Appareil stridulatoire de l'araignée
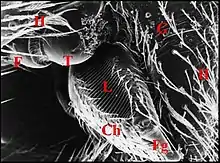
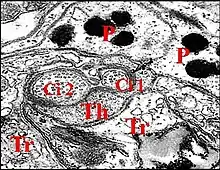
L'appareil stridulatoire des Araignées est une différenciation spéciale de leur tégument, paire, symétrique et pouvant occuper une position très variée sur le corps araneidien. Censé pour la plupart des auteurs produire des sons, il peut intervenir dans l'équilibration, ce que confirme l'étude ultrastructurale, notamment chez les Argyrodes (Theridiidae). Dans ce genre, son aire se localise au niveau du pédicule sur la partie postérieure de prosoma et l'avant de l'abdomen (ci-contre).

Introduction
Il est bien connu que des Myriapodes, certains Crustacés (Busnel, 1963) et surtout, un grand nombre d’ Insectes appartenant à des ordres variés (Homoptères, Orthoptères, Lépidoptères tels que Parnassius mnemosyne…) peuvent émettre des sons.
Divers Arachnides en produisent aussi mais ce phénomène est généralement insoupçonné car les vibrations sonores ne sont perceptibles par les Humains que dans de rares cas. Il est ainsi décrit chez les Scorpiones, les Solifuges (in Berland, 1932) et, en ce qui concerne plus directement cet article, dans l’ordre des Aranéides.
La production de sons par les Araignées a été constatée pour la première fois il y a plus de 130 ans, lorsque Wood-Mason (1876) [1]signala qu’une grande Mygale de l’Inde et Birmanie, Chilobrachys stridulans, peut être à la source d’un bruit que l’Homme perçoit à distance.
Il existe chez les Araignées trois méthodes pour produire des sons, ou, si l’on préfère, trois types d’émissions sonores : la vibration d’appendices, la percussion du substrat et la stridulation.
La vibration des pattes a été mise en évidence par Rovner (1980)[2] chez la grosse Araignée cosmotropicale Heteropoda venatoria (Eusparassidae). Elle se produirait aussi chez les Ctenidae dont le genre Phoneutria.
Encore appelée «tambourinage» («drumming» chez les auteurs anglophones), la percussion du substrat est réalisée avec les palpes, les pattes et (ou) l’abdomen.Elle a été découverte chez des Alopecosa nord-américaines et, par la suite, observée bien souvent chez d’autres Lycosidae, les Clubionidae, les Anyphaenidae, les Ctenidae, les Thomisidae et les Salticidae (revue in Uetz & Stratton, 1982)[3].
Stridulation
La stridulation, ou supposée telle, existerait dans près d’une trentaine de familles d’Araignées.
Selon Busnel (1963)[4], elle peut être définie chez tous les Arthopodes comme un moyen de production sonore qui met en jeu le frottement de deux parties rigides du corps l’une contre l’autre. Pourvues d’une surface tégumentaire spéciale, différenciée, ces deux parties constituent l’organe ou appareil stridulatoire.
Le son émis chez les Araignées n’est audible sans amplificateur que dans quelques cas privilégiés. L’oreille humaine le perçoit alors de très près (20 cm maximum) chez certains petits Theridiidae tels que Steatoda bipunctata (Meyer, 1928)[5], à la distance de 1 m. dans le cas de Micrathena gracilis (Araneidae)(Uetz & Stratton, 1982)[3] et jusqu’à 2 ou 3 ms lorsqu’une Mygale géante comme Theraphosa leblondi , de Guyane française, se met à striduler (Lopez, 1989)[6].
On le décrit de manière imagée comme un bourdonnement (Sicarius, Micrathena), un cliquetis métallique, un sifflement (Mygales), un grincement ou le bruit obtenu en frottant les dents d’un peigne avec l’ongle ou un couteau. Il peut être même enregistré et analysé au laboratoire où des sonogrammes et oscillogrammes ont été obtenus dans divers groupes araneidiens (Uetz & Stratton, 1982)[3].
Les vibrations qui le composent seraient transmises par l’air, par le substrat jouant le rôle d’une table d’harmonie, et perçues ensuite par les autres Araignées de même espèce, peut être aussi d’espèces différentes, ou par des prédateurs éventuels.
Chez les Araignées, cette même perception est dévolue à des organes sensoriels spéciaux, les trichobothries et les sensilles à fentes («slit sensilla») qui siègent sur les pattes. Barth (1967) l’a démontrée sur les plans éthologique et électrophysiologique.
La signification des sons produits est très discutée.
- Valeur sexuelle lorsque la stridulation n’existe que chez le mâle. Ils pourraient alors faciliter l’accouplement comme c’est le cas, semble-t-il, chez les Theridiidae Teutana grossa et Steatoda bipunctata (Gwinner-Hanke, 1970)[7].
- Valeur prémonitrice lorsque le son est émis par les deux sexes et semble représenter un moyen d’intimidation. Il protègerait l’Araignée contre ses ennemis naturels (Homme compris) en créant un effet de surprise. Une telle fonction semble à peu près certaine dans le cas particulier des Mygalomorphes et des Micrathena.
Quoi qu’il en soit, une stridulation effective n’est que rarement constatée «de auditu ». L’appellation pourtant très suggestive de certaines espèces d’Argyrodes (Theridiidae : A.stridulator, A.ululans) semble avoir été inspirée à leurs descripteurs, dont Lawrence (1937)[8], non par la perception d’un bruit qu’elles auraient émis, mais par l’observation de crêtes présumées stridulatoires («stridulatory ridges»)
Chez la plupart des Araignées, l’existence d’une stridulation est seulement présumée lorsqu’elles possèdent un appareil tégumentaire répondant à la définition générale de Busnel (1963)[4], censé stridulatoire et que l’on estime capable de produire des sons.
Anatomie générale et classification
L’appareil stridulatoire des araignées, nom que nous adoptons ici faute de termes plus appropriés, est une différenciation spéciale de leur tégument, paire, symétrique et pouvant occuper une position très variée sur le corps araneidien.
Comme chez les autres Arthropodes, il comporte toujours deux parties bien distinctes, dites «antagonistes», visibles à la loupe binoculaire et surtout, au microscope électronique à balayage (M.E.B.), moyen de recherche incomparable.
- La «lyre» ou «rape» («file» des auteurs de langue anglaise), noms désormais consacrés par l’usage, est fixe, passive et se présente comme un ensemble de rides, crêtes (Fig. 1 à 3,5), rugosités diverses ou même, de tigelles vibrantes.
- L'"archet", «peigne» ou «racloir» («scraper» des anglo-saxons), autres termes définitifs, est mobile, actif et formé par des dents, des épines, des tubercules chitineux (Fig.17 à 20) ou des soies rigides (Fig.8).
- L'archet frotte contre la lyre, ce jeu mécanique étant responsable soit d’une émission sonore présumée ou avérée, soit, du moins chez les Theridiidae Argyrodes, d’une stimulation nerveuse inédite et beaucoup plus subtile qui serait impliquée dans…l’équilibration (Lopez, 1988a ; Lopez, 1988b ; Lopez, 1994a ; Lopez, 1994b)[9],[10],[11],[12], de concert avec des organes lyriformes pédiculaires en « bourrelet », situés sur le pédicule et également étudiés chez les Meta (Lopez & Juberthie-Jupeau, 1994a et b).
Les deux parties peuvent siéger :
- sur un même appendice (surfaces segmentaires opposées d’une articulation de patte) (Rovner, 1975)[13],
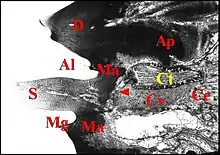
- sur deux appendices différents, tels que les pattes I et II, les deux chélicères, le bulbe copulateur et la gnathocoxa, le pédipalpe et la chélicère (Fig.), le pédipalpe et la patte I, peut être aussi les filières antérieures chez certaines Malkaridae (Forster & Platnick , 1984)[14],
- sur les deux grands tagmes corporels (céphalothorax ou prosoma et abdomen ou opisthosoma), notamment chez les Argyrodes (Fig.),
- sur le pédicule (pédicelle) et l’abdomen,
- sur un appendice (patte IV ou postérieure) et l’abdomen (Fig.).

Ainsi a-t-on pu définir plusieurs types ou catégories d’appareils stridulatoires, en se basant sur les positions respectives de la lyre et du peigne et en créant ainsi au moins 4 classifications :
- 4 types, classés de « a » à « d » par Berland (1932),
- 7 types, présentés sans ordre défini par Millot (1949), et
- 7 types encore, décrits et énumérés de « a » à « g » par Legendre (1963,1970[15]) dans son important travail, devenu classique et qui est basé sur l’examen de 74 espèces appartenant à une quinzaine de familles différentes.
- 8 types selon Uetz et Stratton (1982)[3] qui reprennent la classification de Legendre en lui adjoignant une catégorie particulière dans laquelle un seul et même appendice se trouve concerné (Rovner, 1975)[13].
Cas particuliers choisis
Sont présentés ici trois types d’appareils stridulatoires, « a », « d » et « g » dont la découverte et (ou) l’étude approfondie chez certaines espèces ont permis à A.Lopez de contribuer à leur connaissance.
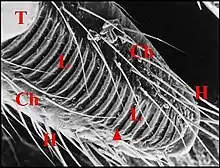
Dans tous les cas, les examens ont été réalisés à la loupe binoculaire, en microscopie photonique sur des coupes colorées par des méthodes histologiques de routine, et surtout, en microscopie électronique à balayage (M.E.B).
Il sera montré d'abord pour la première fois que le type « d »(chélicères-pédipalpes) est présent dans le genre Holocnemus (Pholcidae) et que le type « g » (quatrième paire de pattes, IV- opisthosoma) se retrouve chez quelques Araignées Gasteracanthines de la Guyane française[16]
Le plus important, celui des Argyrodes (« a »), dit aussi «prosoma-opisthosoma», sera ensuite développé séparément et en détails, ultrastructure comprise (microscopie électronique à transmission : M.E.T.)[9],[10],[11],[12].
Appareil stridulatoire d' Holocnemus pluchei
L’appareil stridulatoire de type " d " associe une lyre, portée par la chélicère, et un archet, situé sur le pédipalpe ou patte machoire.
Décrit pour la première fois par Simon (1893) chez l’Araignée sud-africaine Sicarius hahni (Sicariidae), il a été retrouvé ensuite chez les Scytodidae (Pickard-Cambridge, 1895), les Diguetidae de Californie (Chamberlin, 1924) et, beaucoup plus récemment, chez certains Pholcidae américains du genre Physocyclus (Gertsch, 1979,1982).

Il a été également observé dans cette dernière famille (Lopez, 1988) mais il s’agit en l’occurrence d’ Holocnemus pluchei (Scop.)[10],[17], un Pholcide " indigène " très répandu dans les garrigues héraultaises et jusque dans les habitations du Bittérois sensu lato, ainsi que leurs dépendances (Lopez, 1973). Cette découverte précède donc de plusieurs années le travail de Huber (1995)[18]. Holocnemus est fort intéressant aussi sur le plan histologique par ses curieuses cellules pigmentaires étoilées de siège profond, non épidermique (Lopez,)[19]. Des cellules pigmentaires enfouies analogues ont été également observées chez l'espèce américaine Diguetia canities. (Lopez)
Les exemplaires étudiés, uniquement au M.E.B., provenaient de Cessenon (Ouest de l’Hérault, Occitanie, France) , ainsi d’ailleurs que de Téboursouk (Tunisie), leur espèce présentant une aire de répartition circum-méditerranéenne très vaste.
La Lyre
Elle est située sur la face externe de la chélicère, plus précisément sur celle de son article basal, la tige ou paturon. Sous la loupe binoculaire, elle apparaît comme une bande brillante, de couleur rougeâtre, striée en travers et disposée verticalement selon le grand axe de la tige chélicérienne.
Au M.E.B., il s’agit d’une zone oblongue, entièrement glabre mais cernée par de nombreux poils, et déprimée en gouttière peu profonde (Fig.1 à 3, Fig.5). Elle porte sur toute sa surface une série de 30 à 40 crêtes parallèles, équidistantes, bifurquées parfois à leurs extrémités et séparées par des sillons uniformes qui ont tous la même longueur (Fig.3).

Cette lyre existe chez toutes les femelles, où son aspect ne varie guère (Fig.1 à 3), chez les mâles (Fig.4), les immatures où elle paraît déjà bien formée (Fig.5) et dont les crêtes semblent plus longues, un peu moins régulières, plus nombreuses et tendent à s’anastomoser.
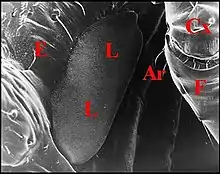
L’ archet
Il est représenté par le trochanter du pédipalpe (Fig.1 à 3). Entrant en rapport intime avec la lyre, cet article est beaucoup plus volumineux et renflé que chez Pholcus phalangioides, espèce très commune d’un genre voisin, dépourvue, pour sa part, d’appareil stridulatoire sur la tige chélicérienne à surface unie (Fig. 6). Il est fortement convexe et porte même à sa surface une petite saillie hémisphérique (Fig.2 : flèche) pouvant accroître les frottements exercés sur les rugosités de la chélicère
Appareil stridulatoire des Gasteracanthinae.
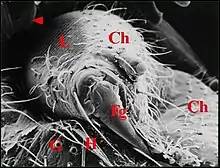
L’appareil stridulatoire du type " g " se compose d’un archet situé sur la 4e paire de pattes ambulatoires (P IV) et d’une lyre placée sur la face inférieure de l’abdomen. Carpenter (1898) l’a découvert chez Rhaebothorax brocchus et Falconer (1910) l’a retrouvé un peu plus tard en Angleterre chez une autre Erigoninae, Eboria caliginosa.
La lyre de ces Araignées est une surface rugueuse correspondant à l’ "opercule", "plaque" ou "couvercle" chitineux du poumon ou phyllotrachée ("booklung cover" des auteurs de langue anglaise). Le peigne qui balaye cette lyre est un relief en forme de dent aiguë siégeant sur le bord interne de la 4e hanche. Selon Savory (1928), ils n’existeraient que chez le mâle.
En dehors des Erigoninae, seuls cités par Legendre (1963), le type " g " n’a été retrouvé ultérieurement, et jusqu’à l' étude de Lopez (1988,) que chez une Lycoside, Pardosa fulvipes (Col.)(Kronestedt, 1973) et surtout, chez les Gasteracanthinae, sous famille d’Araneidae remarquables par leur "armature" épineuse, leurs couleurs souvent vives et l’aspect original des filières .
Hinton et Wilson (1970) précisent que l’archet ("scraper") des Gasteracanthinae est formé par des soies rigides dressées sur la base du fémur de P IV et que leur plaque pulmonaire est couverte de crêtes extrêmement fines et nombreuses ("file"). Leur description est basée sur une étude au M.E.B. concernant les femelles des Micrathena gracilis et Micrathena schreibersi, espèces américaines. De son côté, Levi (1985) présente une planche photographique montrant les appareils stridulatoires de Chaetacis aureola et Micrathena mitrata, le couvercle du poumon de la première ayant une forme quadrangulaire, celui de la seconde une forme arrondie.
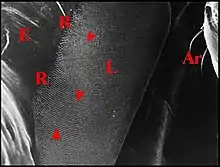
Lopez a examiné l’appareil stridulatoire du type " g " chez des Gasteracanthines de la Guyane [10]. Cette sous-famille est représentée dans notre département néotropical par de nombreuses espèces que les Zoologistes français, Arachnologues compris, semblent avoir pratiquement ignoré jusqu’à sa mise au point (Lopez, 1994c.)[16].
Chaetacis cornuta
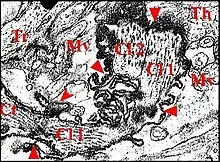
La femelle de Chaetacis cornuta (Taczanowski), seule étudiée, a des plaques pulmonaires allongées, ovalaires, brunâtres, disposées obliquement par rapport à l’axe longitudinal de l’abdomen (Fig.7). Chacune d’elles mesure environ 0,6 mm de long sur 0,2 mm de largeur maximum.
La Lyre
Il s’agit d’un ensemble de crêtes nombreuses (environ 150), fines, très serrées, légèrement incurvées et s’anastomosant, à leur extrémité, en un réseau "gaufré" (Fig.8). Cet aspect rappelle celui qu’offre une espèce voisine, Chaetacis aureola (C.L.Koch) au M.E.B. (Levi, 1985 : pl.3). Toutefois, sa plaque pulmonaire paraît plus petite et montre un moins grand nombre de crêtes que chez C. cornuta.
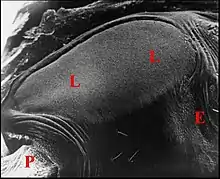
L’ archet
Il est constitué par deux soies courtes, raides, incurvées vers l’arrière et s’implantant sur la base fémorale (Fig.7,8) comme celles que Hinton et Wilson (1970)[20] ont figurées chez des Micrathena. Contrairement à ce qu’a écrit Levi (1985)[21], il ne s’agit donc pas de tubercules coxaux ou fémoraux.
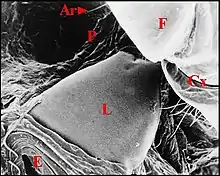
Micrathena schreibersi
Micrathena schreibersi (Perty) est la plus belle des Gastéracanthines guyanaises (Lopez, 1994c)[16].
Bien que Hinton et Wilson (1970)[20] ne l’aient pas spécifié, son mâle possède aussi un appareil stridulatoire.

La lyre
Elle correspond à la plaque pulmonaire, zone oblongue, d’un noir intense et luisant, arrondie à son extrémité postéro-externe, encochée, près du pédicule à l’antéro-interne (Fig.9), mesurant 0,6 mm dans sa plus grande longueur, 0,3 mm de large, et portant environ 150 crêtes qui sont parfois bifurquées (Fig.10).
L’archet
Il est encore formé par des soies fémorales de sorte que l’appareil stridulatoire rappelle, en plus petit, celui de la femelle, seule examinée par Hinton et Wilson (1970)[20].
Micrathena clypeata
Micrathena clypeata (Walckenaer), troisième et dernière Gastéracanthine, est une curieuse espèce, aplatie, foliacée, beaucoup plus répandue en Guyane (Lopez, 1994c)[16] que ne semble l’indiquer la citation trop restrictive de Levi (1985 : Saint Laurent du Maroni)[21].
La lyre
Elle reste confinée à la plaque pulmonaire, a une forme trapézoïdale (Fig.11) et mesure 0,6 mm de hauteur pour une base de 0,7 mm. Les crêtes y sont extrêmement nombreuses et si rapprochées (environ 3 µ) que leur dénombrement est impossible ; elles ne sont pas rectilignes et se dédoublent fréquemment pour s’unir en oblique (Fig.12).
L’ archet
Il est toujours fémoral et constitué par des soies (Fig.11).

L'Appareil stridulatoire des Argyrodes
L’un des appareils stridulatoires les plus répandus est le type « a », signalé pour la première fois par Westring (1843) sans toutefois le nommer ainsi, bien défini par Berland (1932) et surtout par Legendre (1963). Une «lyre» céphalothoracique y serait raclée par le «peigne» ou, mieux, l'archet abdominal.
On le connaît actuellement dans au moins 4 familles : les Theridiidae, les Clubionidae, les Gnaphosidae et les Hahniidae (Uetz et Stratton, 1982)[3].
Lopez l'a étudié chez les Theridiidae du genre Argyrodes dont il est l’une des nombreuses caractéristiques conférant à ces petites Araignées un intérêt majeur. Elles sont en effet remarquables aussi par leur dimorphisme sexuel céphalothoracique, en rapport avec l’existence d’une glande clypéale ou acronale n’existant que chez le mâle, par des glandes à soie originales, par le vif éclat métallique de leur abdomen, la forme curieuse de leurs cocons et par le fait qu’elles vivent généralement en kleptoparasites sur la toile d’autres Aranéides.
Un appareil assez voisin pourrait exister aussi dans le genre Asagena.
Au moins chez les Argyrodes, il s’associe aux organes lyriformes du pédicelle ou pédicule (Lopez, 1996) constituant avec ces derniers un complexe anatomo-fonctionnel sus-pédiculaire présumé stato-récepteur (Lopez, 1994a ; Lopez, 1994b)
Aspect au Microscope électronique à balayage
Le M.E.B. montre sans équivoque l'aspect spectaculaire de l'appareil stridulatoire des Argyrodes, constant et assez homogène malgré des variations (Lopez, 1988 ; Lopez, 1994a ; Lopez, 1994b ; Lopez, 1996) (Fig.1 à 5)
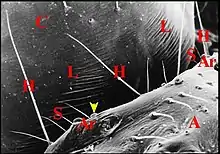
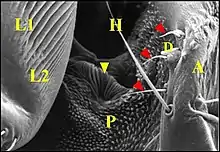
Cet appareil est pair, symétrique et surplombe les parties latéro-dorsales du pédicule où sont visibles les deux organes lyriformes (Fig.13, 14, 16). Ici comme ailleurs, il comporte bien un archet ("peigne") et une lyre chez toutes les espèces d’Argyrodes étudiées, tant au M.E.B que par l’histologie.
La lyre
Située sur la partie postéro-latérale du céphalothorax ou prosoma, elle se compose d'une série de rides ou crêtes subparallèles, à peu prés équidistantes, droites ou légèrement incurvées, séparés par des sillons, simples ou fourchues, parfois anastomosées par des "ponts", un peu inclinées vers l'axe médian du prosoma et se perdant, en périphérie, dans la microsculpture tégumentaire (Fig.13 à 16).
Les crêtes peuvent être les seuls reliefs tégumentaires de la lyre ou s'y associent avec de petites tubérosités (Argyrodes ululans).
Elles sont identiques les unes aux autres chez un même individu ou s'accentuent d'avant en arrière et de haut en bas.
Dans le cas particulier du mâle d'Argyrodes dracus (Fig. 16), elles forment deux groupes : l'un antéro-supérieur, qui en comporte une quinzaine, épaisses, très saillantes, espacées, et l'autre postéro-inférieur où elles sont beaucoup plus nombreuses, fines et très serrée. Il ne s'agit pas là d'un artéfact car cette bipartition a été observée chez plusieurs individus différents et confirmée ensuite par l'étude histologique
Il semblerait que les crêtes des femelles soient généralement moins saillantes et moins régulières que celles des mâles.
L'archet
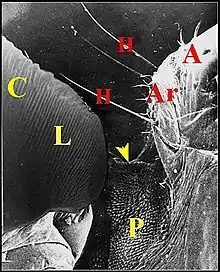
Il se compose de plusieurs unités indépendantes qui s'échelonnent en file oblique et un peu arquée (Fig.13,16 à 18) ; elles y sont bien séparées les unes des autres, excepté dans sa partie supérieure où peut apparaître un couple isolé (Fig.16). Les axes de l'archet et de la lyre ne semblent pas être parallèles ; ils délimitent un angle aigu , à sinus supéro-interne (Fig.15 à 17). Chez A. argyrodes et A.fissifrontella, il y a généralement 5 ou 6 unités par archet (Fig.13,16). Nous en avons compté moins chez A. elevatus et dracus (Fig.14,16).
Ces unités ont un même aspect fondamental : elles montrent un poil, ou soie, et son alvéole dont une partie de la margelle forme une dent excentrique (Fig.16 à 19).
Le poil est long en moyenne de 50 µm chez A.argyrodes (Fig.18,19). Il s'effile progressivement depuis sa base, où l'on ne note pas de constriction, jusqu'au sommet, très aigu et dépourvu de pore.

Orientant cette pointe vers la lyre, il a une surface lisse et présente généralement une courbure à concavité postéro-inférieure. L'alvéole (largeur totale 15 µm chez A.argyrodes) est une cupule plus ou moins profonde et d'orientation antéro-inféro-externe. Elle montre un fond "articulaire" et une margelle plus ou moins saillante. Cette margelle est toujours épaissie en ergot dans sa partie supéro-interne où elle constitue la dent ainsi excentrée : saillie acuminée triangulaire chez A.argyrodes (Fig.18,19) ou A.sp, espèce indéterminée (Fig.21), "biseau" oblique d' A.cognatus, relief globuleux d' A.fissifrontella (Fig. 20). Variable d'une espèce à l'autre, la dent l'est aussi chez un même individu où elle tend parfois à s'émousser du haut en bas de l' archet (A. cognatus). Toujours lisse sur son sommet, elle peut présenter des rides basales (Fig.19) et est dépourvue de tout pore excréteur. Elle a enfin une orientation constante et se situe toujours au-dessus et
en arrière du poil, ce qui pourrait refléter un plan de symétrie bilatérale dans l' organe sous-jacent.
La lyre
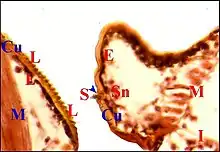
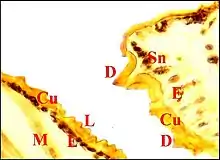
Les coupes histologiques sériées (Fig.21 à 23) montrent que la lyre est un épaississement chitineux d'aspect rigide, formé surtout d'exocuticule. Sa face profonde est lisse, régulière et repose sur un épiderme banal, uniforme, plus ou moins pigmenté, qui ne la pénètre pas. Cet épithélium entre lui-même en rapport avec un petit sinus hémolymphatique et des fibres musculaires striées. La face superficielle est en revanche régulièrement crénelée par la série des crêtes et ne présente aucune solution de continuité, la cuticule étant ininterrompue dans toute la lyre (Fig.21 à 23). Cette dernière est donc bien un simple épaississement cuticulaire périodique, sans participation de l'épiderme sous-jacent.
L'archet
Il est formé par un ensemble d'unités plus ou moins espacées (Fig.21 à 23). Au niveau de chacune d'elles, la cuticule est non seulement épaissie en bloc mais forme aussi à sa surface chaque dent observée au M.E.B. Cette saillie est plus ou moins effilée et régulière ; elle flanque le poil ou soie, qui s'y superpose dans une même coupe histologique (Fig.21) ou est retrouvé, plus dégagé et donc plus apparent, dans la section voisine.
L'épiderme sous-jacent est plus ou moins chargé de mélanine et présente un aspect banal entre les unités de l'archet. Au niveau de chacune d'elles, il se différencie en un petit massif cunéiforme s'engageant dans la cuticule jusqu'à l'alvéole du poil (Fig.21 à 23). Dans ce massif, qui correspond à l'axe sensoriel de la M.E.T, on peut discerner une cavité mal définie, deux cellules nerveuses à gros noyau nettement nucléolé et cytoplasme clair, leurs prolongements qui s'effilent vers la dent et le poil, ainsi que des cellules "satellites" plus petites et plus chromatiques. Leur ensemble entre en rapport avec des hémocytes, des fibres musculaires striées et les diverticules intestinaux de la partie antéro-supérieure de l'abdomen (Fig.23).
Ultrastructure
Alors que la lyre est continue, l'archet se compose de plusieurs unités bien distinctes qui sont indiscutablement sensorielles . Des sensilles cuticulaires d'ultrastructure très voisine se juxtaposent en revanche dans le pédicule adjacent pour y former un organe lyriforme unique.
La lyre
Elle comporte vers l'extérieur une épicuticule régulièrement ondulée, mince (0,4 nm) et présentant deux couches : l'une externe, très dense aux électrons ; l'autre interne, moins dense, moins homogène et d'épaisseur plus réduite. À leur zone de contact, ces couches forment de fines denticulations qui s'interpénètrent et les engrènent étroitement l'une dans l'autre.
L'endocuticule est épaisse d'environ 10 nm et suit les ondulations de l'épicuticule, atténuées toutefois sur sa face interne. Elle présente des canaux poraires étroits dans lesquels l'épiderme sous-jacent, uniformément plat, ne s'engage pas.
L'archet
Toutes les unités qui le constituent possèdent une même ultrastructure confirmant son aspect histologique d'organe nerveux et évoquant d'emblée des sensilles mécanoréceptrices du type I défini et décrit par Mc Iver (1975) chez les Arthropodes.
Chacune d'elles comporte un capteur externe cuticulaire et un axe sensoriel. Ce dernier est formé par 2 neurones bipolaires
et par 3 cellules enveloppes entourant les dendrites.
Capteur externe
Il est formé par le poil, son alvéole et l’excroissance en dent qu’elle porte (Fig.24).
Le poil est lisse, de section circulaire et creusé d’un canal. Ce dernier ne renferme pas de prolongements nerveux et ne communique pas avec l’extérieur. Sa paroi est épaisse, formée par de l’épicuticule et surtout , par de l’exocuticule se poursuivant, à la base, dans la membrane articulaire qui constitue le fond de l’alvéole.
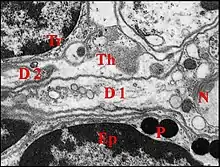
La dent est un épaississement considérable de la margelle dans sa partie antérieure, entièrement chitineux, ayant la forme d’un cône grossier, haut d’environ 10 µm et dépourvu de cavité. Son endocuticule lamellaire émet un apodème, expansion profonde dont l’aspect évoque un " levier " vertical, légèrement incurvé (Fig.23, 27,28). Cet apodème fait saillie dans une cavité qui excave la cuticule sous la membrane alvéolaire, entre la base de la dent et la zone d’implantation du poil, très excentrée.
Axe sensoriel

Comme l’histologie permettait de l’envisager, il est bien constitué par deux neurones. Bipolaire, chacun montre un axone, un corps cellulaire et un dendrite se terminant par un cil.
► L' axone est riche en mitochondries et neurotubules mais ne peut être observé sur un long trajet. Avec son homologue, il rejoint peut être le nerf du pédicule (nerf segmentaire I de Legendre, 1958).
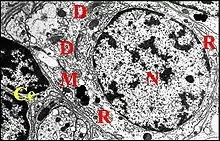
► Le corps cellulaire est ovoïde, assez volumineux et mesure approximativement 15 µm dans son grand axe, 10 dans le petit. Le noyau est plus ou moins arrondi, régulier et à chromatine peu dense, répartie en mottes, les plus petites s’accolant à l’enveloppe nucléaire. Le cytoplasme renferme du réticulum endoplasmique granulaire ou rugueux formant des saccules plus ou moins enroulés, des dictyosomes et de nombreuses mitochondries (Fig.25).
► Deux dendrites, un pour chaque neurone, pénètrent latéralement dans la cellule thécogène, d’abord séparés puis contigus (Fig.26). Leur segment interne (diamètre moyen : 1 µm) renferme des neurotubules, de nombreuses mitochondries, des ribosomes, des corps multivésiculaires et du glycogène. Chacun d’eux donne naissance à un cil à la base duquel se trouve un seul centriole d’où partent des racines ciliaires très courtes. Les microtubules y sont répartis en 9 doublets périphériques. Les cils (Fig.27,28) ont d’abord le même diamètre à leur base (0,4 µm). L’un d’eux devient ensuite plus gros que l’autre puis s’élargit brusquement à son extrémité distale où se différencie un corps tubulaire volumineux (Fig.24, 29 à 31). Ce dernier, long d’environ 15 µm pour un diamètre de 5, renferme de nombreux microtubules associés à un matériel peu osmiophile les unissant entre eux et à la membrane du cil. Il pénètre dans la cavité sous-cuticulaire, y entre latéralement en contact étroit avec l’apodème (Fig.24,29,30) et gagne la base du poil dans sa partie la plus proche de la dent chitineuse.L’autre cil diminue progressivement de diamètre et disparaît au niveau de la base du corps tubulaire.Libres à leur origine sur un trajet très court (1 à 2 µm), les deux



cils sont ensuite enfermés dans une gaine cuticulaire ou scolopale(24,27,29). Elle les enserre d’abord étroitement, au niveau des cellules thécogène et trichogène (partie inférieure), puis forme une cavité spacieuse, remarquable par les sinuosités de ses contours, jusqu’à la base du corps tubulaire . Au niveau de ce dernier, elle se rapproche de la membrane du cil et n’en est plus séparée que par une couche de gros granules (29 à 31). Ils mesurent 150 à 200 nm de diamètre et selon l’incidence des coupes, paraissent libres ou unis par un pédicule à la face interne de la gaine qui paraît alors les bourgeonner. Ils sont hétérogènes, montrent un cortex à deux couches, l’une dense, l’autre claire, ainsi qu’une zone centrale d’aspect réticulé (Fig.31) et refoulent la membrane du cil sans qu’elle se plaque toutefois sur les microtubules sous-jacents. Au-delà du corps tubulaire, la gaine ne contient plus de prolongement nerveux mais s'infléchit, s'étire en cône et va finalement s'insérer sur la base du poil en s'y confondant Fig.24).
Cellules enveloppes
Etroitement accolées, elles sont au nombre de 3 par unité d’archet : cellules thécogène, la plus interne, trichogène et tormogène, la plus externe.Elles sont garnies de microvilli apicales en bordure des cavités extracellulaires (Fig.27)
La cellule thécogène (Fig.26 à 28) est très allongée et entoure les dendrites dès leur apparition. Son noyau est basal, ovoïde et riche en chromatine formant de grosses mottes denses. Très riche en ribosomes libres, son cytoplasme renferme des dictyosomes et de fins granules probablement à l'origine de la gaine scolopale. Cette dernière se matérialise dans la cavité extracellulaire où est également visible un matériel subhomogène, dense aux électrons.
La cellule trichogène (Fig.26 à 28)se referme sur elle-même par un méso en formant un manchon. Sa partie inférieure enserre étroitement la cellule thécogène, tandis que sa partie supérieure présente une cavité extracellulaire que bordent quelques microvillosités. Le noyau est basal, très allongé et riche en chromatine. Moyennement dense aux électrons, le cytoplasme renferme de nombreuses mitochondries, des grains de sécrétion, de nombreuses vésicules de réticulum lisse et des vacuoles à contours irréguliers avec des parois plus ou moins densifiées et un contenu amorphe. Volumineuse et aplatie, la cellule tormogène fait suite à la précédente. Sa cavité extracellulaire est très spacieuse, emplie d'une substance finement grenue, en continuité avec la sous-cuticulaire, prolonge celle de la cellule trichogène et est bordée par de nombreuses microvilli (Fig.28,30).Ces dernières sont irrégulières et prennent souvent l'aspect de lamelles émettant des prolongements anastomosés. Le noyau est latéral, aplati, riche en chromatine. Le cytoplasme contient un grand nombre de mitochondries et de vacuoles.

Commentaires
Sur le plan anatomique
L' appareil stridulatoire d’Holocnemus pluchei correspond au sous-type " d II " (" organe de Simon ") individualisé par Legendre (1963) [22]Présent dans les deux sexes, il diffère donc de celui des Pholcidae américains Physocyclus, propre aux mâles et dont l’archet est un " éperon " aigu siégeant sur la base du fémur pédipalpaire (Gertsch, 1982)[23].
En ce qui concerne les Gasteracanthinae les appareils étudiés se rattachent bien au type " g ". Comme Hinton et Wilson (1970)[20] l’ont suggéré à propos des Micrathena schreibersi et gracilis, il existe bien des différences intergénériques et interspécifiques dans la microsculpture de la plaque pulmonaire. Mais outre l’espacement des crêtes, elles semblent intéresser aussi leur structure même ce que devrait confirmer une révision exhaustive.
Chez les Argyrodes, la lyre est une simple différenciation cuticulaire prosomatique, d’aspect passif et inerte, non associée avec des cellules nerveuses. L’archet en revanche est un ensemble de sensilles, chacune de ces unités, toutes semblables, appartenant au sous-groupe des mécanorécepteurs cuticulaires dans le type I de Mc Iver (1975). Le corps tubulaire en est éminemment caractéristique comme chez tous les autres Arthropodes.
Ces mécanorécepteurs sont " purs ", l’ultrastructure de leur poil, dépourvu de tout pore apical ou autre ouverture externe, et de terminaison nerveuse intra-luminale, permettant d’éliminer une composante chémoréceptrice.comme celle décrite par exemple dans les sensilles appendiculaires de Ciniflo (Dictynidae : Harris & Mill, 1973). Ils peuvent donc être toujours unique (rapprochés de ceux, également " purs ", décrits dans les tarses d’autres Araignées (genre Araneus : Chu-Wang & Foelix, 1973) et retrouvés plus tard dans tous les poils tactiles. Leurs corps tubulaires ont une ultrastructure spéciale très voisine. Tous renferment en effet de curieux granules logés entre la gaine scolopale et la membrane dendritique, granules pouvant
être d’origine cuticulaire. Observés chez des Araignées de familles différentes (Ctenidae, Dictynidae)(Barth, 1971 ; Harris & Mill, 1977), ainsi d’ailleurs que chez les Scorpions et les Tiques (Acariens), ces granules pourraient dériver de la cuticule, d' où leur autre nom de "globules cuticulaires".
L’innervation de chaque unité d’archet est multiple, comme dans tous les mécanorécepteurs d’ Arachnides, et la distingue
donc des sensilles mécanorécepteurs d’ Insectes à neurone toujours unique (Mac Iver, 1975). Mais elle est également double, ce qui l’oppose aussi aux poils tactiles banaux d’ Araignées toujours pourvus de 3 neurones que leurs terminaisons dendritiques rattachent à la base pilaire (Chu-Wang & Foelix, 1973 ; Harris & Mill, 1977), ainsi qu’aux trichobothries qui peuvent en posséder (Christian, 1971). Cette double innervation de l’unité d’archet rappelle en revanche celle des lyrifissures, isolées ou réunies en organes lyriformes, sur les appendices de Cupiennius (Barth, 1971) et surtout, en ce qui nous concerne ici, sur le pédicule des Argyrodes où des organes lyriformes en " bourrelet " (Lopez, 1996) s’associent aux appareils "stridulatoires" pour former un " complexe " au moins anatomique.
Les rapports étroits entre le corps tubulaire et l’ apodème du bord de l’alvéole rappellent ceux qui ont été signalés par Hawke et al. (1973) dans l’ ovipositeur d’un Insecte Hyménoptère, Orgilus lepidus. Une formation évoquant la structure en "levier" qu’émet l’apodème d’ Argyrodes a été décrite par Gaffal et Hansen (1972) dans les sensilles antennaires d’un autre Insecte, l’Hémiptère Dysdercus intermedius. Elle se détache toutefois du poil lui-même et en transmet vraisemblablement au dendrite
la pression mécanique.
Les deux appareils stridulatoires " d " et " g "
Ils ne peuvent intervenir dans l’équilibration.
Lorsque celui d’Holocnemus pluchei entre en activité, son composant trochantérien doit être animé d’un mouvement oscillatoire, uni ou bilatéral, qui lui fait frotter les crêtes de la lyre. Les vibrations émises chez le Pholcide sont vraisemblablement impliquées dans la reconnaissance intra-spécifique. Ce rôle paraît beaucoup plus plausible qu’une fonction d’avertissement, donc de défense, vis à vis des agresseurs éventuels, et surtout, d’une attraction sexuelle car l’appareil est présent chez le mâle, la femelle ainsi que les immatures.
Le frottement des soies fémorales de Gastéracanthines contre les crêtes pulmonaires est générateur de vibrations.
Amplifiées par le poumon jouant ainsi un rôle de caisse de résonance, elles ont pu être enregistrées en laboratoire (Hinton & Wilson, 1970 ; Uetz & Stratton, 1982), sont perçues aussi au toucher lorsqu’on saisit une Micrathena avec les doigts et peuvent être audibles comme un bourdonnement, lorsqu’on l’importune sur sa toile. Ce bruit (" low-pitched buzz ") serait perceptible jusqu’à 1 m de Micrathena gracilis (Hinton & Wilson, 1970 ; Uetz & Stratton, 1982). Levi (1985) n’a pu toutefois confirmer son émission chez plusieurs autres espèces.
Quoi qu’il en soit, l’appareil stridulatoire des Gastéracanthines n’a pas de fonction sexuelle pour les précédents auteurs, mais représente, par ses vibrations, un système défensif :
►Effet de " surprise " temporaire que l’ Araignée met à profit pour s’enfuir, se comportant ainsi comme certains Insectes stridulants (Masters, 1979)
►Imitation, par la fréquence sonore, des battements alaires de certaines Guêpes chasseresses que redoutent les autres
Arthropodes, cette sorte de "mimétisme vibratoire" protégeant ainsi l’ Araignée.
► Diffraction d’une lumière rasante par les lyres pulmonaires, du moins dans le cas de Micrathena schreibersi (Hinton & Wilson, 1970), le spectre émis jouant un rôle de couleurs d’avertissement comme dans le cas de l’appareil stridulatoire abdominal des Mutillidae (Insectes Hyménoptères) (Hinton, Gibbs & Silberglied, 1969). Complémentaires ou pas de la stridulation, ces couleurs d’avertissement "intimideraient" l’adversaire des Micrathena en renforçant l’effet aposématique de leur livrée abdominale brillante.
L’appareil stridulatoire " a "
Cet ensemble fonctionnel des Argyrodes est, sans aucun doute, impliqué dans l'équilibration.
Comme chez certains Insectes, l’apodème de l’unité d’archet et son "levier" pourraient transmettre au corps tubulaire d’ Argyrodes les vibrations ou déplacements de la dent, et la gaine dendritique, les mouvements et l’incurvation plus ou moins marquée du poil sous la poussée de la lyre.
Il est vraisemblable que la transduction du stimulus mécanique en potentiel de récepteur se produit au niveau du corps tubulaire comme dans les autres sensilles d’Arthropodes (Mc Iver, 1975)[24]. Ce corps serait comprimé (Thurm, 1964)[25] ou subirait une incurvation tendant la membrane dendritique contre le cytosquelette des microtubules. Il en résulterait une dépolarisation (Rice & al., 1973)[26].

Quant aux granules ou globules cuticulaires, ils jouent peut-être un rôle de "couplage" ou d’ "accrochage" (Christian, 1971 : "clamping device")
Les cellules enveloppes doivent sécrèter le liquide qui emplit les espaces extracellulaires ("inner and outer receptor lymph cavities ") et y assure pour les dendrites une fonction nutritive ou facilitant leurs activités électriques. De plus, l’une d’elles, la cellule thécogène, est productrice de la gaine scolopale.
En ce qui concerne la fonction réelle de l’appareil dit "stridulatoire", une émission sonore paraît très peu probable chez les Argyrodes. Bien qu’ elle soit suggérée par les noms des deux espèces Argyrodes ululans et stridulator, on ne l'a jamais été démontrée dans le genre .La biologie des Argyrodes est aujourd’hui bien connue sous l’angle des mœurs sexuelles liées en partie à la glande acronale, kleptoparasitaires et prédatrices, au cours desquelles une intervention sonore, audible ou du moins enregistrable, n’a jamais été constatée pas plus d’ailleurs que dans la grande majorité des autres appareils. Il s’ensuit que la prédiction de Legendre (1963) les créditant d’un " rôle beaucoup plus important que celui que nous tentons de (leur)attribuer " est aujourd’hui confirmée.

Une comparaison de la lyre et de l’archet des Argyrodes avec certains champs sensoriels particuliers existant chez les Insectes permet d’en fournir une interprétation fonctionnelle.
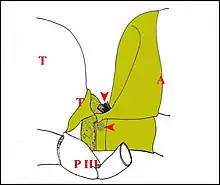
Par leur structure et surtout leur situation dans les deux parties du corps (tagmes : céphalothorax et abdomen) indépendantes, bien que très voisines, et surtout, mobiles l'une par rapport à l'autre lors des déplacements de l' Araignée, la lyre et l’archet peuvent être rapprochés de certains "champs de soies" existant chez les Hyménoptères sociaux. Parmi ces plages sensorielles pilifères décrites en détail chez l ‘Abeille, celle dite de "l’articulation du pétiole" (Lindauer & Nedel, 1959 ; Markl, 1968)[27],[28] rappelle tout particulièrement le prétendu appareil stridulatoire d’ Argyrodes. Il s’agit de deux champs de soies, pairs et symétriques, situés sur le deuxième segment abdominal et que des prolongements et replis thoraco-abdominaux "antagonistes ", les " butoirs " (Schéma), viennent heurter et tendent à courber plus ou moins lorsque l’Abeille se déplace.

Les sensillae trichodea de ces soies, innervées aussi par des cellules bipolaires, ont une ultrastructure (Thurm, 1964)[25] rappelant celle de chaque unité d’archet d’ Argyrodes, si toutefois l’on excepte leur innervation unique, et sont indiscutablement sensibles à la pesanteur comme l’ont montré diverses recherches expérimentales (in Markl., 1968). Par rapport à l’ensemble du corps, elles percevraient l’emplacement de parties toutes entières, mobiles suivant la gravité, et permettraient par voie réflexe, un contrôle musculaire de la position des axes corporels les uns par rapport aux autres.
L’existence de tels mécanorécepteurs paraît conditionnée par celle d’un abdomen volumineux, susceptible de déséquilibrer l’animal à tout instant : "gastre" des Abeilles et Fourmis mais aussi et surtout, abdomen ou opisthosoma des Argyrodes. Ce dernier, que pare souvent un vif éclat métallique (guaninocytes intestinaux), est en effet remarquablement volumineux et disproportionné par rapport au reste du corps (galerie photographique), plus ou moins élevé, conique, pesant et doit donc être sollicité sans cesse par les forces de gravité.
De ce fait, la petite Araignée est exposée à une distorsion du pédicule menaçant les organes fragiles qui le traversent lorsque son abdomen vient à s’incliner latéralement ou bascule contre le céphalothorax. À l’instar des Hyménoptères sociaux, où les sensilles abdominales sont excitées par le thorax, on peut admettre chez Argyrodes que la stimulation des sensilles d’archet (opisthosoma) par la lyre (prosoma) entraine des contractions musculaires suivant un arc réflexe et rétablit ainsi son équilibre : l’opisthosoma oriente en l’air sa face ventrale dans une position de "confort " optimum.
Cette stimulation pourrait être uni ou bilatérale, d’intensité plus ou moins grande suivant le degré d’incurvation des poils et le nombre des sensilles affectées, que la dent joue un simple rôle de butée, limitant la course du poil dans un plan donné, ou qu’elle heurte aussi les stries de la lyre. Dans ce cas, le maintien à un niveau constant du degré d’excitation de l’appareil par le jeu équilibré des poils et des dents deviendrait concevable comme chez les Insectes Hyménoptères (Markl, 1968)[28].
L’ultrastructure de l’appareil "stridulatoire" des Argyrodes et sa comparaison avec les champs sensoriels articulaires d’Insectes plaident donc en faveur d’un rôle dans la statoréception.
Il est possible que du moins chez Argyrodes, le complexe pédiculaire formé avec les organes lyriformes en "bourrelet" ne se réduise pas à de simples rapports anatomiques de voisinage mais soit une véritable association fonctionnelle entre récepteurs de gravité. Coordonnant leur activité suivant un processus qui nous échappe encore, ils permettraient à l’Araignée d’orienter dans l’espace les deux tagmes de son corps, sans préjudice pour le pédicule fragile et en fonction de sa position de repos habituelle.
Un tel concept du type " a " sera peut être étendu ultérieurement à d’autres appareils présumés "stridulatoires", les vibrations sonores, si tant est qu’elles existent réellement, se présentant alors comme un simple épiphénomène.
Notes et références
- (en) Wood-Mason, J., « On the gigantic stridulating spider. », Annals and Magazine of Natural History 16:96.,
- (en) Rovner, J. S., « Vibration in Heteropoda venatoria (Sparassidae) – a 3rd method of sound production in spiders. », J. Arachnol. 8, p.193-200.,
- (en) Uetz, G.W. & G.E.Stratton, « Acoustic communication and reproductive isolation in Spiders », Witt & Rovner ed. : Spider Communication. Mechanisms and Ecological Significance. Princeton Univ. Press, p.123-159.,
- (en) Busnel,R.G., « Acoustic Behaviour of Animals. », Published by Elsevier Publishing Company, Amsterdam, 1963, 933 pp.,
- (de) Meyer, E., « Neue sinnesbiologische Beobachtungen an Spinnen. », Z. Morph. u.Okol. Tiere 12, p.1–69 .,
- Lopez,A., « Sur deux « Tarentules » guyanaises et les poils urticants de l’une d’elles : Theraphosa leblondi (Lat.)(Araneae : Mygalomorphae : Theraphosidae). », Bull. Soc.Et.Sci.nat. Béziers, 12, 53, p.9-20.,
- Gwinner-Hanke, H., « Le comportement de deux araignées stridulantes (Steatoda bipunctata L. et Teutana grossa Koch, Theridiidae, Araneae), avec une référence particulière au comportement reproducteur. », Zeitschrift für Tierpsychologie, 27 (6), 649–678. ,
- LAWRENCE , RF, « Une collection d'Arachnida de Zululand. », Ann . Natal Mus .8 (2), p. 211-273 .,
- Lopez,A., « Araignées nouvelles ou peu connues de la Guyane française. I : le genre Argyrodes (Theridiidae). », Bull.Soc.Sciences nat., 59, p. 15-22., 1988 a.
- Lopez,A., « Les appareils stridulatoires d’Argyrodes dracus Chamb. & Iv. (Theridiidae), d’Holocnemus pluchei (Scop.)(Pholcidae) et autres Araignées. », Bull. Soc. Et. Sci.nat. Béziers, 12, 53, p. 21-31., 1988 b.
- Lopez,A.avec L.Juberthie-Jupeau., « Les organes sensoriels sus-pédiculaires d'Araignées : un complexe présumé stato-récepteur. », Bull.Soc.Et.Sci.nat.Béziers,N.S., XIV(55),1993-1994, p.32-37., 1994a
- Lopez,A., avec L. Juberthie-Jupeau, « L’appareil « stridulatoire » des Argyrodes (Araneae : Theridiidae) : un complexe sensoriel présumé stato-récepteur. », Mém.Biospéol.,XXI, p.91-96., 1994b
- (en) Rovner,J.S., « Sound production by Nearctic wolf spiders – Substratum-coupled stridulatory mechanism. », Science,190, p.1309-1310.,
- (en) Forster & Platnick, « A review of the archaeid spiders and their relatives, with notes on the limits of the superfamily Palpimanoidea (Arachnida, Araneae). », Bulletin of the American Museum of Natural History, no 178, p. 1-106,
- Legendre,R, « Un organe stridulant nouveau chez les Archaeidae (Araneida). », Bull.Soc.Zool.France, 95 (n°1), p.29-30.,
- Lopez, A., « Araignées de Guyane française : II. La sous-famille des Gasteracanthinae O.Pickard-Cambridge (Araneidae) et les proies de Sceliphron fistularium (Dahlbom)(Hymenoptera : Sphecidae). », Bull.Soc.Sciences nat., 82, p. 14-22., 1994c
- Lopez & Marcou, « Holocnemus pluchei », site garrigue languedocienne Internet, (: http://faune-flore-languedocienne.alwaysdata.net/garrigue/Araignees_garrigue.html#Holocnemus_pluchei [archive])
- Huber,B.A., « Mécanisme copulateur chez Holocnemus pluchei et Pholcus opilionoides, avec des notes sur les apophyses chélicérales mâles et les organes stridulateurs chez les Pholcidae (Araneae). », Acta Zoologica 76 (4): 291 - 300.,
- Lopez,A. avec R.Legendre, « Les chromatophores de l’Araignée Holocnemus pluchei (Scop.)(Pholcidae) (Note préliminaire). », Bull. Soc.zool.France, 98 (4), p.487-494.,
- (en) Hinton, H.E. & R.S.Wilson, « Stridulatory organs in spiny orb‐weaver spiders. », Journal of Zoology 162 (4): 481 - 484 ·,
- (en) Levi, H., « The spiny orb-weaver genera Micrathena and Chaetacis (Araneae: Araneidae). », Bulletin of the Museum of Comparative Zoology, vol. 150, no 8, p. 429-618,
- Legendre , R ., « L'audition et l'émission de sons chez les Aranéides. . », Ann. Biol. 2 , 371-390,
- (en) Gertsch, W. J., « The spider genera Pholcophora and Anopsicus (Araneae, Pholcidae) in North America, Central America and the West Indies. », Assoc. Mex. Cave Stud. Bull., 8, pp. 95-144.,
- (en) Mac Iver, S.B., « Structure of cuticular mechanoreceptors of Arthropods. », Ann.Rev.Ent.,20, p.381-397.,
- (en) Thurm,U., « Mecanoreceptors in the cuticle of the honey-bee structure and stimulus mechanism. », Science, N.Y. 145, p.1063–1065,
- (en) Rice,M.J.,Galun,R. & L.H.Finlayson, « Mechanotransduction in Insect Neurones. », Nature New.Biol.,241, p.286-288.,
- (de) Lindauer, M. & J.O.Nedel, « Ein Schweresinnesorgan der Honigbiene. », Z.vergl.Physiol.,42, p.334-364.,
- Markl,H., « Conservation de l'équilibre et sens de la pesanteur chez l'abeille », in "Traité de la Biologie de l' Abeille" ,II, p.146-172.,
Bibliographie
- Berland, L., 1932.- Les Arachnides. Biologie, Systématique. Encycl.Entomol., XVI. Paul Lechevalier & fils edit., Paris, 486 pp.
- Busnel, R.G., 1963.- Acoustic Behaviour of Animals. Published by Elsevier Publishing Company, Amsterdam, 1963, 933 pp.
- Legendre , R ., 1963 - L'audition et l'émission de sons chez les Aranéides. Ann. Biol. 2 , 371-390.
- Lopez, A. avec R.Legendre, 1973 – Les chromatophores de l’Araignée Holocnemus pluchei (Scop.)(Pholcidae) (Note préliminaire). Bull. Soc.zool.France, 98 (4), p.487-494.
- Lopez, A., 1988 b. – Araignées nouvelles ou peu connues de la Guyane française. I : le genre Argyrodes (Theridiidae). Bull.Soc.Sciences nat., 59, p. 15-22.
- Lopez, A., 1988 c. – Les appareils stridulatoires d’Argyrodes dracus Chamb. & Iv. (Theridiidae), d’Holocnemus pluchei (Scop.)(Pholcidae) et autres Araignées. Bull. Soc. Et. Sci.nat. Béziers, 12, 53, p. 21-31.
- Lopez, A., avec L.Juberthie-Jupeau, 1994a - Les organes sensoriels sus-pédiculaires d'Araignées : un complexe présumé stato-récepteur. Bull.Soc.Et.Sci.nat.Béziers, N.S., XIV(55), 1993-1994, p.32-37.
- Lopez, A., avec L. Juberthie-Jupeau, 1994b - L’appareil « stridulatoire » des Argyrodes (Araneae : Theridiidae) : un complexe sensoriel présumé stato-récepteur. Mém.Biospéol., XXI, p.91-96.
- Lopez, A., 1994c. – Araignées de Guyane française : La sous-famille des Gasteracanthinae O.Pickard-Cambridge (Araneidae) et les proies de Sceliphron fistularium (Dahlbom)(Hymenoptera : Sphecidae). Bull.Soc.Sciences nat., 82, p. 14-22.
- Lopez, A., avec L. Juberthie-Jupeau, 1996 . – Les organes lyriformes du pédicule des Araignées : observations microscopiques chez Argyrodes argyrodes (Walck.) (Theridiidae) et chez deux Metinae souterraines : Meta bourneti Sim. et Meta menardi (Latr.) (Araneidae). Mém. Biospéol., 23, p. 157-162.
- Simon, E– Anns.soc.ent.France, 62, p.224-225.
- Uetz, G.W. & G.E.Stratton, 1982.- Acoustic communication and reproductive isolation in Spiders in Witt & Rovner ed. : Spider Communication. Mechanisms and Ecological Significance. Princeton Univ. Press, p.123-159.
- Holocnemus pluchei Lopez & Marcou : http://faune-flore-languedocienne.alwaysdata.net/garrigue/Araignees_garrigue.html#Holocnemus_pluchei
 Portail de l’arachnologie
Portail de l’arachnologie