بروتين راسي بالدهون
بروتينات راسية بالدهون (المعروفة أيضًا بالبروتينات المرتبطة بالدهون) (بالإنجليزية: Lipid anchored proteins) هي نوع من البروتيوليبيدات، وهي بروتينات موجودة على سطح غشاء الخلية وترتبطها روابط تساهمية بالدهون الموجودة داخل غشاء الخلية. تدخل هذه البروتينات وتحتل مكاناً في بنية الطبقة الثنائية للغشاء جنباً إلى جنب مع ذيول الأحماض الدهنية المماثلة. يمكن وضع البروتين المرتبط بالدهون على جانبي غشاء الخلية. وهكذا، يعمل الدهن على تثبيت البروتين في غشاء الخلية.[1][1]

تلعب مجموعات الدهون دوراً في تفاعل البروتين ويمكن أن تساهم في وظيفة البروتين الذي ترتبط به. علاوة على ذلك، يعمل الدهن كوسيط لترابطات الأغشية أو كمحدد لتفاعلات بروتينية معينة.[2] على سبيل المثال، يمكن أن تلعب مجموعات الدهون دوراً مهماً في زيادة كراهية الماء الجزيئي. هذا يسمح بتفاعل البروتينات مع الأغشية الخلوية ومجالات البروتين.[2] في دور ديناميكي، يمكن للدهون عزل البروتين بعيداً عن ركائزه لتعطيل البروتين ثم تنشيطه عن طريق عرض الركيزة.
بشكل عام، هناك ثلاثة أنواع رئيسية من البروتينات المرتبطة بالدهون والتي تشمل البروتينات السابقة، والبروتينات الدهنية أسيلية والبروتينات المرتبطة بالجليكوزيل فوسفاتيديلينوسيتول (GPI).[3] يمكن أن يحتوي البروتين على مجموعات دهنية متعددة مرتبطة به تساهمياً، لكن الموقع الذي يرتبط فيه الدهون بالبروتين يعتمد على كل من مجموعة الدهون والبروتين.
بروتينات مسبقة
البروتينات المسبقة هي بروتينات تحتوي على بوليمرات إيزوبرين (أي هيدروكربون متفرع من خمسة كربون)[2] كارهة للماء مرتبطة تساهمياً في بقايا السيستين للبروتين. وبشكل أكثر تحديداً، ترتبط مجموعات الأيزوبرينويد، عادة فارنيسل (15-كربون) أو جيرانيل جيرانيل (20-كربون) عبر روابط ثيوإيثر مع بقايا السيستين بالقرب من الطرف الكربوكسيلي للبروتين(الطرف C). وهذه البرنلة من سلاسل الدهون تعمل على تسهيل تفاعل البروتينات مع الأغشية الخلوية.

يعد "CAAX" هو أكثر موقع شيوعاً لبرنلة البروتينات، أي الموقع الذي يرتبط فبه فارنيسيل أو جيرانيل جيرانيل تساهمياً. في تسلسل "CAAX"، يمثل الحرف C السيستين الذي يتم معالجته مسبقا، ويمثل الحرف A أي حمض أميني أليفاتي، والحرف X يمثل نوع البرنلة التي ستحدث. إذا كان X هو حمض ألانين أو ميثيونين أو سيرين أو جلوتامين؛ سيتم إضافة مجموعة فارنيسل عن طريق أنزيم فارنيسل ترانسفيراز إلى البروتين، وإذا كان X هو حمض ليوسين؛ فإنه سيتم إضافة مجموعة جيرانيل جيرانيل عن طريق أنزيم جيرانيل جيرانيل ترانسفيراز نوع 1. كل من هذه الإنزيمات متشابهة مع كل منها يحتوي على وحدتين فرعيتين.[4]
الدور والوظيفة
تعتبر البروتينات المسبقة مهمة بشكل خاص في نمو الخلايا حقيقية النواة والتمايز والتشكل. علاوة على ذلك، فإن برنلة البروتين هو تعديل قابل للانعكاس بعد الترجمة لغشاء الخلية. هذا التفاعل الديناميكي للبروتينات مسبقة الصنع مع غشاء الخلية مهم لوظائفها في الإشارة وغالبا ما يتم تحريره في عمليات الأمراض مثل السرطان.[4] وبشكل أكثر تحديداً، بروتين راس الذي يخضع للبرنلة عبر فارنيسيل ترانزفيراز وعندما يتم تشغيله يمكنه تشغيل الجينات المشاركة في نمو الخلايا وتمايزها. بالتالي يمكن أن يؤدي الإفراط في إشارات راس إلى الإصابة بالسرطان.[4] كان فهم هذه البروتينات المسبقة وآلياتها مهماً لجهود تطوير الأدوية في مكافحة السرطان.[4] وتشمل البروتينات المسبقة الأخرى أعضاء من عائلات راب ورو وكذلك لامينات.
