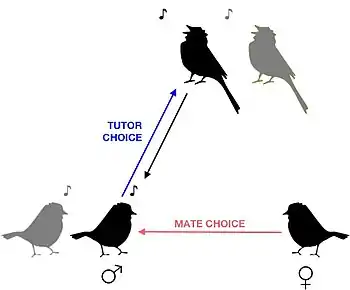
تفضيل نمط محدد من النغمات
تفضيل نمط محدد من النغمات هو قدرة تحتاجها الطيور المغردة لتمييز النغمات الخاصة بالطيور المماثلة لها جنسيًا عن النغمات الخاصة بالطيور المغايرة جنسيًا من أجل أن تختار الإناث شريكها المناسب، وأيضًا من أجل أن تتعلم الذكور اختيار مدرّس نغمات مناسب أثناء تعلمها الأصوات. اكتشف الباحثون الذين يدرسون (العصفور الدوري) أنَّ الطيور الصغيرة تمتلك هذه القدرات منذ ولادتها، لأن الذكور الصغار الذين ينشؤون في عزلة ويتعلمون من تسجيلات صوتية يختارون تعلم الأغاني التي تحتوي على مقاطع صوتية خاصة بأنواعهم فقط.[1] تشير الدراسات التي أُجريت على مراحل مختلفة من الحياة إلى أنَّ تطوير وتعزيز خاصية تفضيل نمط محدد من النغمات يتم خلال عملية التطور كوظيفة اجتماعية.[2]
الآليات العصبية
لم توصف الآلية المسؤولة عن القدرة على التمييز بين أنواع النغمات بشكل كامل من قبل الباحثين في مجال علم الأعصاب، ولكن ثبت احتواء ما لا يقل عن خمسة هياكل مختلفة ضمن المسار السمعي على عصبونات تستجيب بشكل تفضيلي لنغمات محددة. يتسبب هيكل الشبكات العصبية ونمط تشكلها والمستقبلات والقنوات الأيونية في استجابة بعض الخلايا العصبية إلى الحد الأقصى لتردد محفز معين أو طوره أو سعته أو نمطه الزمني ويعرف ذلك باسم تناغم الطيف الزمني.[3] يوفر تناغم الطيف الزمني في المسار السمعي للجهاز العصبي المركزي لطيور الدوري القدرة على التمييز بين النغمات الخاصة بجنسها والأخرى الخاصة بالجنس المغاير. توجد أفضل خصائص للخلايا العصبية السمعية في عصفور زيبرا والكناري والزرزور الأوروبي والبومة المصاصة.
مسارات التعلم والنطق
يتكون نظام النغمات التقليدي للطيور المغردة من جزأين: المسار الأمامي في الدماغ الأمامي المسؤول عن تعلم النغمات، والمسار الخلفي في الدماغ الأمامي أو (مسار مولّد النغمات) المسؤول عن إنتاج النغمات (نطقها). يحتوي كلا المسارين على عصبونات تستجيب لنغمات محددة.[4] فقدت أنثى الكناري القدرة على التمييز بين النغمات الخاصة بنوعها الجنسي والنغمات الخاصة بالنوع الجنسي المغاير لها بعد الضرر الذي لحق بالمركز الصوتي في الدماغ (HVC) وهو نواة تقع في قمة المسارين السابقين.[5] ومع ذلك تستجيب معظم الخلايا العصبية في معظم الذكور إلى الحد الأقصى لصوت نغمات الطائر ذاته، حتى أكثر من استجابته لنغمات المعلم أو أيّ نغمة أخرى.[6][7] تتحول الخلايا العصبية في المركز الصوتي في الدماغ من الاستجابة القصوى لنغمات المعلم (35-69 يومًا بعد خروج الطائر من البيضة) إلى الاستجابة بشكل أفضل لأغنية الطائر الذاتية (بعد 70 يومًا من خروج الطائر من البيضة).[8]
قوقعة الأذن
درست الخلايا الشعرية للطيور على نطاق واسع في قوقعة البومة البيضاء،[9] ومن المعروف الآن أنَّ كلًا من التركيب الشكلي لحليمات الخلايا الشعرية والقنوات الأيونية التي تميز أغشية الخلايا الشعرية هي التي تمنح خصائص ضبط طبقات النغمة.
المهاد السمعي
أظهرت الدراسات التي أجريت على عصفور زيبرا أنَّ النواة الموجودة في المهاد السمعي (على بعد خطوتين من القوقعة) لا تنقل المدخلات الصوتية من الهياكل الحسية المحيطية إلى مناطق الدماغ الأمامي العلوية. تُظهر نوى المهاد أنماطًا مختلفة من التعبير الجيني كاستجابةً لمحفزات مختلفة، وتدخلهم في عملية التمييز الصوتي.[10] تحتوي الخلايا العصبية الموجودة في النواة البيضوية على حقول استقبال مخصصة للاستجابة لمزيج محدد من الأطياف الصوتية والزمنية الموجودة في اجزاء معينة من النغمة.
أهمية التطور
تعتبر الآليات العصبية المسؤولة عن تفضيل نغمات محددة والموجودة في المسار السمعي مهمة للتعرف على الأنواع الجنسية في سياق التطور. تلعب ميزات التعرف على النوع الجنسي دورًا رئيسيًا في كل من أصل الانعزال التكاثري والحفاظ عليه. وبالإضافة إلى ذلك فإن وجود آلية عصبية مشتركة لتفضيل نغمات محددة له آثار على تطور نغمات الذكور وتفضيلات الأنثى، مما قد يساعد في تفسير التنوع الهائل للأنماط الظاهرة للنغمات في الطيور المغردة الموجودة حاليًا.
المراجع
- Marler, P. and S. Peters. 1977. Selective vocal learning in a sparrow. Science 198:519-521.
- Amin, N., A. Doupe, and F. Theunissen. 2007. Development of selectivity for natural sounds in the songbird auditory forebrain. Journal of Neurophysiology 97:3517.
- Woolley, A.M.N., T.E. Fremouw, A. Hsu and F.E. Theunissen. 2005. Tuning for spectro-temporal modulations as a mechanism for auditory discrimination of natural sounds. Nature Neuroscience 8:1371-1379.
- Solis, M.M., M.S. Brainard, N.A. Hessler and A.J. Doupe. 2000. Song selectivity and sensorimotor signals in vocal learning and production. Proceedings of the National Academy of Sciences USA 97:11836-11842.
- Brenowitz, E.A. 1991. Altered perception of species-specific song by female birds after lesions of a forebrain nucleus. Science 251:303-304.
- Margoliash, D., and M. Konishi. 1985. Auditory representation of autogenous song in the song system of white-crowned sparrows. Proceedings of the National Academy of Sciences USA 82:5997-6000.
- Margoliash, D. 1986. Preference for autogenous song by auditory neurons in a song system nucleus of the white-crowned sparrow. Journal of Neuroscience 13:4737-4747.
- Nick, T.A., and M. Konishi. 2005. Neural song preference during vocal learning in the zebra finch depends on age and state. Journal of Neurobiology 62:231-242.
- Konishi, M., T.T. Takahashi, H. Wagner, W.E. Sullivan and C.E. Carr. 1988. Neurophysiological and anatomical substrates of sound localization in the owl. In “Auditory Function”. G.M. Edelman, W.E. Gall and W.M. Cowan, Eds. Wiley: New York.
- Brauth, S.E., W. Liang and W.S. Hall. 2006. Contact-call driven and tone-driven zenk expression in the nucleus ovoidalis of the budgerigar (Melopsittacus undulates). NeuroReport 17:1407-1410.

 بوابة علم الأحياء التطوري
بوابة علم الأحياء التطوري بوابة طيور
بوابة طيور