Organisme hématophage
Les organismes hématophages, du grec hemato « sang » et -phage « manger », ou « sanguinivores », sont des organismes qui se nourrissent de sang. Ce sont souvent des ectoparasites (parasites qui n'entrent pas à l'intérieur de leur hôte, mais qui se fixent provisoirement sur sa peau).
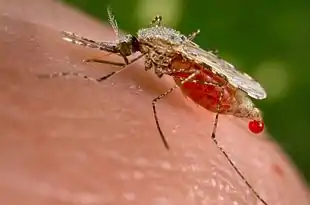




Dans l'écosystème, ils occupent des points particuliers des réseaux trophiques – les hématophages sont souvent à la fois la proie et le prédateur d'un même hôte – ce qui leur confère des caractéristiques inhabituelles dans le système prédateur-proie[1].
Exemples d'organismes hématophages
- Parmi les mammifères, il y a les chauves-souris vampires.
- Chez les oiseaux, une des espèces de pinson de Darwin sur les îles Galápagos, le Géospize à bec pointu, se nourrit de sang qu'il puise sur les fous.
- Parmi les organismes aquatiques (proches des poissons), on peut citer les lamproies.
- Chez les annélides, on peut citer certaines espèces de sangsues (dont plusieurs des espèces trouvées en France).
- Parmi les arthropodes, on peut citer divers insectes piqueur-suceurs, tels que :
- les moustiques ;
- les simulies ;
- la tribu Stomoxyini des Muscinae (mouche charbonneuse, mouche des cornes) ;
- les taons qui mordent pour arracher la chair ;
- les phlébotomes ;
- les Ceratopogonidae ;
- les puces (Siphonaptera) et les poux (Phthiraptera) ;
- les punaises hématophages (triatomes et cimicidés) ;
ou des acariens tels que :
- les tiques ;
- les poux rouges des volailles ;
- les aoûtats.
Modes de l'hématophagie chez les arthropodes
L'entomologiste M. M. J. Lavoipierre distingue deux modes essentiels de l'hématophagie[2] : la piqûre ou l'arthropode solénophage (du grec solen, tuyau, et phagein, manger) où l'animal perce la peau grâce à une trompe longue, perforante et cathétérisant les capillaires veineux (d'où le terme anglais de « capillary feeding »). Cela concerne le groupe des punaises, des moustiques et des poux. Avec la piqûre telmophage (du grec têlma marécage, et phagein manger), l'animal lacère le derme afin de créer un microhématome, une mare de sang (d'où le terme anglais de « pool feeding ») dont il aspire le contenu (exemple : moucherons de la famille des phlébotomes, taons)[3],[4].
Recherche de l'hôte et orientation spatiale
Les hématophages, grâce à un système perceptif spécialisé (olfaction, thermoperception, chémotactisme, vision nocturne…), repèrent leur hôte au moyen de différents indices tels que :
- rayonnement infrarouge (chaleur émanant du corps d'un hôte potentiel), qui suffit par exemple - à lui seul - à provoquer une réponse d’extension du proboscis, mais qui peut cependant être modulé par l’expérience (démontré chez Rhodnius prolixus par Lallement[5]) ;
- vibrations ;
- facteurs chimiques.
Les moyens pour l'hématophage de percevoir son hôte
La perception de la température et/ou de molécules émises par l’hôte se fait par des récepteurs sensoriels spécialisés :
- perception visuelle : certains hématophages comme la tique n'en sont pas dotés. Des insectes comme les punaises ont une vision suffisante pour susciter des réponses optomotrices (réflexe de stabilisation en réponse aux mouvements d'objets de l'environnement[6]) ;
- thermo-perception : les triatomes comme Triatoma infestans apprécient la température d'une source distante et sa direction[7] ;
- perception des vaisseaux sanguins : divers hématophages détectent le vaisseau au travers de la peau et/ou au fur et à mesure de la pénétration du rostre. Les nymphes de triatomine ou le moustique femelle sont ainsi capables de repérer sous la peau un vaisseau sanguin qui est à leur portée. La triatomine le fait, grâce pense-t-on à des thermorécepteurs antennaires[8] ;
- sensibilité « chimio-olfactive » permettant une perception de molécules (hormones ?) et odeurs ;
- perception du CO2 : ce gaz est un traceur émis par l'expiration. Certains moustiques et les « mouches tsé-tsé » le perçoivent[9] mais ne se montrent pas non plus piégés par d'autres sources de CO2 (compostage, combustion),
- perception de la transpiration (ou des marques laissées par elle) : les moustiques Aedes (ex. : Aedes atropalpus) sont dotés de capteurs de ce type[10],
- perception de la vapeur d'eau : associée à la température, c'est semble-t-il le facteur d'attraction qui devient prédominant quand une punaise hématophage arrive près de son hôte[6],
- anémo-perception : il existe des réponses anémotactique se traduisant par l'orientation de l'insecte vers l'origine d'un courant d'air (en l'absence d'autres stimuli)[6],
- perception de l'acide lactique : c'est le cas chez le moustique Aedes aegypti[11],
- des caractéristiques d'osmolarité/salinité[12] et chimiques confirment à l'insecte qu'il est bien en train de prélever du sang et d'entamer alors son repas[13],
- perception auditive. Ce cas semble plus rare mais une étude a montré qu'une mouche hématophage se guide vers des grenouilles via le chant des mâles[14].
