تولد الأوعية
تولد الأوعية[1] أو تكوّن الأوعية الدموية الجديدة[1] (بالإنجليزية: Angiogenesis) هي عملية فيزيولوجية تتشكل من خلالها أوعية دموية جديدة تتفرع عن أوعية موجودة مسبقًا.[2][3][4] بينما تحدث عملية تكوّن الأوعية أثناء تكوّن الجنين،[5] وبعدها تبدأ عملية تولد الأوعية لمعظم -إذا لم نقل للكل- وذلك بنمو الأوعية الدموية خلال النمو أو خلال المرض.[6]
| تولد الأوعية | |
|---|---|
 تولد الأوعية يلي تكوين الأوعية. تولد الأوعية يلي تكوين الأوعية. | |
| تسميات أخرى | Angiogenesis |
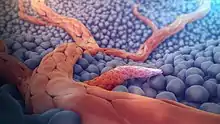
إن عملية تولد الأوعية هي عملية حيوية وحية في النمو والتطور وكذلك في عملية شفاء الجروح وخلال عملية تشكل النسيج الحبيبي. ومع ذلك فهي عملية أساسية لتحول وانتقال السرطان من حالة سرطان حميد إلى ورم خبيث، وذلك يُعالَج باستخدام مواد كابحة ومانعة لتولد الأوعية. اقتُرِحَ الدور الرئيس لتولد الاوعية في نمو السرطان لأول مرة في عام 1971 من قبل العالم جودا فولكمان، والذي وصف السرطان بأنه ساخن ودموي.[7]
الأنواع
تولد الأوعية بالتفريخ
كانت عملية تفريخ الأوعية أول شكل حُدِّد لتولد الأوعية الدموية وبسبب ذلك أصبح مفهوماً أكثر بكثير من تولد الأوعية الدموية الانغلافي. يحدث في عدة مراحل جيدة التوصيف. تأتي الإشارة الأولية من مناطق الأنسجة الخالية من الأوعية الدموية. يتسبب نقص الأكسجة الملحوظ في هذه المناطق في أن تطلب الأنسجة وجود المغذيات والأكسجين الذي يسمح للأنسجة بالقيام بأنشطة التمثيل الغذائي. وبسبب هذا فإن الخلايا المتنيّة تفرز عامل نمو بطانة الأوعية الدموية (VEGF-A) وهو عامل نمو محفز لتولد الأوعية.[8] تنشط هذه الإشارات البيولوجية المستقبلات على الخلايا البطانية الموجودة في الأوعية الدموية الموجودة مسبقًا. ثانيًا: تبدأ الخلايا البطانية المنشطة -والمعروفة أيضًا باسم الخلايا الطرفية- في إطلاق إنزيمات تسمى الببتيداز التي تعمل على تحلل الغشاء القاعدي للسماح للخلايا البطانية بالهروب من جدران الوعاء الدموي الأصلية. ثم تتكاثر الخلايا البطانية في المطرس المحيط وتشكل أشطاء صلبة تربط الأوعية المجاورة. توجد الخلايا التي تتكاثر خلف الخلايا الطرفية وتُعرف باسم خلايا الساق. يسمح تكاثر هذه الخلايا لشطء الشعيرات بالنمو في الطول في وقت واحد.
عندما تمتد الأشطاء نحو مصدر التحفيز الوعائي تهاجر الخلايا البطانية جنبًا إلى جنب باستخدام جزيئات الالتصاق التي تسمى الإنتغرينات. ثم تشكل هذه الأشطاء حلقات لتصبح تجويفًا كاملًا للأوعية بينما تهاجر الخلايا إلى موقع تكوين الأوعية. يحدث الشطء بمعدل عدة ملليمترات في اليوم، ويمكن الأوعية الجديدة من النمو عبر الفجوات في الأوعية الدموية. إنه يختلف بشكل ملحوظ عن تولد الأوعية بالانشطار لأنه يشكل أوعية جديدة تمامًا بدلاً من انشطار الأوعية الموجودة.
