Cytoplasme
Le cytoplasme désigne le contenu d'une cellule vivante et la région comprise entre la membrane plasmique et le noyau d'une cellule eucaryote ou le nucléoïde d'une cellule procaryote[1]. En d'autres termes, le protoplasme délimité par la membrane plasmique[2],[3] est constitué du cytoplasme et du noyau chez les cellules eucaryotes, du cytoplasme et du nucléoïde pour les cellules procaryotes. Il est constitué d'environ 80 % d'eau[4] et se définit comme une émulsion colloïdale à l'aspect granuleux constituée de cytosol, solution aqueuse de sels minéraux et de divers composés organiques qui, avec le cytosquelette, forme le hyaloplasme. Chez les eucaryotes, le cytoplasme comprend plusieurs organites comme les réticulum endoplasmiques, des vacuoles, des mitochondries ou des chloroplastes formant le morphoplasme, mais ne comprend pas le noyau (dont le contenu est appelé nucléoplasme). Il comprend également des ribosomes, qui ne sont pas considérés comme des organites. Chez les procaryotes, le cytoplasme englobe la totalité du matériel cellulaire contenu dans la membrane interne (ou cytoplasmique). Il ne contient pas d'organites mais des ribosomes, des substances de réserves, des plasmides et un chromosome circulaire.
Le cytoplasme peut se diviser et former à l'extérieur de la cellule une masse gélatineuse, appelée ectoplasme, et qui contribue au mouvement cellulaire. Il peut alors présenter à l'intérieur une zone plus fluide, appelée endoplasme, où l'on trouve la majorité des organites[5]. Le cytoplasme est un milieu riche en nutriments qui traversent la membrane plasmique et migrent vers les organites.
Chez les eucaryotes, le cytoplasme est constitué d'un milieu plus ou moins homogène, le cytosol, dans lequel baignent les organites. Le cytoplasme se compose d'un réseau de membranes, connues sous le nom de réticulum endoplasmique (lisse et rugueux) qui servent de surface de travail pour toutes les activités biochimiques. Il contient une petite partie du génome d'un organisme. Même si la plus grande partie de ce génome est contenue dans le noyau cellulaire, certains organites du cytoplasme, les mitochondries ou les chloroplastes (chez les végétaux), possèdent leur propre ADN[6],[7]. Ce génome cytoplasmique se transmet la plupart du temps par voie maternelle (c'est-à-dire par le gamète femelle).
Cytosquelette

Le cytoplasme est constitué d'un réseau de filaments protéinés qui confère à la cellule sa forme et son organisation interne et lui permet de se déplacer. Ces filaments forment le cytosquelette. On peut néanmoins rencontrer plusieurs types de filaments :
- les filaments d'actine ou microfilaments, typiques des cellules musculaires ;
- les microtubules, qui apparaissent disséminées dans le hyaloplasme ou forment des structures plus complexes, comme les fuseaux achromatiques ;
- les filaments intermédiaires, comme les filaments de kératine typiques des cellules épidermiques.
À son tour, cette structure maintient un lien entre les protéines et génère des structures plus complexes et plus stables. Ils sont également responsables du mouvement cytologique.
Cytosol
Le milieu intracellulaire se compose d'un liquide appelé hyaloplasme ou cytosol, laquelle matrice contient des organites. Beaucoup de processus biochimiques, dont la glycolyse, se déroulent dans le cytoplasme.
Dans une cellule eucaryote, le cytosol peut occuper de 50 à 80 % du volume de la cellule. Le reste du volume est occupé par des molécules formant une suspension colloïdale. En règle générale, ces molécules sont en fait des macromolécules.
Étant un liquide aqueux, le cytosol ne présente pas de forme ou de structure stable, même si, temporairement, il peut prendre deux types d'aspect :
- une consistance de gel (état gel) ;
- une consistance de fluide (état sol).
Les changements de formes du cytosol permettent à la cellule de s'adapter aux nécessités métaboliques et joue également un rôle important lors du mouvement cellulaire.
Ribosome
Les ribosomes sont des granules cytoplasmiques présents dans toutes les cellules, mesurant environ une vingtaine de nanomètres. Par ailleurs, ils sont porteurs de l'ARN ribosomique. Les ribosomes ne sont pas des organites.
La synthèse de protéines se déroule dans ces ribosomes, au sein du cytoplasme[8]. L'ARN messager (ARNm) et l'ARN de transfert (ARNt) sont synthétisés dans le noyau cellulaire avant d'être libérés dans le cytoplasme. L'ARN ribosomique (ARNr) est libéré dans le cytoplasme sous la forme d'une sous-unité ribosomique. Deux types de ces sous-unités peuvent alors se combiner avec des molécules d'ARNm pour former un ribosome actif[9].
Les ribosomes actifs peuvent alors soit rester en suspension dans le cytoplasme, soit se fixer sur le réticulum endoplasmique rugueux[10]. Ceux qui restent en suspension ont pour fonction principale de synthétiser les protéines suivantes :
- Les protéines qui forment le cytosol ;
- Les protéines qui construiront les éléments structuraux ;
- Les protéines qui composent les éléments mobiles du cytoplasme ;
- Les protéines membranaires et matricielles des peroxysomes.
Le ribosome se compose de deux parties, une sous-unité mineure et une sous-unité majeure, qui sortent chacune du noyau[11]. Des expériences ont montré qu'elles sont maintenues ensemble grâce à des charges, car lorsque l'on abaisse la concentration en ions Mg2+, les sous-unités tendent à se séparer.
Lysosome
Les lysosomes sont des vésicules sphériques dont le diamètre varie entre 0,1 et 1 micromètre[12]. Ils contiennent près de cinquante enzymes, pour la plupart hydrolysables en solution acide. Les enzymes ont en effet besoin de cette solution acide pour avoir un fonctionnement optimal[13]. Les lysosomes permettent de maintenir ces enzymes à l'écart du reste de la cellule car, dans le cas contraire, ils pourraient réagir chimiquement avec les éléments et les organites de celle-ci.
Ils utilisent leurs enzymes pour recycler les différents organites de la cellule en les absorbant, en les assimilant puis en libérant ses composantes au sein du cytosol[13]. Ce processus est connu sous le nom d'autophagie, processus au cours duquel la cellule digère ses propres structures devenues inutiles. Ces structures sont englobées par des vésicules qui proviennent du réticulum endoplasmique et de l'appareil de Golgi, formant alors un autophagosome. Si celui-ci s'unit à un lysosome primaire, ils forment alors un autophagolysosome et suit le même processus que celui présenté précédemment.
Au cours de l'endocytose, les matériaux sont rassemblés à l'extérieur de la cellule, puis recouverts par la membrane plasmique, pour former une phagosome. Le lysosome s'unit au phagosome pour former alors une phagolysosome et déverse son contenu dans le lysosome après avoir été dégradé. Une fois hydrolysées, les molécules utiles sont libérées à l'intérieur de la cellule pour rejoindre les voies métaboliques. Le reste est éliminé par exocytose.
Les lysosomes déversent également leurs enzymes vers le milieu extérieur par exocytose, pour dégrader aussi d'autres structures.
Compte tenu de leur rôle, on en trouve beaucoup à l'intérieur des globules blancs car ils permettent la dégradation des corps étrangers.
Vacuole
La vacuole est une poche de liquide entourée d'une membrane. Dans une cellule végétale, on trouve une seule vacuole de grande taille, alors que dans une cellule animale, elles sont nombreuses et de petite taille. La membrane qui l'entoure est appelée tonoplaste.
La vacuole des cellules végétales contient une solution de sels minéraux, sucres, d'acides aminés et parfois de pigments comme l'anthocyane. Elle permet non seulement d'agir comme une réserve alimentaire (de sucres et d'acides aminés), mais également de donner une pigmentation aux pétales (grâce à l'anthocyane) et de remplir la fonction des lysosomes grâce à ses enzymes.
Le rôle des vacuoles dans les cellules animales est d'agir comme un endroit où sont stockées les protéines[14]. Ces protéines sont conservées pour une utilisation ultérieure, ou plutôt pour être exportées en dehors de la cellule, par exocytose. Au cours de ce processus, les vacuoles fusionnent avec la membrane et son contenu est alors libéré à l'extérieur. La vacuole peut aussi fonctionner en endocytose. Le phénomène est alors inversé et des matériaux qui ne pouvaient pas franchir la membrane sont transportés à l'intérieur de la cellule[15].
Réticulum endoplasmique
Le réticulum endoplasmique est un système complexe de membranes reliées entre elles et qui forment un squelette cytoplasmique[16]. Il s'organise en un vaste système de canaux contenant des ribosomes. Sa forme peut varier car leur nature dépend de la disposition et de l'arrangement des cellules, qui peuvent être comprimées ou espacées.