Orientation dans l'espace après le repas
Une fois le repas pris, les nymphes de certains insectes nocturnes et non volants (triatomine par exemple) doivent retrouver leur refuge diurne[15] :
- Perception de phéromones « d’agrégation » et « d'arrêt » : ces phéromones d'agrégation[16],[17] sont émises notamment par leurs excréments[18],[19]. Il est alors contrôlé par son cycle diurne[20].
- Modulation régulation du comportement hématophage : les comportements de « quête » (recherche active et/ou attente à l'affut de l'hôte) peuvent être modulés par l'état physiologique des individus :
- Un individu affamé peut rechercher plus activement un hôte ;
- Un individu fraichement nourri a des réactions de quête inhibées ;
- Un individu vulnérable car venant de subir une mue voit aussi son comportement de quête inhibé, puis il reprend progressivement ;
- Un repas complet peut même induire une répulsion des insectes face à des stimuli chimiques ou thermiques autrement attractifs.
Autres récepteurs : les tiques n'ont pas de récepteurs antennaires, mais d'autres types d'organes notamment disposés sur les pattes. Elles n'ont pas de vision mais s'orientent précisément vers leurs hôtes, peut-être attirées par leur odeur, plus que par leur température car elles peuvent parasiter des serpents, lézards, orvets qui sont des animaux à sang froid.
Remarque : la sensibilité olfactive de la punaise hématophage Rhodnius prolixus varie selon l'heure de la journée (inhibée le jour, activée la nuit)[1]. Cette punaise répond en outre différemment à différents stimuli chimiques[1].
Efficience relative des récepteurs sensoriels
On estime que la précision de la perception sensorielle se traduit en grande partie par le nombre de chemorécepteurs. Et elle semble varier selon le mode de vie de l'hématophage, et plus précisément selon que l'hématophage vive en association permanente ou non avec son hôte[21].
On peut distinguer
- Les espèces généralement proches de leur hôte : ce sont par exemple les poux et certaines puces (puces « domestiques » pour l'homme). Ces organismes ne possèdent que 10 à 50 récepteurs antennaires, respectivement. La punaise des lits Cimex lectularius en possède 56, soit un tout petit peu plus que la puce ;
- Les espèces vivant plus ou moins loin de leur hôte : ce sont par exemple les moustiques Culex ou la « mouche des étables », Stomoxys calcitrans (environ 5000 récepteurs antennaires). Le triatome Triatoma infestans qui passe une partie de sa vie loin de son hôte en possède 2900[22].
Ces chemorécepteurs ne servent pas qu'à repérer l'hôte le plus proche, mais aussi au sein de l'espèce à la communication hormonale, y compris pour les besoins de la sexualité. Chez une espèce dont les mâles et femelles sont souvent éloignés, un nombre plus important de récepteurs est nécessaire. Des rythmes circadiens régulent leurs fonctions[23],[24].
Pathologies transmises par des hématophages
Exemples :
- Malaria (= Paludisme), due à un protozoaire (Plasmodium falciparum) injecté lors de la piqûre des moustiques femelles du genre Anopheles
- Filariose lymphatique (= Eléphantiasis), due à des vers parasites filiformes (filaires) Wuchereria bancrofti et Brugia malayi transmis par plusieurs genres de moustiques (Anopheles, Culex, Aedes,...) ;
- Dengue, virus transmis par des moustiques femelles du genre Aedes ;
- Onchocercose (= « cécité des rivières »), vers filiformes de l'espèce Onchocerca volvulus, transmis par des mouches noires de la famille des Simuliidés ;
- Leishmanioses cutanées; due à des protozoaires flagellés (du genre Leishmania, famille des Trypanosomidae), transmis par des diptères du genre Phlebotomus et du genre Lutzomyia ;
- Trypanosomiase américaine (= « Maladie de Chagas »), due à un protozoaire (trypanosome ) ; Trypanosoma cruzi transmis via les excréments de punaises hématophages de la sous-famille des triatomines ;
- Trypanosomiase (= « maladie du sommeil ») due à un parasite trypanosome, Trypanosoma brucei, transmis par la piqûre de différentes mouches du genre Glossina (dites « mouches tsé-tsé » ou « moutmout » en Afrique) ;
- rage et quelques autres maladies (par des morsures souvent accidentelles de chauves-souris parfois) ;
- Infection à virus Zika
Écoépidémiologie
De nombreux insectes hématophages sont des insectes vecteurs de maladies. Chez ces insectes, souvent il n'y a que la femelle qui soit hématophage, en préparation de la formation des œufs, pour laquelle elle a besoin d'un apport important de protéines.
On a récemment montré qu'il existe de nombreux facteurs endogènes expliquant l'attractivité d'un individu pour un hématophage[1].
Certains parasites sont littéralement capables de manipuler leur hôte pour le rendre plus "attractif" pour les hématophages, ce qui leur permet de diffuser leurs gènes et descendance.
Voir aussi
Articles connexes
Bibliographie
- (fr) Aurélie Bodin, Modulation du comportement de recherche de l'hôte chez les insectes hématophages : Importance des facteurs endogènes ; Thèse de doctorat en Sciences de la Vie (Université François Rabelais), 2008-10-31, PDF, 145 pages
Références
- Aurélie Bodin, Modulation du comportement de recherche de l'hôte chez les insectes hématophages : Importance des facteurs endogènes ; Thèse de doctorat en Sciences de la Vie (Université François Rabelais), 2008-10-31, PDF, 145 pages
- (en) M. M. J. Lavoipierre, « Feeding mechanism of blood-sucking arthropods », Nature, vol. 208, no 5007, , p. 302-303.
- Marcel Leclercq, Les mouches, nuisibles aux animaux domestiques, Vander, , p. 25.
- Nicolas Valeix, Parasitologie. Mycologie, De Boeck Superieur, , p. 5.
- Lallement, H. (2007). Habituation et conditionnement classique aversif de la réponse d’extension du proboscis chez un insecte hématophage, Rhodnius prolixus. Rapport de stage M2, Université de Tours, 66 pp
- Thèse d'Aurélie Bodin, déjà citée (p. 30/145).
- Lazzari, C.R. et Núnez, J. (1989). The response to radiant heat and the estimation of the temperature of distant sources in Triatoma infestans. Journal of Insect Physiology, 35: 525-529.
- Ferreira, R.A., Lazzari, C.R., Lorenzo, M.G. et Pereira, M.H. (2007), Do haematophagous bugs assess skin surface temperature to detect blood vessels ?, PLoS One 2, e932, DOI:10.1371/journal.pone.0000932.
- Bogner, F. (1992), Response properties of CO2-sensitive receptors in tse-tse flies (Diptera: Glossina palpalis). Physiological Entomology, 17: 19-24.
- Bowen, M.F., Davis, E.E., Haggart, D. et Romo, J. (1994). Host-seeking behavior in the autogenous mosquito Aedes atropalpus, Journal of Insect Physiology, 40: 511-517.
- Bowen, M.F. et Davis, E.E. (1989). The effects of allatectomy and juvenile hormone replacement on the development of host-seeking behaviour and lactic-acid receptor sensitivity in the mosquito, Aedes aegypti. Medical and Veterinary Entomology, 3: 53- 60.
- Guerenstein, P.G. et Núñez, J.A. (1994). Feeding response of the haematophagous bugs Rhodnius prolixus and Triatoma infestans to saline solutions: a comparative study, Journal of Insect Physiology, 40: 747-752.
- Friend, W.G. et Smith, J.J.B. (1977). Factors affecting feeding by blood-sucking insects, Annual Review of Entomology, 22: 309-331.
- News du magazine Science (2017), Watch flies suck this frog’s blood by eavesdropping on its mating calls, 29 novembre 2017.
- Lorenzo, M.G. et Lazzari, C.R. (1998). Activity pattern in relation to refuge exploitation and feeding in Triatoma infestans (Hemiptera: Reduviidae). Acta Tropica, 70: 163-170
- Lorenzo Figueiras, A.N., Kenigsten, A. et Lazzari, C.R. (1994). Aggregation in the haematophagous bug Triatoma infestans: chemical signals and temporal pattern. Journal of Insect Physiology, 40: 311-316.
- Lorenzo Figueiras, A.N. et Lazzari, C.R. (1998). Aggregation in the haematophagous bug Triatoma infestans: a novel assembling factor. Physiological Entomology, 23: 33-37.
- Lorenzo, M.G. et Lazzari, C.R. (1996). The spatial pattern of defecation in Triatoma infestans and the role of faeces as a chemical mark of the refuge. Journal of Insect Physiology, 42: 903-907.
- Lorenzo, M.G. et Lazzari, C.R. (1998). Activity pattern in relation to refuge exploitation and feeding in Triatoma infestans (Hemiptera: Reduviidae). Acta Tropica, 70: 163-170.
- Bodin, A., Barrozo, R.B., Couton, L. et Lazzari, C.R. (2008). Temporal modulation and adaptive control of the behavioural response to odours in Rhodnius prolixus. J. Insect Physiol. doi:10.1016/j.jinsphys.2008.07.004
- Chapman, R.F. (1982). Chemoreception: the significance of receptor numbers. Advances in Insects Physiology, 16: 247-356
- Lehane, M. (2005). The biology of blood-sucking in insects. Second edition. Cambridgeuniversity press, Cambridge
- Liang, D. et Schal, C. (1990). Circadian rhythmicity and development of the behavioural response to sex pheromone in male brown-banded cockroach, Supella longipalpa. Physiological Entomology, 15: 355-361
- Linn, C.E., Campbell, M.G., Poole, K.R., Wu, W.Q. et Roelofs, W.L. (1996). Effects of photoperiod on the circadian timing of pheromone response in male Trichoplusia ni: Relationship to the modulatory action of octopamine. Journal of Insect Physiology, 42: 881-891.
 Portail de la zoologie
Portail de la zoologie