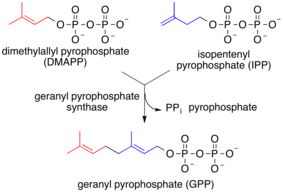
بعض سلاسل البرنلة هامة تشارك في مسار اختزال HMG-CoA وهي جيرانيل جيرانيول وفارنيسول ودوليكول. وتشارك هذه البوليمرات إيزوبرين (مثل بايروفوسفات جيرانيل وبايروفوسفات فارنيسيل) في التكثيف عن طريق الإنزيمات مثل برينيل ترانسفيراز الذي يتحول في النهاية إلى الكولسترول.
بروتينات دهنية أسيلية
البروتينات الدهنية الأسيلية هي بروتينات تم تعديلها بعد الترجمة لتشمل الارتباط التساهمي للأحماض الدهنية في بعض بقايا الأحماض الأمينية.[4] الأحماض الدهنية الأكثر شيوعاً التي ترتبط تساهمياً بالبروتين هي حمض الميريستيك المشبع (14-كربون) وحمض البالمتيك (16-كربون)، يمكن تعديل البروتينات لاحتواء أحد هذه الأحماض الدهنية أو كليهما.
ن-ميريستول
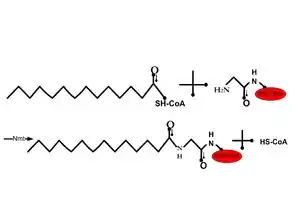
ن-ميريستول (أي ارتباط حمض ميريستيك) بشكل عام هو تعديل بروتين لا رجعة فيه، يحدث عادة أثناء تخليق البروتين، حيث يرتبط حمض ميريسيتك مع مجموعة ألفا أمين بالنهاية الطرفية N من بقايا جلايسين عبر رابطة أميدية.[4] يتم تسهيل هذا التفاعل بواسطة ن-ميريستويل ترانسفيراز. تبدأ هذه البروتينات عادة بتسلسل ميثيونين-جلايسين ومع إما سيرين أو ثريونين في الموضع 5. وتشارك ميريستويلاتد بروتين في سلسلة تحويل الإشارة وتفاعلات البروتين- البروتين وفي الآليات التي تنظم استهداف البروتين ووظيفته.[4] أحد الأمثلة على أهمية ن-ميريستول للبروتين هوالاستموات وموت الخلايا المبرمج. بعد أن يتم ربط ناهض موت المجال التفاعلي Bid بالبروتين BH3، فإنه يستهدف البروتين للانتقال إلى غشاء الميتوكوندريا لإطلاق السيتوكروم سي؛ مما يؤدي في نهاية المطاف إلى موت الخلايا.[4] وهنك بروتينات ميريستويلاتد أخرى تشارك في تنظيم موت الخلايا المبرمج هي الأكتين والجيلسولين.
س-بالميتويليشن
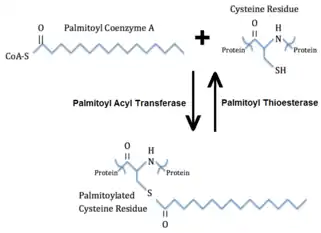
س-بالميتويليشن (أي ارتباط حمض البالمتيك) هو تعديل البروتين قابل للانعكاس، حيث يتم بط حمض البالمتيك مع بقايا السيستين محددة عبر رابطة ثيوايستر. يمكن أيضا استخدام مصطلح أسيلة عندما ترتبط سلاسل الأحماض الدهنية المتوسطة والطويلة الأخرى بالبروتينات بالميتويلاتد. توجد البروتينات بالميتويلاتد بشكل رئيسي على الجانب السيتوبلازمي من غشاء البلازما حيث تلعب دورا في إشارات الغشاء. يمكن إزالة مجموعة بالميتويل بواسطة ثيوستراز بالميتويل. ويعتقد أن عكس بالميتويل قد ينظم تفاعل البروتين مع الغشاء وبالتالي يكون له دور في عمليات الإشارة. علاوة على ذلك، يسمح هذا بتنظيم توطين البروتينات تحت الخلوية واستقرارها.[4] أحد الأمثلة التي يلعب فيها بالميتويل للبروتين دورا في مسارات إشارات الخلية هو تجميع البروتينات في المشبك العصبي. عندما تكون كثافة بالميتويلاتد بروتين بعد المشبكي 95(PSD-95)، فإنه يقتصر على الغشاء ويسمح له بالارتباط والقنوات الأيونية في الغشاء بعد المشبكي. وبالتالي، يمكن أن تلعب بالميتويلات دورا في تنظيم إطلاق الناقل العصبي.[4]
يتوسط بالميتويليشن تقارب البروتين لطوافات الدهون ويسهل تجميع البروتينات.[4] يمكن أن يزيد التجميع من قرب جزيئين. بدلاً من ذلك، يمكن للتجميع عزل البروتين بعيدا عن الركيزة. على سبيل المثال، بالميتويليشن من فوسفوليباز د يعزل الأنزيم بعيداً عن فوسفاتيديل كولين الركيزة لها. عندما تنخفض مستويات الكوليسترول أو تزيد مستويات PIP2 يتم تعطيل التوطين بوساطة البالميتات، ينتقل الإنزيم إلى PIP2 حيث يواجه ركائزه ويكون نشطاً عن طريق عرض الركيزة.[4][4][4]
بروتينات GPI
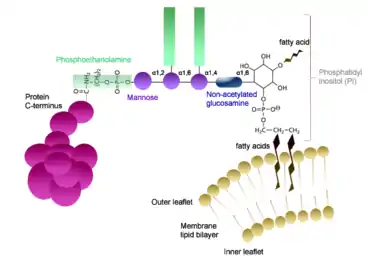
ترتبط البروتينات المرتبطة بالجليكوزيل فوسفاتيلينوسيتول (البروتينات المرتبطة - GPI) بمجموعة جزيئية معقدة من GPI عبر ارتباط أميد بمجموعة الكربوكسيل الطرفية للبروتين.[4] يتكون مركب GPI من عدة مكونات رئيسية مترابطة جميعها: فسفويثانولامين، رباعي السكاريد الخطي (يتكون من ثلاثة مانوز وجلوكوز امينيل) وفوسفاتيديل ينوسيتول.[4] ترتبط مجموعة فوسفاتيديل ينوسيتول برابطة غليكوسيدية مع الجلوكوزامين غير رباعي السكاريد. بعدها تتكون رابطة فوسفوديستر بين المانوز في الطرف غير المختزل (من رباعي السكاريد) والفوسفويثانولامين. ثم يتم ربط فوسفويثانولامين برابطة أميدية بالطرف C لمجموعة الكربوكسيل للبروتين المعني. يحدث ارتباط GPI من خلال عمل مركب GPI- ترانساميداز. يتم إدخال سلاسل الأحماض الدهنية للفوسفاتيديل ينوسيتول في الغشاء، وبالتالي فهي ما يثبت البروتين في الغشاء.[4] توجد هذه البروتينات فقط على السطح الخارجي لغشاء البلازما.
الأدوار والوظيفة
تختلف بقايا السكر في رباعي السكاريد وبقايا الأحماض الدهنية في مجموعة فوسفاتيديلينوسيتول اعتمادًا على البروتين. هذا التنوع الكبير هو ما يسمح لبروتينات GPI بأن يكون لها نطاق واسع من الوظائف بما في ذلك العمل كأنزيمات محللة للماء، وجزيء الالتصاق، ومستقبلات، ومثبط للبروتياز، وبروتينات تنظيمية مكملة.[4] علاوة على ذلك، تلعب بروتينات GPI دورًا مهمًا في تكوين الجنين، والتطور وتكوين الخلايا العصبية وجهاز المناعة والإخصاب. وبشكل أكثر تحديدًا، بروتين GPI IZUMO1R/JUNO (سمي على اسم إلهة الخصوبة الرومانية) موجود على بلازما البيض لها دور أساسي في اندماج البويضة والحيوانات المنوية. إن إطلاق بروتين IZUMO1R /JUNO GPI من غشاء بلازما البيض لا يسمح للحيوانات المنوية بالاندماج مع البويضة ويقترح أن هذه الآلية قد تساهم في كتلة تعدد النطاف في غشاء البلازما في البيض.[4] الأدوار الأخرى التي يسمح بها تعديل GPI هي الارتباط بالمجالات الدقيقة الغشائية أو التماثل المتجانس العابر أو الفرز القمي في الخلايا المستقطبة.
مراجع
- Gerald Karp (2009)، Cell and Molecular Biology: Concepts and Experiments، John Wiley and Sons، ص. 128–، ISBN 978-0-470-48337-4، مؤرشف من الأصل في 17 أغسطس 2021، اطلع عليه بتاريخ 13 نوفمبر 2010.
- Voet D, Voet JG, Pratt CW (2013)، Fundamentals of Biochemistry: Life at the Molecular Level (ط. 4th)، John Wiley & Sons, Inc.، ص. 263، ISBN 978-0470-54784-7.
- "Lipid anchors on membrane proteins."، Current Opinion in Structural Biology، 1 (4): 522–9، أغسطس 1991، doi:10.1016/s0959-440x(05)80072-7.
- "Thematic review series: lipid posttranslational modifications. Structural biology of protein farnesyltransferase and geranylgeranyltransferase type I"، Journal of Lipid Research، 47 (4): 681–99، أبريل 2006، doi:10.1194/jlr.R600002-JLR200، PMID 16477080.
 بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي