مدخل إلى التطور
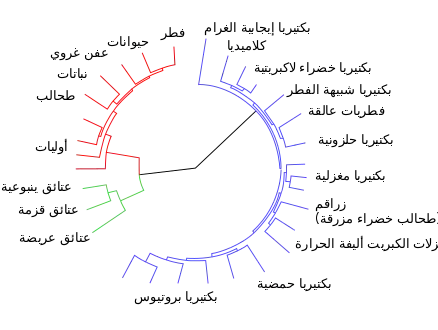
التطور في علم الأحياء هو عملية التغير على مر الزمان في خلة موروثة أو أكثر في تجمعات الأفراد.[1] يمكن أن يؤدي التطور في النهاية إلى تغيير كافة مواصفات النوع قيد التطور مما يؤدي إلى نشوء نوع جديد من الكائنات الحية. مصطلح التطور العضوي organic evolution أو التطور البيولوجي يستخدم غالبا لتفريق هذا المصطلح عن استعمالات أخرى.
| جزء من سلسلة مقالات حول |
| التطور |
|---|
 |
| بوابة علم الأحياء التطوري |
بدأ تطور نظرية التطور الحديثة بإدخال مصطلح الاصطفاء الطبيعي في مقالة مشتركة لتشارلز داروين وألفرد راسل والاس. من ثم حققت النظرية شعبية واسعة بعد الإقبال على قراءة كتاب داروين أصل الأنواع. كانت نظرية داروين ووالاس الأساسية أن التطور يحدث وفق خلة قابلة للتوريث تؤدي إلى زيادة فرصة بعض الأفراد الحاملين لهذه الخلة trait بالتكاثر أكثر من الأفراد الذين لا يحملونها. هذه النظرية كانت جديدة تماما ومخالفة لمعظم أسس النظريات التطورية القديمة، خصوصاً النظرية المطورة من قبل جان باتيست لامارك.
حسب نظرية داروين ووالاس: يحدث التطور نتيجة تغير أو طفرة في خلات قابلة للتوريث ضمن مجموعة حيوية على امتداد أجيال متعاقبة، كما يحدده التغيرات في التواترات الأليلية للجينات. ومع الوقت، يمكن أن تنتج هذه العملية ما نسميه انتواعاً، أي تطور نوع جديد من الأحياء بدءاً من نوع موجود أساسا. بالنسبة لهذه النظرية فإن جميع المتعضيات الموجودة ترتبط ببعضها البعض من خلال سلف مشترك، كنتيجة لتراكمات التغيرات التطورية عبر ملايين السنين.
التطور هو أيضاً مصدر للتنوع الحيوي على كوكب الأرض، بما فيها الأنواع المنقرضة المسجلة ضمن السجل الأحفوري أو المستحاثي. [2][3] الآلية الأساسية التي ينتج بها التغير التطوري هي ما تدعوه النظرية: الاصطفاء الطبيعي (بالإنجليزية: natural selection) (الذي يتضمن البيئي والجنسي والقرابة مع الانحراف الوراثي). تقوم هاتان العمليتان أو الآليتان بالتأثير على التنوع الجيني المتشكل عن طريق الطفرات، والتأشيب الجيني وانسياب المورثات. لذا يعتبر الاصطفاء الطبيعي عملية يتم بها بقاء ونجاة الأفراد ذوي الخلات الأفضل (للحياة) وبالتالي التكاثر. بعبارة أخرى: الاصطفاء يحابي أصحاب الصلاحية الأكبر. إذا كانت هذه الخلات قابلة للتوريث فإنها ستنتقل إلى الأجيال اللاحقة، مما يجعل الخلات الأكثر نفعاً وصلاحية للبقاء أكثر شيوعاً في الأجيال اللاحقة. [1][4][5] فبإعطاء وقت كاف، يمكن أن تنتج هذه العملية العفوية تلاؤمات متنوعة نحو تغيرات الشروط البيئية [6]
الفهم الحديث للتطور يعتمد على نظرية الاصطفاء الطبيعي، التي وضعت أسسها أساساً في ورقة مفصلية عام 1858 من قبل تشارلز داروين وألفرد راسل والاس ونشرت ضمن كتاب داروين الشهير أصل الأنواع. في الثلاثينات من القرن العشرين، ترافق الاصطفاء الطبيعي الدارويني مع نظرية الوراثة المندلية لتشكل ما يدعى الاصطناع التطوري الحديث، وعرفت أيضاً بالداروينية-الجديدة. الاصطناع الحديث يصف التطور كتغير في تواتر الأليلات ضمن مجموعة حيوية من جيل إلى الجيل الذي يليه. [6] هذه النظرية سرعان ما أصبحت المبدأ المركزي المنظم للحياوة الحديثة، نسبة لقدرتها التفسيرية والتنبؤية العالية، تربط حالياً بشكل مباشر مع دراسة أصل مقاومة المضادات الحيوية في الجراثيم، الاجتماعية العليا (بالإنجليزية: eusociality) في الحشرات، والتنوع في النظام البيئي (بالإنجليزية: ecosystem) للأرض. مع أن هناك إجماع علمي لدعم صلاحية وصحة نظرية التطور [7]، لتطبيقاتها وقدرتها التفسيرية والتنبؤية لأصول الأجناس والأنواع الحية، فإن هذه النظرية تبقى في قلب جدالات دينية واجتماعية حول مفاهيمها ومدى صحتها بسبب صدامها مع بعض الرؤى حول نظرية الخلق في بعض الديانات.
دراسة التطور
تاريخ الفكر التطوري
تعود فكرة التطور البيولوجي إلى عهود قديمة فبعض الفلاسفة الإغريق كانوا يؤمنون بهذه الفكرة مثل أناكسيماندر وأبيقور إضافة لبعض فلاسفة الهند مثل باتانجالي. كما أن بعض من عاشوا في العالم الإسلامي في القرن الثالث والرابع مثل الجاحظ والمسعودي كانوا يعتقدون بهذه النظرية[بحاجة لمصدر]. ولكن في القرنين الثامن عشر والتاسع عشر أعاد جان-بابيست لامارك وتشارلز داروين إحياء هذه النظرية القديمة مع بعض التعديلات. [8] الآلية التي يحدث بها التغير التطوري من نوع لآخر: ألا وهي الاصطفاء الطبيعي. بعد الكثير من العمل على نظريته قام داروين بنشر عمله عن التطور بعد تسلمه رسالة من ألفرد راسل والاس يكشف له والاس فيها عن اكتشافه الشخصي حول موضوع الاصطفاء الطبيعي. لهذا ينسب لوالاس دور مشارك في التأسيس لهذه النظرية. [9] أثار نشر كتاب داروين قدراً كبيراً من الجدل العلمي والاجتماعي. فبالرغم من أن حدوث تطور بيولوجي من نوع ما أصبح مقبولاً من قبل عدد كبير من العلماء، فإن أفكار داروين خاصة حول حدوث تطور تدريجي من خلال الاصطفاء الطبيعي تمت مهاجمتها ونقدها بقوة. إضافة لذلك كان داروين قادراً على تبيان الاختلاف بين النواع مفسراً إياه بالاصطفاء الطبيعي ثم التلاؤم، إلا انه كان عاجزاً عن تفسير كيفية نشوء الاختلاف أو كيف يتم تعديل النواع عبر الأجيال، كان لا بد من انتظار نشوء علم الوراثة على يد ماندل. [10]
استطاع غريغور مندل بالعمل على وراثة النبات من كشف حقيقة انتقال ميزات معينة في حبات البازلاء، هذا الانتقال يحدث بأشكال متنوعة وكانت قابلة للتوريث بنسب واضحة قابلة للتنبؤ. [11] أعيد إحياء عمل ماتدل في عام 1901، وفسر بداية على أنه دعم «للقفزة» المعاكسة للداروينية، أو ما يدعى بمدرسة القفز التطوري saltationist ومعاكسة لفكرة التدرجية. [12]
الاصطفاء الطبيعي


كانت المتاحف ومقتنيات التاريخ الطبيعي من التسليات الشائعة في القرن التاسع عشر. في التوسع الأوروبي والبعثات البحرية وُظف خبراء تاريخ طبيعي وأمناء متاحف ضخمة تعرض عينات محفوظة وأخرى حية من التنوعات الحيوية. تشارلز داروين كان خريج إنجليزي تعلم وتدرب في مجالات علم التاريخ الطبيعي. والمؤرخون الطبيعيون كانوا يجمعون مقتنيات العينات هذه ويفهرسونها ويصفونها ويدرسونها. وقد وُظف تشارلز داروين خبير تاريخ طبيعي على متن سفينة بيغل، التي عينت للقيام ببعثة حول العالم لمدة خمس سنوات. وقد قام داروين خلال رحلته بمراقبة الكثير من الكائنات الحية وتجميعها لكونه مهتماً جداً بالأشكال المتنوعة للحياة على طول سواحل أمريكا الجنوبية وجزر غالاباغوس المجاورة.[13][14]
لقد حصَّل داروين خبرة واسعة مع تجميعه ودراسته للتاريخ الطبيعي للكائنات الحية من الأماكن البعيدة. ومن خلال دراسته، صاغ فكرة أنَّ كل نوع كان قد تطور من أسلاف ذوي ميزات مشابهة. وفي عام 1838، شرح كيف تؤدي إلى ذلك عملية سماها الاصطفاء الطبيعي.[15]
حجم أي تجمع يتعلق بعدد ومدى توفر الموارد التي بإمكانها دعمه. من الضروري توفر تكافؤ أو توازن بين حجم التجمع والموارد المتاحة كي يتبت التجمع على نفس الحجم على مر السنوات. وبما أنَّ الكائنات الحية تنتج عدداً من النسل أكبر من أن تقدر البيئة على إعالته، فلن يستطيع كل الأفراد من كل جيل النجاة. لا بد من أن يكون هناك صراع تنافسي من أجل الموارد التي تساعد على البقاء. كنتيجة لذلك، استنتج داروين أنَّه لم تكن الصدفة وحدها التي تحدد فرص البقاء. بل إنَّ بقاء الكائن الحي يتعلق بالاختلافات (أي الخلات) التي توجد في كل كائن حي، والتي تعزز أو تضعضع من بقائه وتكاثره. يكون من المرجح أكثر أن ينتج الأفراد المتكيفين جيداً نسلاً أكبر عدداً من منافسيهم الأقل تكيفاً. وهكذا تختفي الخلات التي تعيق البقاء والتكاثر مع مرور الأجيال. الخلات التي تساعد الكائن الحي على البقاء والتكاثر تتراكم عبر الأجيال. أدرك داروين أنَّ هذا التفاوت بالقدرات بين الأفراد قد يسبب تغيرات تدريجية في التجمع، واستعان بمصطلح الاصطفاء الطبيعي لوصف هذه السيرورة.[16][17]
شكلت مراقبات التنوعات في الحيوانات والنبانات الأساس الذي تقوم عليه نظرية الاصطفاء الطبيعي. على سبيل المثال، لاحظ داروين أنه توجد بين السحالب (الأوركيد) والحشرات علاقة وثيقة تتيح تلقيح النبات. وقد لاحظ أنَّ السحالب لديها تشكيلة من البنيات التي تجذب الحشرات، بحيث تلتصق حبوب اللقاح من الزهور بأجساد الحشرات. وبهذه الطريقة، تنقل الحشرات حبوب اللقاح من السحلب الذكر إلى الأنثى. مع أنَّ مظاهر السحالب مفصلة ودقيقة، إلا أنَّ هذه الأجزاء المتخصصة فيها تصنع من نفس البنيات الأساسية التي تشكل الزهور الأخرى. وضع داروين في كتابه تلقيح السحالب الافتراض بأنَّه تم تكييف زهور السحالب من أجزاء موجودة سابقاً بواسطة الاصطفاء الطبيعي.[18]
كان داروين ما زال يدرس أفكاره ويختبرها حين تلقى رسالة من ألفرد والاس يشرح فيها نظرية تشبه كثيراً النظرية الخاصة بداروين. هذا أدى لنشر مشترك فوري لكلتا النظريتين. رأى كلاهما تاريخ الحياة كأنه شجرة عائلة، بحيث كل تفرع من أغصان الشجرة يمثل سلفاً مشتركاً. أطراف الأغصان مثلت الأنواع الحديثة، والتفرعات مثلت الأسلاف المشتركة التي تتحدر منها أنواع مختلفة. لتفسير هذه العلاقات، قال داروين أنَّ كل الكائنات الحية كانوا أقرباء، وهذا يعني أن الحياة لا بد من أنها تحدرت من بضعة أشكال، أو حتى من سلف مشترك واحد. وقد سمى هذه السيرورة التحدر مع تعديلات.[19]
نشر داروين نظريته للتطور بواسطة الاصطفاء الطبيعي في أصل الأنواع عام 1859. نظريته تعني أنَّ كل أشكال الحياة، بما فيها البشرية، هي نتيجة السيرورات الطبيعية المستمرة. وهذا التضمين بأن كل الحياة على الأرض تعود لسلف مشترك لقي معارضة من قبل بعض الجماعات الدينية. واعتراضاتهم هي على النقيض من نسبة الدعم العلمي للنظرية الذي يصل إلى أكثر من 99 بالمئة.
كثيراً ما يُستعمل مصطلح البقاء للأصلح كمرادف للاصطفاء الطبيعي ويُساوى معه. ولكن المصطلح البقاء للأصلح نشأ في كتاب هربرت سبنسر مباديء علم الأحياء عام 1864، أي بعد خمس سنوات من نشر داروين لأعماله الأصلية. بقاء الأصلح يشير يصف عملية الاصطفاء الطبيعي بصورة خاطئة. وذلك لأن الاصطفاء الطبيعي لا يتعلق بالبقاء فقط، وليس من الضروري أنَّ الأصلح هو من يبقى دائماً.[20]
مصدر التنوع
نظرية الاصطفاء الطبيعي لداروين وضعت الأساس للنظرية التطورية الحديثة، وتجاربه وملاحظاته أظهرت أنَّ الكائنات الحية في التجمعات تباينوا فيما بينهم، وأنَّ بعض هذه الاختلافات كانت تُوَرَّث، وأنَّ هذه الاختلافات يمكن أن تخضع للاصطفاء الطبيعي. ولكنه لم يقدر على تفسير مصدر هذه الاختلافات. ومثل الكثير ممن سبقوه، أخطأ داروين بالاعتقاد أنَّ الخلات القابلة للتوريث هي ناتجة عن الاستخدام والإهمال، وأنَّ الميزات التي يكتسبها الكائن الحي في حياته يمكن أن تُوَرث إلى نسله. وفد بحث عن أمثلة، مثل أنَّ الطيور الكبيرة التي تتغذى على الأرض تصبح أرجلها أقوى من خلال التمرين وتصبح أجنحتها أضعف كنتيجة لعدم الطيران، إلى أن أصبحت غير قادرة على الطيران على الإطلاق، مثل النعامة.[21] سُمي الفهم الخاطئ هذا بوراثة الخواص المكتسبة، وكان جزءاً من نظرية تطافر الأنواع transmutation of species التي عرضها جان باتيست لامارك عام 1809. وفي أواخر القرن التاسع عشر أصبحت تُعرف هذه النظرية بال لاماركية. قام داروين بوضع نظرية غير ناجحة سماها شمولية التخلق pangenesis في سبيل محاولة تفسير كيف يمكن أن تورث الخواص المكتسبة. في آخر سنوات الثمانين من القرن الثامن عشر، أشارت تجارب أوغست وايزمن إلى أن التغيرات الناجمة عن الاستخدام وعدمه لا يمكن توريثها، وفقدت اللاماركية التأييد تدريجياً.[22]
قامت أعمال غريغور مندل الرائدة في علم الوراثة بتزويد المعلومات الناقصة الضرورية لتفسير كيف يتم توريث الميزات الجديدة من جيل الآباء إلى جيل الأبناء. أظهرت تجارب مندل على عدة أجيال من البازلاء أنَّ الوراثة تعمل عن طريق تفرقة المعلومات الوراثية وإعادة خلطها خلال تكوين الخلايا الجنسية، ومن ثم تأشيب تلك المعلومات خلال التلقيح. هذا الأمر يشابه خلط مجموعة من أوراق اللعب المختلفة، بحيث يحصل الكائن الحي على نصف الأوراق المخلوطة عشوائياً من كل من الأبوين. قام مندل بتسمية المعلومات الوراثية بالعوامل factors، ولكنها أصبحت تُعرف بالجينات فيما بعد. الجينات هي وحدات الوراثة الأساسية في الكائنات الحية. وهي تحتوي على المعلومات التي توجه نمو الكائنات الحية وسلوكياتها.
الجينات هي مصنوعة من الدنا. الدنا هو جزيء طويل يتكون من جزيئات تسمى نوكليوتيدات. المعلومات الوراثية هي مشفرة في تسلسل النوكليوتيدات الذي يشكل الدنا، مثلما يحمل تسلسل أحرف الكلمات معنى الجملة. الجينات هي بمثابة تعليمات قصيرة تتكون من «أحرف» أبجدية الدنا. المجموعة الكاملة للجينات مُجتمعة تحمل تعليمات كافية لتكون بمثابة «كتيب تعليمات» لبناء الكائن الحي وتشغيله. ولكن من الممكن أن تتغير هذه التعليمات التي تحددها الأبجدية الدناوية من خلال الطفرات، وهي تغيرات تطرأ على التعليمات الوراثية. الجينات في الخلية تنتظم في كروموسومات، وهي رزم تحمل الدنا. إنَّ ما يسبب تكوين توليفات جديدة من الجينات في النسل هو إعادة خلط الكروموسومات. بما أنَّ الجينات تتفاعل مع بعضها البعض خلال نمو الكائن الحي، فإن التوليفات الجديدة للجينات الناتجة عن التكاثر الجنسي قد تزيد من التنوع الجيني في التجمع حتى بدون حدوث طفرات.[23] يمكن أن يزداد التنوع الجيني في التجمع أيضاً عندما يتزاوج أفراد هذا التجمع مع أفراد من تجمعات أخرى، مما يؤدي لانسياب الجينات فيما بين التجمعات. وهذا قد يدخل إلى التجمع جينات لم تكن موجودة به سابقاً.[24]
التطور ليس عملية عشوائية. مع أنَّ الطفرات التي تحدث في الدنا هي عشوائية، إلا أنَّ الاصطفاء الطبيعي ليس بعملية تقوم على الصدفة: البيئة تحدد احتمال النجاح التناسلي. التطور هو نتيجة محتومة لعملية النسخ التي تحدث بها أخطاء، ولتكاثر الكائنات الحية ذاتية التناسخ عبر بلايين السنين في ظل ضغوط البيئة الاصطفائية. نتيجة التطور ليست كائناً حياً كامل التصميم. نتائج الاصطفاء الطبيعي النهائية هي كائنات حية متكيفة على بيئتها الحالية. الاصطفاء الطبيعي ليس بتقدم نحو هدف أو غرض نهائي. التطور لا يسعى لإنتاج أشكال حياة متقدمة أو عاقلة أو معقدة أكثر.[25] على سبيل المثال، البراغيث (طفيليات لا أجنحة لها) تحدرت من طويلات الأجنحة السلفية. والأفاعي هي سحالي لم تعد بحاجة إلى أطراف - مع أنَّ الأصلات (جنس من الثعابين) ما زالت تنمو لديها بنيات صغيرة جداً من بقايا سيقان سلفها الخلفية.[26][27] الكائنات الحية هي ليست إلا نتيجة للاختلافات التي تنجح أو تفشل، والأمر يعتمد على الظروف البيئية في حينها.
التغيرات البيئية السريعة عادة تسبب الانقراضات.[28] ونسبة الأنواع المنقرضة من كل الأنواع التي عاشت على الأرض هي 99.9 بالمئة.[29] منذ بدء الحياة على الأرض، أدت خمسة انقراضات جماعية رئيسية لانخفاض فجائي في التنوع النوعي. وأحدثها كان انقراض العصر الطباشيري-الثلاثي الذي حدث قبل 65 مليون سنة.[30]
إثباتات التطور
تسلسل من التكيفات:
- سمكة باندر تناسب المياة الضحلة الموحلة
- تيكتاليك بزعانف تشبه الأطراف يمكن أن تحملها إلى اليابسة
- فقاريات رباعية الأطراف بالمستنقعات المليئة بالحشائش مثل:
- اكانسوستيجا والذي كان لديه أرجل بثمان أصابع
- اكستيوستيجا بأطراف
شمل الأحفاد أيضا لحميات زعانف مثل سيلاكانث]] خلف التطور وراءه العديد من السجلات التي تروي تاريخ الأنواع المختلفة وزمن نشوئها. الأحافير بمجموعها مع التشريح المقارن للنباتات والحيوانات الموجودة حاليا، تشكل سجلا تشريحيا ومورفولوجيا. وبالمقارنة التشريحية والشكلية بين الأنواع الحالية والأنواع المنقرضة يمكن لعلماء المستحاثات ان يقوموا بمعرفة الارتباطات والأصول المشتركة بين هذه الأنواع. تقوم بعض المستحاثات المهمة بإثبات الصلة بين أنواع منقرضة وأنواع موجودة حاليا عن طريق ما يدعى أنواع "انتقالية"، مثال هذه الأنواع الانتقالية أرخأيوبتركس الذي أثبت العلاقة بين الديناصورات والطيور. [31][32]
لاحقاً سمح تطور علم الوراثة الجزيئي وخصوصاً إمكانية سلسلة الدنا DNA، للبيولوجيين بدراسة سجل التطور عن طريق البنى الوراثية للمتعضيات الحالية والمنقرضة، مما وسع بشكل كبير وعدل من إمكانية إيجاد الصلات والقرابات بين الأنواع وأحيانا كان يؤدي لتعديلات جذرية في التصنيف الحيوي للأحياء. عن طريق التشابه والاختلاف بين تسلسلات الدنا DNA للمتعضيات الحية يقوم علماء الحياوة حاليا بإيجاد وتعديل العلاقات وصلات القرابة بين الأنواع الحية قديمها وجديدها. فعن طريق الدراسات الوراثية تبين أن 95% من الصيغة الوراثية (الجينية) متشابهة بين الإنسان والبعام. [33][34]
تم جمع إثباتات أخرى من بعض البنى التشريحية الموجودة في بعض الكائنات الحية كما عند الباندا أو شكل الأقدام عند السحالي أو انعدام العيون عند الأسماك الكهفية، مما قدم إثباتات لإمكانية التنامي التطوري. تقدم دراسات أخرى إثباتات عن طريق تمثيل صلات تطورية تتضمن التوزع الجغرافي للأنواع. مثلا: أحاديات الفتحة Monotreme، مثل platypus ومعظم الجرابيات marsupials مثل الكنغر والكوالا وجدوا فقط في أستراليا وضحوا أن سلفهم المشترك مع الثدييات المشيمية placental mammals عاش قبل غمر الجسر الأرضي القديم بين أستراليا وآسيا.
جميع هذه الإثباتات من علم الإحاثة، التشريح، علم الوراثة، والجغرافيا، إضافة لمعلومات أخرى حول تاريخ الأرض قام العلماء بربطها سوية ضمن إطار تقدم نظرية التطور من خلاله وتجعلها نظرية علمية متماسكة. فمثلا علم المناخ الإحاثي paleoclimatology يشير إلى العصر الجليدي الدوري الذي كان فيه مناخ الأرض أكثر برودة، مما أدى لنشوء وانتشار أنواع حية قادرة على تحمل البرد القارس ومن أهم هذه الأنواع الماموث ماموث صوفي.
الإثباتات المورفولوجية

غالبا ما تعتبر المستحاثات إثباتات حرجة لتقييم عمق الصلات بين الأنواع. بما أن استحاثة متعضية يعتبر أمرا نادر الحدوث، يحدث غالبا بوجود أجزاء صلبة مثل الأسنان والعظام، لذلك فإن المستحاثات غالبا ما تعتبر بأنها تقدم معلومات ضئيلة أو متوسطة الأهمية حول علاقات القرابة بين الأنواع. مع هذا فيمكن حدوث استحاثة لبعض النواع بدون أجزاء صلبة (أسنان، عظام) في بعض الظروف: rapid burial، بيئات منخفضة الأكسجين، أو تأثير ميكروبيولوجي. [36] تؤمن السجلات الأحفورية أنماطا متعددة من البيانات المهمة لدراسة التطور. أولا يحوي السجل الأحفوري الأمثلة المعروفة المبكرة للحياة، إضافة لحدوث أولى حالات القرابة الفردية بين الأنواع. فمثلا حسب السجل الأحفوري ظهر أول حيوانات معقدة في عصر الكمبري المبكر، أي حوالي 520 مليون سنة مضت. ثانيا سجلات الأنواع المنفردة تعطي معلومات بخصوص الأنماط والأشكال التي مر بها النوع وسرعة التغير والتطور في هذا النوع. مظهرة مثلا فيما إذا تطور هذا النوع إلى نوع جديد مختلف (في عملية ندعوها الانتواع speciation) تدريجيا وبشكل متزايد، أو خلال قترات زمنية قصيرة نسبيا ضمن الزمن الأراضي (الجيولوجي). ثالثا يعتبر السجل الأحفوري وثيقة للأنماط الكبيرة الانتشار والأحداث المهمة في تاريخ الحياة، والعديد من هذه المستحاثات قد أثرت بشكل فعلي في تصورنا للتاريخ التطوري لعلاقات القربى. مثلا الانقراض الكبير نتج في عدة أزمنة مؤديا لفقدان مجموعات كاملة من الأنواع، مثل الديناصورات غير الطيارة non-avian dinosaurs، في حين أنها لا تؤثر على مجموعات أخرى من الأنواع. مؤخرا استطاع علماء الحياوة الجزيئية أن يستعملوا الزمن منذ تقارب الأنساب lineages المختلفة ليقوموا بمعايرة سرعة أو معدل التراكم التطافري، حساب أزمنة تطور مجينات الأنساب المختلفة.
علم الوراثة العرقي كعلم يدرس أصل الأنواع وأسلافها، قدم وكشف أن البنى ذات التنظيم الداخلي المتشابه يمكن أن تقوم بوظائف متقاربة. أقدام الفقاريات تعتبر مثالا شائعا لهذه البنى المتشابهة.
علم المناعة
يعتمد هذا الدليل على تحديد درجة القرابة بين الأنواع المختلفة وفق التفاعلات المناعية التي تحدث بين مولد الضد(المستضد) والضد الذي يشكله جسم الكائن الحي عند دخول مادة غريبة فيه؛ نتيجة لهذا التفاعل المناعي (ضد-مستضد) يحدث ترسيب للمواد المتفاعلة ويتشكل معقد مجرد من السمية.
مثال: عند حقن بروتينات مأخوذة من جسم الإنسان في جسم خنزير الهند يتولد في دمه أضداد (وهذه الأضداد قادرة على حض مولدات الضد عند الإنسان على التفاعل المناعي معها)؛ عند أخذ الأضداد السابقة وحقنها في بلاسما دم القرد تحرض عنده ارتصاصا (أو ترسيبا) نسبته 50% ويفسر هذا الترسيب وجود صلة تشابه بين القردة والبشر بنسبة متوسطة؛ مثل هذا الترسيب لا يحصل عند حقن الأضداد في دم الجرابيات أو الطيور لعدم وجود التشابه بينها وبين الإنسان.
أسلاف الكائنات الحية
في علم الأحياء، تفترض نظرية السلف المشترك العام universal common descent أن جميع الأحياء الموجودة على سطح الأرض تنحدر من سلف مشترك وحيد عام أو لنقل حوض جيني أصلي وحيد. تستند نظرية السلف المشترك إلى وجود سمات مشتركة بين كافة المتعضيات الحية. في أيام داروين كانت الاثباتات على التشارك بالسمات يستند فقط إلى الملاحظات المرئية للتشابهات الشكلية، مثل حقيقة أن جميع الطيور حتى التي لا تطير تملك أجنحة. أما اليوم فهناك إثباتات أقوى تقوم على أساس علم الوراثة تؤكد وجود السلف المشترك. مثلا، كل خلية حية تستخدم نفس الحمض النووي كمادة وراثية (دنا ورنا)، وتستخدم نفس الحموض الأمينية كوحدات بناء للبروتينات. إضافة لذلك فإن جميع الأحياء تستخدم نفس الشفرة الوراثية (باختلافات ضئيلة ونادرة) لترجمة الحموض النووية إلى تسلسل الحموض الأمينية الذي تشكل البروتينات في النهاية. هذه التماثل العام لهذه السمات المشتركة في خلايا جميع الكائنات الحية يطرح بقوة فرضية السلف الواحد للأحياء.
المعلومات حول التطور المبكر للحياة يتضمن مدخلات من حقول معرفية كالاراضة (الجيولوجيا) وعلوم الكواكب planetary science. هذه العلوم توفر معلومات حول تاريخ الأرض المبكر والتغيرات التي أنتجت الحياة بالرغم من العديد من لامعلومات قد دمرتها الأحداث الجيولوجية عبر الزمن.
تاريخ الحياة
التطور الكيميائي (أو ما يدعى بالتخلق اللاحيوي) من تفاعلات كيميائية ذاتية التحفيز self-catalytic chemical reactions إلى ظهور الحياة (انظر أصل الحياة) لا يمثل جزءا من نظرية التطور الحيوية، من غير الواضح حتى الآن اللحظة التي تشكلت فيها الظروف المناسبة لهذه المجموعة المعقدة من التفاعلات إن حدثت لتشكل لنا الظاهرة الفريدة والغريبة والتي تدعى الحياة.

لم يعرف الكثير بعد عن التطورات المبكرة في الحياة. لكن من الواضح بشكل غير قابل للشك أن جميع الكائنات الحية تتشارك بسمات مشتركة بشكل واضح[وفقاً لِمَن؟]، بما في ذلك البنية الخلوية ونفس الشفرة الوراثية[بحاجة لمصدر]. معظم العلماء يفسرون هذا التشابه على أنه تشارك لجميع الأحياء بسلف وحيد مشترك، تطور عنه العمليات الخلوية الأساسية جميعها، مع هذا لا يوجد إجماع علمي على وجود علاقة بين الممالك الثلاث: الجراثيم القديمة والبكتريا، وحقيقيات النوى) كما ليس هناك إجماع حول أصل الحياة. محاولات إلقاء الضوء على التاريخ المبكر للحياة يركز بشكل عام على سلوك الجزيئات الضخمة macromolecule بخاصة الرنا وسلوك الأنظمة المعقدة.
نشوء التركيب الضوئي الأكسجيني (قبل حوالي 3 بلايين عام مضى) والنشوء التالي لغلاف جوي غير مرجع غني بالأكسجين يمكن تعقبه من خلال تشكل مناجم الحديد المجمع banded iron، ولاحقا الأسرة الحمراء من أكسيد الحديد. تعتبر عملية التركيب الضوئي شرطا أساسيا لتطور تنفس خلوي هوائي، الذي يعتقد أنه تشكل قبل بليونين من السنين.
الموقف العلمي المعارض
حمل لواء معارضة نظرية التطور في بداية ظهور النظرية العالم السويسري جان لويس رودلف أغاسي (Louis Agassiz)، حصل على دكتوراه من ميونيخ ثم أتم دراسة الطب 1830 وبعد ذلك أصبح أستاذ التاريخ الطبيعي في جامعة نيوشاتيل في سويسرا، ومن منجزاته دراسة شاملة عن أحافير الأسماك وطبعت في خمس مجلدات ما بين عامي 1833 و 1844 وحصل أغاسي على جائزة ولستون (Wollaston Medal) على هذا الإنجاز، وبعد ذلك انتقل أغاسي لدراسة المثلجات وكان من الرواد في هذا المجال. انتقل أغاسي إلى الولايات المتحدة وأمضى 27 عاماً معظمها كأستاذ في جامعة هارفارد.[37]
ظهرت معارضته لداروين بسبب تصريح الأول بأن أصل البشر واحد وهو ما رفضه أغاسي، حيث اعتبر في كتبه بأن الافارقه جنس دوني مختلف عن باقي البشر.[38] كما ذهب مع بعثة علمية إلى البرازيل في محاولة فاشلة لإيجاد دلائل تدحض نظرية التطور.[39]
الدين ونظرية التطور
ينفي بعض المتدينين صحة هذه النظرية حيث أنها تطرح رؤية بديلة لقصة الخلق على عكس ما جاء به الإسلام والمسيحية واليهودية؛ وهذا ما أثار جدلا عنيفا وبخاصة حول كتاب أصل الأنواع لداروين (كتب عام 1859) ووصل الخلاف حول نظرية التطور إلى القضاء (في القضية التي عرفت بمحاكمة القرد عام 1925) فيرى مؤيدو النظرية انه تحرش قضائي، ومنذ ذلك الحين استمر النزاع بين أنصار نظرية التطور ومعارضيها، الذين صاغوا بدورهم نظريات مثل نظرية الخلق (أو مانجده تحت تسمية معجزة الخلق في كتابات الباحثين المسلمين مثل عدنان أوكطار، انظر نظرية الخلق الإسلامية) ونظرية التصميم الذكي.
المراجع
- Futuyma, Douglas J. (2005)، Evolution، Sunderland, Massachusetts: Sinauer Associates, Inc، ISBN 0-87893-187-2.
- Futuyma, Douglas J. (2005)، Evolution، Sunderland، ماساتشوست: Sinauer Associates, Inc، ISBN 0-87893-187-2.
- Gould, Stephen J. (2002)، The Structure of Evolutionary Theory، Belknap Press، ISBN 0-674-00613-5، مؤرشف من الأصل في 19 مارس 2022.
- Lande, R. (1983)، "The measurement of selection on correlated characters"، Evolution، 37: 1210–1226.
- Haldane, J.B.S. (1953)، "The measurement of natural selection"، Proceedings of the 9th International Congress of Genetics، 1: 480–487.
- "Mechanisms: the processes of evolution"، Understanding Evolution، جامعة كاليفورنيا (بركلي)، مؤرشف من الأصل في 15 ديسمبر 2018، اطلع عليه بتاريخ 14 يوليو 2006.
- 95% من العلماء يؤمنون بنظرية التطور في أمريكا نسخة محفوظة 21 سبتمبر 2018 على موقع واي باك مشين.
- In the years after Darwin's publication and fame, numerous "predecessors" to natural selection were discovered, such as ويليام تشارلز ولز and باتريك ماثيو, who had published unelaborated and undeveloped versions of similar theories earlier to little or no attention. Historians acknowledge that Darwin was the first to develop the theory rigorously and developed it independently. On Matthew, one historian of evolution has written that he "did suggest a basic idea of selection, but he did nothing to develop it; and he published it in the appendix to a book on the raising of trees for shipbuilding. No one took him seriously, and he played no role in the emergence of Darwinism. Simple priority is not enough to earn a thinker a place in the history of science: one has to develop the idea and convince others of its value to make a real contribution. Darwin's notebooks confirm that he drew no inspiration from Matthew or any of the other alleged precursors."
- Bowler, Peter J. (2003)، Evolution: The History of an Idea، Berkeley: University of California Press، ص. 158، مؤرشف من الأصل في 14 أبريل 2022.
- Darwin's proposal of a توريث (أحياء) mechanism, شمولية التخلق, lacked evidence and was ultimately rejected, being replaced by علم الوراثة.Darwin’s Theory of Pangenesis نسخة محفوظة 13 يوليو 2006 على موقع واي باك مشين.
- Bowler, Peter J. (1989)، The Mendelian Revolution: The Emergence of Hereditarian Concepts in Modern Science and Society، Baltimore: Johns Hopkins University Press.
- The convinced Mendelians, such as وليام باتسون and Charles Benedict Davenport, and biometricians, such as Walter Frank Raphael Weldon and كارل بيرسون, became embroiled in a bitter debate, with Mendelians charging that the biometricians did not understand biology, and biometricians arguing that most biological traits exhibited continuous variation rather than the "jumps" expected by the early Mendelian theory. It is now recognized that the Mendelians were investigating Mendelian traits, those traits where existing variation is controlled by one gene and therefore is discrete, and the biometricians were investigating complex traits, where those traits were controlled by multiple genes, and the variation is therefore continuous.
- (Farber 2000, p. 136)
- (Watson 2005, p. 1257)
- Eldredge, Niles (Spring 2006)، "Confessions of a Darwinist"، The Virginia Quarterly Review: 32–53، مؤرشف من الأصل في 24 ديسمبر 2013، اطلع عليه بتاريخ 23 يناير 2008.
- Quammen, David (2004)، "Was Darwin Wrong?"، National Geographic Magazine، National Geographic، مؤرشف من الأصل في 24 فبراير 2018.
- Wyhe, John van (2002)، "Charles Darwin: gentleman naturalist"، The Complete Work of Charles Darwin Online، University of Cambridge، مؤرشف من الأصل في 27 سبتمبر 2018.
- Wyhe, John van (2002)، "Fertilisation of Orchids"، The Complete Works of Charles Darwin، University of Cambridge، مؤرشف من الأصل في 25 أكتوبر 2018، اطلع عليه بتاريخ 07 يناير 2008.
- Wyhe, John van (2002)، "Charles Darwin: gentleman naturalist"، The Complete Work of Charles Darwin Online، University of Cambridge، مؤرشف من الأصل في 27 سبتمبر 2018، اطلع عليه بتاريخ 16 يناير 2008.
- Delgado, Cynthia (2006)، "Finding the Evolution in Medicine"، NIH Record (National Institutes of Health)، مؤرشف من الأصل في 10 أبريل 2014، اطلع عليه بتاريخ 21 ديسمبر 2007.
- (Darwin 1872, p. 108.) Effects of the increased Use and Disuse of Parts, as controlled by Natural Selection "نسخة مؤرشفة"، مؤرشف من الأصل في 12 يونيو 2016، اطلع عليه بتاريخ 26 يناير 2018.
{{استشهاد ويب}}: صيانة CS1: BOT: original-url status unknown (link) - Ghiselin, Michael T. (September/October 1994)، "The Textbook Letter"، The Textbook League، مؤرشف من الأصل في 23 سبتمبر 2019، اطلع عليه بتاريخ 23 يناير 2008.
{{استشهاد بدورية محكمة}}: Cite journal requires|journal=(مساعدة)، تحقق من التاريخ في:|تاريخ النشر=(مساعدة)، الوسيط|الفصل=تم تجاهله (مساعدة) - "Understanding Evolution:Sex and genetic shuffling"، University of California, Berkely، مؤرشف من الأصل في 06 نوفمبر 2018.
- "Understanding Evolution:Gene flow"، University of California, Berkely، مؤرشف من الأصل في 07 نوفمبر 2018.
- (Gould (a) 1981, p. 24)
- Bejder L, Hall BK (2002)، "Limbs in whales and limblessness in other vertebrates: mechanisms of evolutionary and developmental transformation and loss"، Evol. Dev.، 4 (6): 445–458، doi:10.1046/j.1525-142X.2002.02033.x، PMID 12492145.
- Boughner JC, Buchtová M, Fu K, Diewert V, Hallgrímsson B, Richman JM (2007)، "Embryonic development of Python sebae - I: Staging criteria and macroscopic skeletal morphogenesis of the head and limbs"، Zoology (Jena)، 110 (3): 212–230، doi:10.1016/j.zool.2007.01.005، PMID 17499493.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Drummond, A; Strimmer, K (يوليو 2001)، "Evolution Library"، Bioinformatics (Oxford, England)، WGBH Educational Foundation، 17 (7): 662–3، ISSN 1367-4803، PMID 11448888، مؤرشف من الأصل في 08 يونيو 2019، اطلع عليه بتاريخ 23 يناير 2008.
{{استشهاد بدورية محكمة}}: الوسيط|الفصل=تم تجاهله (مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - "Evolution: a jouney into where we're from and where we're going"، WGBH Educational Foundation، 2001، مؤرشف من الأصل في 22 أكتوبر 2019، اطلع عليه بتاريخ 23 يناير 2008.
{{استشهاد بدورية محكمة}}: Cite journal requires|journal=(مساعدة)، الوسيط|الفصل=تم تجاهله (مساعدة). - Bambach, R.K.؛ Knoll, A.H.؛ Wang, S.C. (ديسمبر 2004)، "Origination, extinction, and mass depletions of marine diversity"، Paleobiology، 30 (4): 522–42، doi:10.1666/0094-8373(2004)030<0522:OEAMDO>2.0.CO;2، ISSN 0094-8373، مؤرشف من الأصل في 27 ديسمبر 2008، اطلع عليه بتاريخ 24 يناير 2008.
{{استشهاد بدورية محكمة}}: صيانة CS1: التاريخ والسنة (link) - Feduccia, Alan (1996)، The Origin and Evolution of Birds، New Haven: Yale University Press، ISBN 0-300-06460-8. and the recently-discovered تيكتاليك, which clarifies the development from سمك to رباعيات أطراف.
- Daeschler, Edward B., Shubin, Neil H., & Jenkins Jr, Farish A. (2006)، "A Devonian tetrapod-like fish and the evolution of the tetrapod body plan"، نيتشر (مجلة)، 440: 757–763، doi:10.1038/nature04639، مؤرشف من الأصل في 27 مايو 2017، اطلع عليه بتاريخ 14 يوليو 2006.
{{استشهاد بدورية محكمة}}: الوسيط غير المعروف|شهر=تم تجاهله (مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Chimpanzee Sequencing and Analysis Consortium (2005)، "Initial sequence of the chimpanzee genome and comparison with the human genome"، نيتشر (مجلة)، 437: 69–87.
- Britten, R.J. (2002)، "Divergence between samples of chimpanzee and human DNA sequences is 5%, counting indels"، Proc Natl Acad Sci U S A، 99: 13633–13635.
- Bejder L, Hall BK (2002)، "Limbs in whales and limblessness in other vertebrates: mechanisms of evolutionary and developmental transformation and loss"، Evol. Dev.، 4 (6): 445–58، doi:10.1046/j.1525-142X.2002.02033.x، PMID 12492145.
- Schweitzer M.H.؛ وآخرون (2005)، "Soft-tissue vessels and cellular preservation in Tyrannosaurus rex"، Science، 307 (5717): 1952–1955.
{{استشهاد بدورية محكمة}}: Explicit use of et al. in:|مؤلف=(مساعدة) - E.P. Evans: "The Authorship of the Glacial Theory", North American review Volume 145, Issue 368, July 1887. Accessed on February 25, 2008.
- Stephen Jay Gould, "Flaws in a Victorian Veil," Chapter 16 in The Panda's Thumb.
- Louis Agassiz نسخة محفوظة 26 يونيو 2017 على موقع واي باك مشين.
مصادر
- الحقيقـة الناصعة مجلة العلوم الأمريكية - النسخة العربية
- تطور أسماك إلى حيوانات رباعية الأرجل مجلة العلوم الأمريكية - النسخة العربية
- اختبار الانتخاب (الاصطفاء) الطبيعي مجلة العلوم الأمريكية - النسخة العربية
- التطور البيولوجي في الحياة اليومية مجلة العلوم الأمريكية - النسخة العربية
- أصل الأنواع: نشأة الأنواع الحية عن طريق الانتقاء الطبيعي أو الاحتفاظ بالأعراق المفضلة في أثناء الكفاح من أجل الحياة / تأليف تشارلس داروين؛ ترجمة مجدي محمود المليجي؛ تقديم سمير حنا صادق. القاهرة: المجلس الأعلى للثقافة، 2004.
- مقدّمة الماكروديناميكا الاجتماعية: النمذجة الرياضية لتطوّر المنظومة العالمية قبل سبعينيّات القرن الماضي «مجلة كلية الآداب لجامعة القاهرة» ٬ مجلد 68 ٬ سنة 2008 ٬ الجزء الثاني ٬ صفحات 148–181. (بالعربية).
- أندريه كاراطائف - مقدمة المكروديناميكا الاجتماعية. النمذجة الرياضية لتطور المنظومة العالمية. ISBN 5-484-00414-4(بالإنجليزية)
- نظرية التطور عند مفكري الإسلام، دراسة مقارنة / محفوظ علي عزام. القاهرة، دار الهداية، 1986.
انظر أيضًا

 بوابة تربية وتعليم
بوابة تربية وتعليم بوابة علم الأحياء التطوري
بوابة علم الأحياء التطوري بوابة علم الأحياء
بوابة علم الأحياء بوابة علوم
بوابة علوم