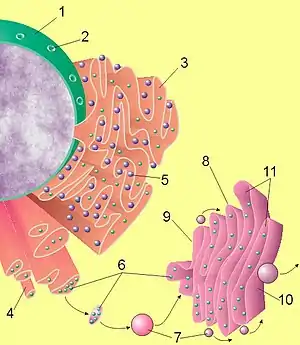
(1) Noyau.
(2) Pore nucléaire.
(3) Réticulum endoplasmique rugueux (RER).
(4) Réticulum endoplasmique lisse (REL).
(5) Ribosome du RER.
(6) Protéines transportées.
(7) Vésicule.
(8) Appareil de Golgi.
(9) Côté cis de l'appareil de Golgi.
(10) Côté trans de l'appareil de Golgi.
(11) Lumière de l'appareil de Golgi.
Il s'agit d'un ensemble de cavités fermées aux formes très variables, allant de lames aplaties, jusqu'à des vésicules, en passant par des tubes enroulés à l'aspect sinueux. Ils communiquent les uns avec les autres et forment un réseau complètement isolé du hyaloplasme grâce à la membrane du réticulum endoplasmique. Par conséquent, le liquide cytoplasmique se divise en deux parties : l'espace citernal (ou lumière) contenu à l'intérieur du réticulum endoplasmique et l'espace cytosolique à l'extérieur.
Les principales fonctions du réticulum endoplasmique sont les suivantes :
- faire circuler des substances qui ne sont pas libérées dans le cytoplasme ;
- servir d'espace pour les réactions chimiques ;
- synthétiser et transporter des protéines produites par les ribosomes fixés aux membranes (dans le RER uniquement) ;
- glycosyler les protéines (RER seulement) ;
- produire des lipides et des stéroïdes (REL seulement) ;
- fournir un squelette structurel pour maintenir la forme de la cellule.
Réticulum endoplasmique rugueux
Quand la membrane est entourée de ribosomes, on la qualifie de réticulum endoplasmique rugueux (RER)[17]. Le RER a pour fonction principale de synthétiser les protéines. Pour cette raison, il produit plus de cellule en croissance ou qu'il sécrète des enzymes[18]. De la même manière, si la cellule est abîmée, cela peut conduire à une augmentation de la synthèse protéinique pour permettre la réparation des dommages.
Les protéines sont transformées et déplacées dans une région du RER : l'appareil de Golgi.
Réticulum endoplasmique lisse
En l'absence de ribosomes, on le qualifie de réticulum endoplasmique lisse (REL). Sa fonction principale est de produire les lipides de la cellule (en pratique, les phospholipides et le cholestérol) qui forment les briques élémentaires des membranes cellulaires[13]. Le reste des lipides cellulaires (acide gras et triglycéride) sont synthétisés dans le cytosol. C'est pour cette raison qu'il est plus abondant dans les cellules de sécrétion (telle que la glande sébacée). Cela reste rare malgré tout pour la plupart des cellules.
Appareil de Golgi
L'appareil de Golgi (du nom de son découvreur, Camillo Golgi) a une structure similaire à celle du réticulum endoplasmique, mais est plus compact que ce dernier. Il se compose de sacs en forme de disques et se situe à proximité du noyau cellulaire. Chaque pile de saccules s'appelle un dictyosome[19]. Il mesure environ un micromètre de diamètre et se compose d'environ six saccules, alors que dans les cellules eucaryotes ce nombre peut atteindre la trentaine. L'extrémité de chaque dictyosome est évasée et entourée de vésicules qui ont soit fusionné avec, soit qui s'en sont séparées par gemmation[20].

L'appareil de Golgi est structurellement et biochimiquement polarisé. Il possède deux faces distinctes : le côté cis (ou de formation) et le côté trans (ou de maturation)[21]. La face cis est située à proximité des membranes du réticulum endoplasmique. Ses membranes sont fines et sa composition est similaire à celles du réticulum. Elles sont entourées par des vésicules golgiennes, aussi appelé vésicules de transition qui sont issues du réticulum. La face trans est généralement proche de la membrane du plasma. Ses membranes sont plus épaisses et similaires à celle du plasma. On trouve sur ces faces des vésicules plus grandes, les vésicules sécrétrices.
Leur rôle est de modifier les substances synthétisées dans le RER[22]. Ces transformations peuvent être une agrégation des restes de glucides pour obtenir une structure finale ou alors pour être protéolysé et ainsi acquérir une conformation active. Par exemple, dans le RER des cellules acineuses du pancréas synthétisent la pro-insuline qui acquiert la conformation finale de l'insuline grâce aux transformations qu'elle subit dans l'appareil de Golgi. Les enzymes que l'on trouve à l'intérieur des dictyosomes sont capables de modifier des macromolécules par glycosylation (ajout de glucides) et par phosphorylation (ajout de phosphates). À cette fin, l'appareil de Golgi transporte certaines substances telles que des nucléotides et des sucres dans les organites du cytoplasme. Les protéines sont également marquées par des séquences de signaux qui déterminent leur destination finale, comme le mannose-6-phosphate qui est ajouté aux protéines des lysosomes. L'appareil de Golgi produit également les glycoprotéines nécessaires à la sécrétion qui permet d'ajouter un glucide à une protéine.
En outre, l'appareil de Golgi sécrète des enzymes, telles que les enzymes digestives du pancréas. Elles traversent tous les sacs de l'appareil et quand elles arrivent au niveau de la face trans du dictyosome (sous la forme de vésicules de sécrétion), elles sont transportées vers leur destination finale, en dehors de la cellule, par exocytose. L'appareil de Golgi est le plus important des organites pour la synthèse des hydrates de carbone. Cette tâche est réalisée par des enzymes golgiennes grâce à un résidu de xylose. Parmi les autres fonctions de l'appareil de Golgi, on trouve également la séparation des glucides (comme ceux utilisés pour la réparation cellulaire), le transport et le stockage des graisses et la formation des lysosomes primitifs.
Mitochondrie

La mitochondrie est un organite qui se trouve dans toutes les cellules eucaryotes, bien qu'elles puissent être absentes des cellules très spécialisées. Leur taille est généralement de cinq micromètres de large sur 0,2 µm d'épaisseur et possède une double membrane[23]. L'espace inter-membranaire a une composition similaire à celle du hyaloplasme. Le rôle de cet espace est principalement de produire des molécules qui servent de précurseurs à la biosynthèse de macromolécules dans le hyaloplasme et d'être le lieu où se déroulent l'oxydation respiratoire et la synthèse des protéines mitochondriales. Cette dernière fonction se déroule de la même manière que la synthèse des protéines dans le hyaloplasme.
Les membranes de la mitochondrie sont constituées de phospholipides et de protéines, qui se combinent pour former un réticulum lipido-protéiné[23]. La membrane la plus externe contrôle l'entrée et la sortie de substances dans la cellule et sépare l'organite du hyaloplasme. La membrane externe contient des protéines de transport spécialisées qui permettent le passage de molécules du cytosol jusqu'à l'espace inter-membranaire[24]. Ces protéines forment de grands canaux d'eau ou de porines, qui rendent la membrane très perméable, contrairement à la membrane interne. La membrane est également constituée d'enzymes comme ceux qui activent l'oxydation des acides gras dans la matrice.
La membrane interne est repliée vers le centre de la mitochondrie ce qui donne lieu à des extensions, les crêtes mitochondriales ou encore cristae. Ceux-ci s'étendent sur toute la largeur de l'organite et présentent des granules à leur surface[24]. De facto, c'est donc la membrane qui possède la plus grande surface, en raison de ces crêtes mitochondriales. C'est également la membrane cellulaire la plus riche en protéines. On y trouve notamment : les protéines qui forment la chaîne respiratoire transportant les électrons vers l'oxygène moléculaire, un complexe enzymatique appelé ATP synthase, qui agit comme catalyseur lors de la synthèse de l'ATP, ainsi que des protéines de transport qui permettent le passage des ions et des molécules à travers la membrane mitochondriale interne. Cette membrane présente l'avantage de ne pas avoir de cholestérol entre ses lipides et d'être riche en un phospholipide rare, la cardiolipine. D'une façon générale, sa fonction principale est d'être le lieu du processus respiratoire.
La mitochondrie se divise donc en deux espaces distincts : l'espace inter-membranaire et la matrice[25]. La matrice a une consistance de gel en raison de la présence d'une concentration élevée de protéines hydrosolubles et de 50 % d'eau. Par ailleurs, elle contient des molécules d'ADN (l'ADN mitochondrial) qui contiennent l'information pour synthétiser un bon nombre de protéines mitochondriales, des molécules d'ARN mitochondrial formant les ribosomes mitochondriaux (ou mitoribosomes), des ions, du calcium, des phosphates, de l'ADP, de l'ATP, des coenzymes A et une grande quantité d'enzymes.
Finalement, la mitochondrie permet non seulement l'oxydation du pyruvate en CO2 couplée à la réduction des porteurs électroniques NAD+ et FAD (NADH et FADH2), mais également le transfert des électrons depuis le NADH et le FADH2 vers le dioxygène couplé à la génération d'une force proton-motrice, ou encore l'utilisation de l'énergie stockée dans le gradient électrochimique de protons pour la synthèse de l'ATP du complexe FO-F1.
Peroxysome

Les peroxysomes sont des microcorps à membrane sphérique, dont le diamètre est compris entre 0,15 et 1,7 micromètre. Ils se forment par gemmation à partir du réticulum endoplasmique lisse. Ces granules n'ont pas de structure interne. Ils possèdent un grand nombre d'enzymes métaboliques, en particulier l'enzyme catalase, qui catabolise la dégradation du peroxyde d'hydrogène (d'où provient le nom de peroxysome). La dégradation du peroxyde d'hydrogène est représentée par l'équation :
Ils effectuent des réactions d'oxydation qui ne produisent pas d'énergie directement utilisable par le reste de la cellule (ils ne génèrent pas d'ATP)[25]. À l'intérieur des peroxysomes est également dégradée la purine et, dans les plantes, elles sont impliquées dans la photorespiration.
Notes et références
- (es) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en espagnol intitulé « citoplasma » (voir la liste des auteurs).
- Lodish et al, Biologie moléculaire de la cellule, 4e édition pour la traduction, Paris, De Boeck, 2014, p. 13
- « citoplasma », WordReference, (consulté le )
- « Definición de citoplasma », Definición.org (consulté le )
- V. A. Shepherd, « The cytomatrix as a cooperative system of macromolecular and water networks », Current Topics in Developmental Biology, vol. 75, , p. 171–223 (ISSN 0070-2153, PMID 16984813, DOI 10.1016/S0070-2153(06)75006-2, lire en ligne, consulté le )
- « Letra E », Diccionario Ecológico, Ambiente Ecológico (ISSN 1668-3358, consulté le )
- J.S. Raisman et Ana M. González, « El genoma extranuclear », Hipertextos del Área de Biología, Facultad de Agroindustrias (consulté le )
- « Cloroplastos », Biología, Soko (consulté le )
- Guillermo Becco, « Síntesis de proteínas », Monografias.com (consulté le )
- Juan Sebastián Ramírez, « Estructura y función celular », Monografias.com (consulté le )
- « Biología », Área de Ciencias, Preuniversitario Popular Víctor Jara (consulté le )
- Juan Sebastián Ramírez, « Estructura y función celular », Ilustrados.com (consulté le )
- Julio Pérez Márquez, « Lisosoma », La Celúla, Université d'Alcalá (consulté le )
- « Lisosomas », Manual de Biología Celular, Centro Nacional de Información y Comunicación Educativa (consulté le )
- Mavel Egaña, « Célula. Morfología celular. Teoría celular », monografias.com (consulté le )
- « Estructura y funcionalidad de la membrana celular », Liceo Alemán de Santiago (consulté le )
- « Biología-1 », Diccionarios digitales, (consulté le )
- J.S. Raisman et Ana M. González, « Célula Eucariota: citoplasma », Facultad de Agroindustrias (consulté le )
- « Los orgánulos celulares », hiru.com (consulté le )
- « Aparato de Golgi », Citoplasma, tematiqos.com
- « Aparato de Golgi », Manual de Biología Celular, Centro Nacional de Información y Comunicación Educativa (consulté le )
- « Cuerpo de Golgi », Biología, Soko (consulté le )
- L'appareil de Golgi
- « Mitocondrias », Trabajos de medicina, iEspaña (consulté le )
- « Mitocondrias », Manual de Biología Celular, Centro Nacional de Información y Comunicación Educativa (consulté le )
- « Citoplasma », Elementos de Biología, Université d'Arizona (consulté le )
Voir aussi
Bibliographie
- Glenn Toole et Susan Toole, Biology for Advanced Level, Cheltenham: Stanley Thornes Publishers Ltd, , 698 p. (ISBN 0-7487-3957-2, lire en ligne)
- C J Clegg et D G Mackean, Advanced Biology : Principles and Applications, Cheltenham: Stanley Thornes Publishers Ltd, , 712 p. (ISBN 978-0-7195-7670-6)
Articles connexes
- Biologie cellulaire
- Chytosoïde
- Déterminant cytoplasmique
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire