Triade catalytique
En biologie moléculaire, on désigne généralement par triade catalytique les trois résidus d'acides aminés qui interviennent ensemble dans le site actif de certaines hydrolases et transférases telles que des peptidases, des amidases, des estérases, des lipases et des β-lactamases. Une façon courante de produire des résidus nucléophiles pour une catalyse covalente consiste à faire intervenir une triade acide–base–nucléophile[2],[3]. Ces résidus forment un réseau de transfert de charges qui polarise et active le résidu nucléophile, lequel attaque le substrat en formant un intermédiaire covalent hydrolysé ensuite pour libérer à nouveau l'enzyme. Le nucléophile d'une telle triade est généralement la sérine ou la cystéine, mais peut également être la thréonine.
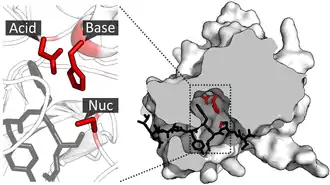
Les résidus constituant la triade catalytique peuvent être très éloignés les uns des autres le long de la séquence peptidique (structure primaire) des enzymes considérées, mais se retrouver à proximité les uns des autres en raison du repliement des polypeptides en une structure tertiaire tridimensionnelle complexe.
Les triades catalytiques constituent l'un des meilleurs exemples de convergence évolutive : la nature même des réactions chimiques catalysées ont conduit de façon indépendante aux mêmes types de sites actifs à partir d'au moins 23 superfamilles distinctes[3]. Leur mécanisme réactionnel est par conséquent l'un des mieux compris de toute la biochimie[4].
Constituants d'une triade catalytique
Nucléophile
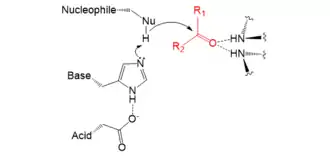
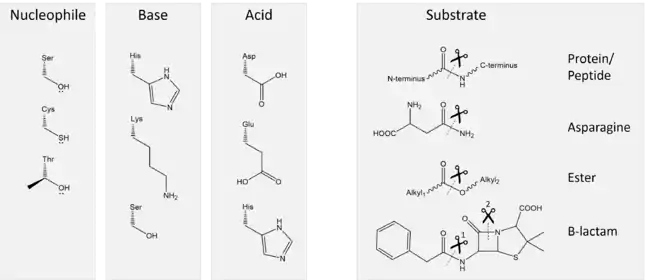
La chaîne latérale d'un résidu nucléophile réalise la catalyse covalente sur le substrat. Le doublet non liant de l'atome d'oxygène ou de soufre attaque l'atome de carbone du carbonyle, qui est électropositif. Aucun des 22 acides aminés protéinogènes ne possèdent de groupes fonctionnels suffisamment nucléophiles pour catalyser de nombreuses réactions difficiles.
Les principaux nucléophiles enzymatiques sont l'hydroxyle OH du groupe alcool de la sérine et le sulfhydryle SH du groupe thiol ou de l'ion thiolate S− de la cystéine. Inclure ces nucléophiles dans une triade catalytique accroît leur activité. Quelques peptidases utilisent l'alcool secondaire de la thréonine ; cependant, en raison de la présence d'un groupe méthyle supplémentaire, ces peptidases utilisent l'amide N-terminale comme base plutôt qu'un autre résidu d'acide aminé[3],[5].
Base
Aucun acide aminé naturel n'étant suffisamment nucléophile pour catalyser seul les réactions chimiques en question, les triades catalytiques font intervenir une base qui polarise et déprotone le nucléophile afin d'accroître sa réactivité. Cette base déprotone également le premier produit de réaction pour favoriser l'élimination du groupe partant.
C'est généralement un résidu d'histidine qui joue le rôle de base dans les triades catalytiques, étant donné que son pKa permet à la fois une catalyse efficace, la formation d'une liaison hydrogène avec le résidu d'acide, et la déprotonation du résidu nucléophile. Les β-lactamases de type TEM-1 utilisent quant à elles un résidu de lysine. La valeur très élevée du pKa de la lysine — voisine de 11 — est compensée par un résidu de glutamate et plusieurs autres résidus, qui agissent pour stabiliser sa forme déprotonée pendant le cycle catalytique[6]. Afin de limiter l'encombrement stérique, les protéases à thréonine utilisent leur amide N-terminale comme base afin d'accroître la réactivité catalytique du résidu de thréonine[7],[8].
Acide
Le résidu acide agit en alignant et en polarisant le résidu basique. Il s'agit généralement d'un résidu d'aspartate ou de glutamate. Certaines enzymes se contentent en fait d'une « dyade catalytique » dans la mesure où la composante acide peut n'être pas nécessaire à certaines protéases à cystéine. Ainsi, la papaïne utilise l'asparagine comme troisième membre de la triade, celle-ci ne pouvant en aucun cas agir comme acide mais simplement orienter convenablement l'histidine pour la réaction. De la même façon, la peptidase du virus de l'hépatite A contient une molécule d'eau là où devrait se trouver le résidu acide. Enfin, les peptidases de cytomégalovirus utilisent deux résidus d'histidine contigus, l'un agissant comme base et l'autre à la place normalement occupée par un résidu acide[3] ; n'étant pas acide, mais au contraire basique, ce second résidu d'histidine conduit à une efficacité catalytique inférieure pour cette enzyme.
Exemples de triades
Ser-His-Asp
La chymotrypsine est considérée comme l'un des paradigmes des enzymes à triade catalytique, ici la triade sérine–histidine–aspartate.
- La chymotrypsine se lie à son substrat au niveau d'une boucle contenant un grand résidu hydrophobe.
- L'aspartate forme une liaison hydrogène avec l'histidine, ce qui fait passer de 7 à près de 12 le pKa de l'atome d'azote du cycle imidazole. Ceci permet à l'histidine d'agir comme une base forte et de déprotoner la sérine.
- La sérine intervient comme nucléophile en attaquant l'atome de carbone du carbonyle et forçant l'atome d'oxygène de ce carbonyle à accepter un électron, ce qui conduit à la formation d'un intermédiaire tétraédrique. Cet intermédiaire est stabilisé par un trou oxyanion impliquant l'amide de la sérine.
- L'histidine donne ensuite son proton à l'atome d'azote du carbone α. L'azote et le fragment peptidique C-terminal sont éliminés par diffusion.
- Une molécule d'eau donne alors un proton à l'histidine et l'hydroxyle –OH restant attaque l'atome de carbone du carbonyle, formant un autre intermédiaire tétraédrique. L'hydroxyle est un moins bon nucléofuge que le fragment peptidique C-terminal de sorte que c'est la sérine qui est éliminée en récupérant un proton de l'histidine.
- Le fragment peptidique N-terminal est éliminé par diffusion.
La même triade a également fait l'objet d'une évolution convergente dans les repliements α/β hydrolase telles que certaines lipases et estérases, mais la chiralité y est cependant inversée. L'acétylcholinestérase utilise l'équivalent sérine–histidine–glutamate.
Cys-His-Asp
Plusieurs familles de protéases à cystéine utilisent cette combinaison d'acides aminés comme triade catalytique, par exemple la protéase TEV et la papaïne. Cette triade à cystéine agit comme celle des protéases à sérine mais avec cependant quelques différences notables. En particulier, le rôle de l'aspartate dans la triade de la papaïne n'est pas clair et plusieurs protéases à cystéine utilisent en réalité une dyade catalytique, comme la peptidase du virus de l'hépatite A.
Ser-His-His
La triade catalytique de la peptidase du cytomégalovirus utilise deux résidus d'histidine contigus à l'emplacement de la base et de l'acide. Retirer l'histidine « acide » conduit à une activité seulement dix fois moindre, alors que le retrait du résidu d'aspartate de la chymotrypsine conduit à une activité 10 000 fois moindre. Cette triade particulière a été interprétée comme une possible manière de produire une enzyme à l'activité réduite afin de contrôler le taux de clivage[5].
Notes et références
- (en) Jason Phan, Alexander Zdanov, Artem G. Evdokimov, Joseph E. Tropea, Howard K. Peters III, Rachel B. Kapust, Mi Li, Alexander Wlodawer et David S. Waugh, « Structural Basis for the Substrate Specificity of Tobacco Etch Virus Protease », Journal of Biological Chemistry, vol. 277, no 52, , p. 50564-50572 (PMID 12377789, DOI 10.1074/jbc.M207224200, lire en ligne)
- (en) Andrew R. Buller et Craig A. Townsend, « Intrinsic evolutionary constraints on protease structure, enzyme acylation, and the identity of the catalytic triad », Proceedings of the National Academy of Sciences of the United States of America, vol. 110, no 8, , E653-E661 (PMID 23382230, PMCID 3581919, DOI 10.1073/pnas.1221050110, lire en ligne)
- (en) Guy Dodson et Alexander Wlodawer, « Catalytic triads and their relatives », Trends in Biochemical Sciences, vol. 23, no 9, , p. 347-352 (PMID 9787641, DOI 10.1016/S0968-0004(98)01254-7, lire en ligne)
- (en) Hans Neurath, « Proteolytic enzymes past and present: The second golden era », Protein Science, vol. 3, no 10, , p. 1734-1739 (PMID 7849591, PMCID 2142620, DOI 10.1002/pro.5560031013, lire en ligne)
- (en) Özlem Doğan Ekici, Mark Paetzel et Ross E. Dalbey, « Unconventional serine proteases: Variations on the catalytic Ser/His/Asp triad configuration », Protein Science, vol. 17, no 12, , p. 2023-2037 (PMID 18824507, PMCID 2590910, DOI 10.1110/ps.035436.108, lire en ligne)
- (en) C. Damblon, X. Raquet, L. Y Lian, J. Lamotte-Brasseur, E. Fonze, P. Charlier, G. C Roberts et J. M Frère, « The catalytic mechanism of beta-lactamases: NMR titration of an active-site lysine residue of the TEM-1 enzyme », Proceedings of the National Academy of Sciences of the United States of America, vol. 93, no 5, , p. 1747-1752 (PMID 8700829, PMCID 39852)
- (en) James A. Brannigan, Guy Dodson, Helen J. Duggleby, Peter C. E. Moody, Janet L. Smith, Diana R. Tomchick et Alexey G. Murzin, « A protein catalytic framework with an N-terminal nucleophile is capable of self-activation », Nature, vol. 378, no 6555, , p. 416-419 (PMID 7477383, DOI 10.1038/378416a0, lire en ligne)
- (en) Hua Cheng et Nick V. Grishin, « DOM-fold: A structure with crossing loops found in DmpA, ornithine acetyltransferase, and molybdenum cofactor-binding domain », Protein Science, vol. 14, no 7, , p. 1902-1910 (PMID 15937278, PMCID 2253344, DOI 10.1110/ps.051364905, lire en ligne)
 Portail de la biochimie
Portail de la biochimie  Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire