بروتين فيروسي
البروتين الفيروسي هو البروتين الذي ينتج من ترجمة الحمض النووي للفيروس وهو إما جزء منه أو لا يُنتج سوى أثناء تضاعف الفيروس أو كمونه. تصنف الفيروسات البروتينية حسب وظيفتها إلى: بنيوية، لابنيوية، تنظيمية وملحقة.[1] الفيروسات هي كائنات غير حية ولا تمتلك الوسائل لتتكاثر لوحدها، وفي سبيل ذلك تعتمد على أيض الخلية المضيفة للحصول على الطاقة وعلى إنزيماتها وموادها الأولية. لذلك لا يشفر جينوم الفيروسات العديد من البروتينات التي تستخدمها وبدل ذلك تستخدم ماكينة الخلية المضيفة لإنتاج البروتينات الفيروسية التي تحتاجها للتكاثر.[2]

بروتينات بنيوية فيروسية
معظم البروتينات البنيوية هي مكونات للقفيصة أو الغلاف الفيروسي:
القفيصة
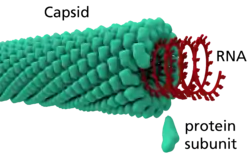
المادة الحاملة للمعلومات الجينية الخاصة بالفيروس مخزنة داخل بنية بروتينية تسمى القفيصة، والقفيصة هي «الدرع» الذي يحمي الحمض النووي الفيروسي من تفكيكه بواسطة إنزيمات الخلية المضيفة أو أنواع أخرى من المبيدات أو المعديات. وتعمل كذلك على ربط الفيروس بالخلية المضيفة وتمكين الفيروس من اختراق غشائها. تتشكل القفيصة من نسخ عديدة من بروتين بنيوي واحد أو عدة بروتينات بنيوية مختلفة، وكل واحد من هذه البروتينات الفيروسية يشفَّر بواسطة جين واحد من الجينوم الفيروسي. تصميم بنية القفيصة يسمح للفيروس باستخدام عدد صغير من الجينات الفيروسية لتشكيل قفيصة كبيرة.[3]
ترتبط العديد من القسيمات البروتينية - وهي بروتين قليل القسيمات مكونة على الأقل من سلسلتي بروتينين- لتشكل قسيم قفيصي وبدورها ترتبط القسيمات القفيصية لتشكل القفيصة.[1] يمكن أن تترتب القسيمات القفيصية على شكل عشروني السطوح، لولبي، مخروطي، أو بنية معقدة أخرى. لكن قفيصة العديد من الفيروسات مثل فيروس الهربس البسيط تكون على شكل عشروني السطوح.[2] يتكون كل وجه من الوجوه العشرين المثلثة الخاصة بعشريني السطوح من ثلاث وحدات بروتينية لامتناظرة.[2]
الغلاف الفيروسي
قفيصة بعض الفيروسات محاطة بغشاء يسمى الغلاف الفيروسي. في معظم الحالات، تحصل القفيصة على غلافها من الغشاء الخلوي للخلية المضيفة حين تغادرها عبر عملية تسمى التبرعم.[4] يتكون الغلاف الفيروسي من ليبيد ثنائي الطبقة مرصع ببروتينات فيروسية منها البروتينات السكرية الفيروسية.[1] ترتبط هذه البروتينات السكرية بمستقبلات ومستقبلات مساعدة محددة على سطح غشاء الخلية المضيفة وتسمح للفيروس بالارتباط بالخلية الهدف والدخول إليها.[1] بعض هذه البروتينات السكرية:
- الراصة الدموية، نورامينيداز وبروتين M2 في فيروس الإنفلونزا.
- إنف المكون من الوحدات الفرعية gp120 والبروتين السكري gp41 في فيروس العوز المناعي البشري.[1]

تلعب البروتينات السكرية الفيروسية دورا حاسما في اندماج غلاف الفيروس بالخلية، ويبدأ ذلك الاندماج عندما ترتبط البروتينات السكرية بالمستقبلات الخلوية.[5]
بروتينات دمج الغشاء الفيروسية
اندماج الغلاف الفيروسي مع الغشاء الخلوي يتطلب طاقة كبيرة ليحدث، وتعمل بروتينات دمج الغشاء الفيروسية كمحفزات للتغلب على حاجز الطاقة العالي هذا.[6] بعد ارتباط البروتين السكري الفيروسي بالمستقبلات الغشائية، تخضع بروتينات دمج الغشاء الفيروسية لتغير في هيئتها البنيوية.[6] يسهل هذا التغير في الهيئة الإخلال باستقرار الغشاء الغلوي واندماجه مع الغلاف الفيروسي عبر السماح لحلقات الاندماج (FLs) أو بروتينات الدمج الكارهة للماء الموجودة على الغلاف الفيروسي بالتآثر مع الغشاء الخلوي.[6] معظم بروتينات دمج الغشاء الفيروسية ينتهي بها الحال على هيئة حلقة جذعية بعد الاندماج وفيها تكون جميع الـ FLs/FPs والنطاق عبر الغشائي على نفس الجانب من البروتين.[6]
سمحت البروتينات السكرية الفيروسية وبنيتها ثلاثية الأبعاد -قبل وبعد الاندماج- باكتشاف طيف واسع من الهيئات البنيوية.[6] تم تصنيف بروتينات دمج الغشاء الفيروسية إلى أربعة أقسام مختلفة وكل قسم يمتلك خصائص هيئات بنيوية محددة:
- القسم 1: هيئة ما بعد الاندماج لها مركز مميز ببنية لفة ملتفة مكونة من مثلوث من لوالب ألفا مميزة على شكل حلقة جذعية، ومن الأمثلة على هذا القسم بروتين الدمج الخاص بفيروس العوز المناعي gp41.[6]
- القسم 2: بروتينات لا تملك مركزا بنيته لفة ملتفة. وتحتوي بنية نطاق خاري مميز عبارة عن صحيفة بيتا ممتدة والتي تعيد الانطواء لتشكل مثلوث حلقات جذعية، من الأمثلة على القسم الثاني بروتين E الخاص بفيروس الضنك وبروتين E الخاص بفيروس غرب النيل.[5][6]
- القسم 3: هيئية بنيوية تكون توليفة بين هيئتي القسمين 1 و2 من الأمثلة على ذلك البروتين السكري G الخاص بفيروس داء الكلب.[6]
- القسم 4: هي بروتينات عبر غشائية صغيرة مرتبطة بالدمج (FAST)، ولا تشكل مثلوثاتِ حلقات جذعية أو بنى حلقات جذعية في حد ذاتها، وهي أصغر بروتينات دمجٍ فيروسية معروفة. بروتينات FAST تشفرها أعضاء فيروسات من عائلة فيروسات الريو غير المغلفة.[6]
بروتينات لابنيوية فيروسية
البروتينات اللابنيوية الفيروسية هي بروتينات يشفِّرها جينوم الفيروس ويعبر عنها في الخلية المصابة، لكن لا يتم تجميعها داخل الفيروس ولا تكون جزءا من بنيته.[1] أثناء تضاعف الفيروسات، تقوم بعض البروتينات اللابنيوية بوظائف حاسمة تؤثر على عملية التضاعف في حد ذاتها. وبالمثل، تقوم بعض هذه البروتينات كذلك أثناء تجميع الفيروس بوظائف مهمة تؤثر على عملية التجميع. من بعض وظائف هذه البروتينات اللابنيوية تشكيل الريبليكون، التعديل المناعي، والتنشيط المفروق للجينات المشفِّرة للبروتينات الفيروسية.[1]
تكوين الريبلكون
تتآثر بروتينات الفيروس اللابنيوية مع بروتينات الخلية المضيفة لتشكيل الريبلكون والذي يعرف كذلك بمركب التضاعف.[1] في فيروس التهاب الكبد C، تتآثر البروتينات اللابنيوية مع بروتين النقل الخاص بالغشاء الحويصلي hVAP-33 لتجميع الريبلكون.[1] يغير البروتين اللابنيوي الفيروسي 4b أو (NS4B) غشاء الخلية المضيفة ويبدأ عملية تكوين مركب التضاعف.[1][7] توظَّف بروتينات لابنيوية فيروسية أخرى مثل NS5A وNS5B وNS3 في المركب كذلك ويتآثر البروتين NS4B معهم ويرتبط بالرنا الفيروسي.[1][7]
التعديل المناعي
يمكن أن تُعدَّل الاستجابة المناعية الخاصة بالمضيف ضد الخلية المصابة عبر خصائص التعديل المناعي الخاصة بالبروتينات اللابنيوية الفيروسية.[1] تشفِّر العديد من أجناس فيروسات الدنا الطويل بروتيناتٍ تخرب الاستجابة المناعية للمضيف، وهذا يسمح بتكاثر الفيروس.[8] فهم وظيفة مثل تلك البروتينات يوفر إمكانية تطوير علاجات داوئية حيوية للاستجابات الالتهابية لدى البشر، وذلك لثبوت أن هذه البروتينات تقوم بتخريب العوامل المناعية المتوسطة للاستجابة الالتهابية.[9] يمنع البروتين اللابنيوي الفيروسي NS1 في فيروس غرب النيل تنشيط النظام المتمم عبر ارتباطه ببروتين مراقبة النظام المتمم العامل H [الإنجليزية] .[1] ونتيجة لذلك ينخفض التعرف المتمم في الخلايا المصابة، وتبقى هذه الأخيرة غير متضررة بواسطة نظام مناعة المضيف.
بروتينات منظمة وملحقة فيروسية
البروتينات الفيروسية المنظِمة والملحقة لها وظائف متعددة. تتحكم هذه البروتينات في التعبير الجيني عن جينوم الفيروس وتؤثر عليه بما في ذلك معدلات نسخ جينات البروتينات البنيوية.[1] كما تؤثر على الوظائف الخلوية للخلية المضيفة مثل التنظيم الجيني وموت الخلية المبرمج.[1] في فيروسات الدنا والفيروسات الراجعة، يمكن أن تحسن البروتينات الفيروسية المنظِمة النسخ الجيني الفيروسي وكذلك يمكن لهذه البروتينات تحسين النسخ الجيني الخلوي الخاص بالمضيف.[10]
تُشفَّر البروتينات الفيروسية الملحقة -وتعرف كذلك بالبروتينات المساعِدة أو الإضافية أو الثانوية- بواسطة جينوم الفيروسات الراجعة.[11] معظم البروتينات الفيروسية الملحقة يمكنها القيام بوظائفها في أنواع محددة من الخلايا فقط.[11] وكذلك ليس لها تأثير كبير على تضاعف الفيروس، لكن في بعض الأحيان يتطلب الحفاظ على عملية تضاعف الفيروس مساعدة البروتينات الفيروسية الملحقة.
بروتنيات راجعة داخلية المنشأ
سينسيتين 1 هو فيروس راجع داخلي المنشأ موجود في جينوم الثدييات ويسمح بالاندماج الغشائي لخلايا الأرومة الغاذية الخلوية في التخلق المشيمي.[12]
المراجع
- Uversky, Longhi, Vladmir, Sonia (2011)، Flexible Viruses، Wiley، ص. 4، ISBN 9781118135549.
- Slonczewski, Foster, Joan, John (2013)، Microbiology: An Evolving Science. Third Edition، W. W. Norton & Company، ص. 192–195، ISBN 978-0393123678.
- Lodish, Harvey؛ Berk؛ Zipursky؛ Matsudaira؛ Baltimore؛ Darnell (01 يناير 2000)، "Viruses: Structure, Function, and Uses" (باللغة الإنجليزية)، مؤرشف من الأصل في 12 أبريل 2020.
{{استشهاد بدورية محكمة}}: Cite journal requires|journal=(مساعدة) - Pornillos, Owen؛ Garrus؛ Sundquist (01 ديسمبر 2002)، "Mechanisms of enveloped RNA virus budding"، Trends in Cell Biology، 12 (12): 569–579، doi:10.1016/S0962-8924(02)02402-9، PMID 12495845.
- White, Judith M.؛ Delos؛ Brecher؛ Schornberg (01 يناير 2008)، "Structures and Mechanisms of Viral Membrane Fusion Proteins"، Critical Reviews in Biochemistry and Molecular Biology، 43 (3): 189–219، doi:10.1080/10409230802058320، ISSN 1040-9238، PMC 2649671، PMID 18568847.
- Podbilewicz, Benjamin (2014)، "Virus and Cell Fusion Mechanisms"، Annual Review of Cell and Developmental Biology، 30 (1): 111–139، doi:10.1146/annurev-cellbio-101512-122422، PMID 25000995.
- Gouttenoire, Jérôme؛ Penin؛ Moradpour (01 مارس 2010)، "Hepatitis C virus nonstructural protein 4B: a journey into unexplored territory"، Reviews in Medical Virology، 20 (2): 117–129، doi:10.1002/rmv.640، ISSN 1099-1654، PMID 20069613.
- Engel؛ Angulo (2012)، "Viral immunomodulatory proteins: usurping host genes as a survival strategy."، Advances in Experimental Medicine and Biology، 738: 256–278، doi:10.1007/978-1-4614-1680-7_15، ISBN 978-1-4614-1679-1، PMID 22399384.
- Lucas؛ McFadden (2004)، "Secreted immunomodulatory viral proteins as novel biotherapeutics."، J Immunol، 173 (8): 4765–74، doi:10.4049/jimmunol.173.8.4765، PMID 15470015.
- Flint, Jane؛ Shenk (1997)، "Viral Transactivating Proteins"، Annual Review of Genetics، 31 (1): 177–212، doi:10.1146/annurev.genet.31.1.177، PMID 9442894.
- Coffin, John M.؛ Hughes؛ Varmus (01 يناير 1997)، "Accessory Proteins and Assembly" (باللغة الإنجليزية)، Cold Spring Harbor Laboratory Press، مؤرشف من الأصل في 3 مايو 2016.
{{استشهاد بدورية محكمة}}: Cite journal requires|journal=(مساعدة) - Mi S, Lee X, Li X, Veldman GM, Finnerty H, Racie L, LaVallie E, Tang XY, Edouard P, Howes S, Keith JC Jr, McCoy JM (2000)، "Syncytin is a captive retroviral envelope protein involved in human placental morphogenesis"، Nature، 403 (6771): 785–9، Bibcode:2000Natur.403..785M، doi:10.1038/35001608، PMID 10693809.
| المكونات |  | |
|---|---|---|
| دورة حياة الفيروس | ||
| الجينات | ||
| حسب العائل |
| |
| أخرى |
| |
| ||
 بوابة علم الفيروسات
بوابة علم الفيروسات بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي