus spinorhinus : a new clade of lambeosaurine hadroaurids from Eurasia », Cretaceous Research, vol. 30(5), , p. 1238-1246 (DOI 10.1016/j.cretres.2009.06.005)
- (en) S. Conti, B. Villa, A.G. Sellés, A. Galobart, M.J. Benton et A. Prieto-Marquez, « The oldest lambeosaurine dinosaur from Europe: insights into the arrival of Tsintaosaurini », Cretaceous Research, vol. 107, , p. 104286 (DOI 10.1016/j.cretres.2019.104286)
- (en) J.F. Serrano, A.G. Sellés, B. Villa, A. Galobart et A. Prieto-Marquez, « The osteohistology of new remains of Pararhabdodon isonensis sheds light into the life history and paleoecology of this enigmatic European lambeosaurine dinosaur », Cretaceous Research, vol. 118, , p. 104677 (DOI 10.1016/j.cretres.2020.104677)
- (en) P. Cruzado-Caballero, J.I. Canudo, M. Moreno-Azanza et J.I. Ruiz-Omeñaca, « New material and phylogenetic position of Arenysaurus ardevoli, a lambeosaurine dinosaur from the Late Maastrichtian of Arén (Northern Spain) », Journal of Vertebrate Paleontology, vol. 33(6), , p. 1367-1384 (DOI 10.1080/02724634.2013.772061)
- (en) A.A. Ramírez-Velasco, F.J. Aguillar, R. Hernández-Rivera, J.L. Gudiño Maussán, M.L. Rodriguez et J. Alvarado-Ortega, « Tlatolophus galorum, gen. et sp. nov., a parasaurolophini dinosaur from the upper Campanian of the Cerro del Pueblo Formation, Coahuila, northern Mexico », Cretaceous Research, vol. 126, , p. 104884 (DOI 10.1016/j.cretres.2021.104884)
- (en) P.M. Brito, D.M. Martill, I. Eaves, R.E. Smith et S.L.A. Cooper, « A marine Late Cretaceous (Maastrichtian) coelacanth from North Africa », Cretaceous Research, vol. 122, , p. 104768 (DOI 10.1016/j.cretres.2021.104768)
- (en) A. Houssaye, N. Bardet, J.C. Rage, X. Pereda Suberbiola, B. Bouya, M. Amaghzaz et M. Amalik, « A review of Pachyvaranus crassispondylus Arambourg, 1952, a pachyostotic marine squamate from the latest Cretaceous Phosphates of the margin of the Southern Tethys », Geological Magazine, vol. 148(2), , p. 237-249 (DOI 10.1017/S0016756810000580)
- (en) N. Bardet, X. Pereda Suberbiola, M. Iarochène, F. Bouyahyaoui, B. Bouya et M. Amaghzaz, « Mosasaurus beaugei Arambourg, 1952 (Squamata, Mosasauridae) from the Late Cretaceous phosphates of Morocco », Geobios, vol. 37(3), , p. 315-324 (DOI 10.1016/j.geobios.2003.02.006)
- (en) T.H. Rempert, A.P.M. Vinkeles Melchers, A.N. Rempert, M.R. Haque et A.R. Armstrong, « Occurrence of Mosasaurus hoffmannii Mantell, 1829 (Squamata, Mosasauridae) in the Maastrichtian Phosphates of Morocco », Journal of Paleontological Sciences, vol. 10, , p. 1-22 (lire en ligne)
- (en) N. Bardet, X. Pereda Suberbiola, M. Iarochène, M. Amalik et B. Bouya, « Durophagous Mosasauridae (Squamata) from the Upper Cretaceous phosphates of Morocco, with description of a new species of Globidens », Netherlands Journal of Geosciences, vol. 84(3), , p. 167-176 (DOI 10.1017/S0016774600020953)
- (en) N. Bardet, X. Pereda Suberbiola, M. Iarochène, B. Bouya et M. Amaghzaz, « A new species of Halisaurus from the Late Cretaceous phosphates of Morocco, and the phylogenetical relationships of the Halisaurinae (Squamata: Mosasauridae) », Zoological Journal of the Linnean Society, vol. 143, , p. 447-472 (DOI 10.1111/j.1096-3642.2005.00152.x)
- (en) A.R.H. Leblanc, M.W. Caldwell et N. Bardet, « A new mosasaurine from the Maastrichtian (Upper Cretaceous) phosphates of Morocco and its implications for mosasaurine systematics », Journal of Vertebrate Paleontology, vol. 32(1), , p. 82-104 (DOI 10.1080/02724634.2012.624145)
- (en) A. Le Blanc, S. Mohr et M. Caldwell, « Insights into the anatomy and functional morphology of durophagous mosasaurines (Squamata: Mosasauridae) from a new species of Globidens from Morocco », Zoological Journal of the Linnean Society, vol. 186(4), , p. 1026-1052 (DOI 10.1093/zoolinnean/zlz008)
- (en) A.S. Schulp, N. Bardet et B. Bouya, « A new species of the durophagous mosasaur Carinodens (Squamata, Mosasauridae) and additional material of Carinodens belgicus from the Maastrichtian phosphates of Morocco », Netherlands Journal of Geosciences, vol. 88(3), , p. 161-167 (DOI 10.1017/S0016774600000871)
- (en) C.R.C. Strong, M.W. Caldwell, T. Konishi et A. Palci, « A new species of longirostrine plioplatecarpine mosasaur (Squamata, Mosasauridae) from the Late Cretaceous of Morocco, with a re-evaluation of the problematic taxon Platecarpus ptychodon », Journal of Systematic Paleontology, vol. 18(21), , p. 1769-1804 (DOI 10.1080/14772019.2020.1818322)
- (en) N.R. Longrich, N. Bardet, F. Khaldoune, O. Khadiri Yazami et .-E. Jalil, « Pluridens serpentis, a new mosasaurid (Mosasauridae: Halisaurinae) from the Maastrichtian of Morocco and implications for mosasaur diversity », Cretaceous Resarch, vol. 126, , p. 104882 (DOI 10.1016/j.cretres.2021.104882)
- (en) N.R. Longrich, N. Bardet, A.S. Schulp et N.-E. Jalil, « Xenodens calminechari gen. et sp. nov., a bizarre mosasaurid (Mosasauridae, Squamata) with shark-like cutting teeth from the upper Maastrichtian of Morocco, North Africa », Cretaceous Resarch, vol. 123, , p. 104764 (DOI 10.1016/j.cretres.2021.104764)
- (en) R. Longrich, N-E. Jalil, F. Khaldoune, O.K. Yazami, X. Pereda-Suberbiola et N. Bardet, « Thalassotitan atrox, a giant predatory mosasaurid (Squamata) from the Upper Maastrichtian Phosphates of Morocco », Cretaceous Research, , p. 105315 (DOI 10.1016/j.cretres.2022.105315)
- (en) P. Vincent, N. Bardet, X. Pereda Suberbiola, B. Bouya, M. Amaghzaz et S. Meslouh, « Zarafasaura oceanis, a new elasmosaurid (Reptilia: Sauropterygia) from the Maastrichtian Phosphates of Morocco and the palaeobiogeography of latest Cretaceous plesiosaurs », Gondwana Research, vol. 19(4), , p. 1062-1073 (DOI 10.1016/j.gr.2010.10.005)
- (en) N. Bardet, N.-E. Jalil, F. De Lapparent De Broin, D. Germain, O. Lambert et M. Amaghzaz, « A Giant Chelonioid Turtle from the Late Cretaceous of Morocco with a Suction Feeding Apparatus Unique among Tetrapods », PLOS ONE, vol. 8(7), , e63586 (PMID 23874378, PMCID 3708935, DOI 10.1371/journal.pone.0063586)
- (en) F. Lapparent de Broin, N. Bardet, M. Amaghzaz et S. Meslouh, « A strange new chelonioid turtle from the latest Cretaceous phosphates of Morocco », Comptes Rendus Palevol, vol. 13(2), , p. 87-95 (DOI 10.1016/j.crpv.2013.07.008, lire en ligne)
- (en) S. Jouve, N. Bardet, N.-E. Jalil, X. Pereda Suberbiola, B. Bouya et M. Amaghzaz, « The oldest African crocodylian: phylogeny, paleobiogeography, and differential survivorship of marine reptiles through the Cretaceous-Tertiary Boundary », Journal of Vertebrate Paleontology, vol. 28(2), , p. 409-421 (DOI 10.1671/0272-4634(2008)28[409:TOACPP]2.0.CO;2)
- (en) X. Pereda Suberbiola, N. Bardet, S. Jouve, M. Iarochène, B. Bouya et M. Amaghzaz, « A new azhdarchid pterosaur from the Late Cretaceous phosphates of Morocco », dans Buffetaut, E. & Mazin, J.-M., Evolution and Palaeobiology of Pterosaurs, London, Geological Society of London, Special Publications, 217, (ISBN 978-1862391437), p. 80–90
- (en) N.R. Longrich, D.M. Martill, B. Andres et D. Penny, « Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary », PLOS Biology, vol. 16(3), , e2001663 (PMID 29534059, PMCID 5849296, DOI 10.1371/journal.pbio.2001663)
- (en) N.R. Longrich, X. Pereda Suberbiola, N.-E. Jalil, F. Khaldoune et E. Jourani, « An abelisaurid from the latest Cretaceous (late Maastrichtian) of Morocco, North Africa », Cretaceous Research, vol. 76, , p. 40-52 (DOI 10.1016/j.cretres.2017.03.021)
- (en) X. Pereda Suberbiola, N. Bardet, M. Iarochène, B. Bouya et M. Amaghzaz, « The first record of a sauropod dinosaur from the Late Cretaceous phosphates of Morocco », Journal of African Earth Sciences, vol. 40, , p. 81-88 (DOI 10.1016/j.jafrearsci.2004.07.002)
 Portail des dinosaures
Portail des dinosaures  Portail de la paléontologie
Portail de la paléontologie  Portail du Maroc
Portail du Maroc