تولد الأوعية بالانغلاف
تولد الأوعية بالانغلاف -المعروف أيضًا باسم تولد الأوعية بالانشطار- هو تكوين وعاء دموي جديد عن طريق انشطار وعاء دموي موجود إلى قسمين.
لوحظ الانغلاف لأول مرة في الفئران حديثي الولادة. في هذا النوع من تولد الأوعية الدموية يمتد جدار الشعيرات الدموية إلى التجويف لتقسيم وعاء واحد إلى قسمين. هناك أربع مراحل لتولد الأوعية بالانغلاف. أولاً: ينشئ الجداران الشعريان المتعارضان منطقة تماس. ثانيًا: يُعاد تنظيم تقاطعات الخلايا البطانية وتثقب الطبقة الثنائية الوعاء للسماح لعوامل النمو والخلايا بالاختراق في التجويف. ثالثًا: تُكوَّن نواة بين السفينتين الجديدتين في منطقة التلامس المملوءة بالخلايا الحولية والأرومات الليفية العضلية. تبدأ هذه الخلايا في وضع ألياف الكولاجين في اللب لتوفير نسيج بيني خارج الخلية لنمو تجويف الوعاء الدموي. أخيرًا يُجسَّد اللب دون أي تعديلات على الهيكل الأساسي. يعتبر الانغلاف مهمًا لأنه إعادة تنظيم للخلايا الموجودة. يسمح بزيادة كبيرة في عدد الشعيرات الدموية دون زيادة مقابلة في عدد الخلايا البطانية. هذا مهم بشكل خاص في التطور الجنيني حيث لا توجد موارد كافية لإنشاء الأوعية الدموية الدقيقة الغنية بالخلايا الجديدة في كل مرة يتطور فيها وعاء جديد.[9]
علم وظائف الأعضاء
تنبيه ميكانيكي
لا يتميز التنبيه الميكانيكي لتكوين الأوعية الدموية بشكل جيد. فهناك الكثير من الجدل فيما يتعلق بإجهاد القص الذي يعمل على الشعيرات الدموية للتسبب في تولُّد الأوعية على الرغم من أن المعرفة الحالية تشير إلى أن زيادة تقلصات العضلات قد تزيد من توليد الأوعية الدموية.[10] قد يكون هذا بسبب زيادة إنتاج أحادي أكسيد النيتروجين أثناء التمرين. ينتج عن أكسيد النيتريك توسع الأوعية الدموية.
تنبيه كيميائي
يحدث التنبيه الكيميائي لتولد الأوعية بواسطة عدد من البروتينات المولدة للأوعية (على سبيل المثال: الإنتغرين والبروستاغلاندين)، كما تشمل هذه البروتينات العديد من عوامل النمو مثل: VEGF وFGF.
فكرة عامة
| المحفز | آلية العمل |
|---|---|
| عوامل نمو الأرومة الليفية | يعزز تكاثر وتمايز الخلايا البطانية وخلايا العضلات الملساء والأرومات الليفية |
| عامل النمو البطاني الوعائي | يؤثر على النفاذية. |
| مستقبل VEGFR ونيوروبيلين-1 | تكامل إشارات الحياة. |
| أنجيوبويتين1 وأنجيوبويتين2 | استقرار الأوعية. |
| عامل نمو مشتق من الصفيحات ومستقبل عامل نمو مشتق من الصفيحات | يجلب خلايا العضلات الملساء. |
| عامل النمو المحول بيتا وإندوجلين ومستقبل عامل نمو المحول بيتا | ↑ إنتاج النسيج البيني خارج الخلية |
| هستامين | |
| كادرين 5 وCD31 | جزيئات الموصلات الخلوية للخلايا البطانية |
| إفرين | يحدد تكوين الشرايين والأوردة. |
| منشط البلازمينوجين | يعيد تشكيل النسيج البيني خارج الخلية، ويطلق وينشط عوامل النمو |
| مثبط منشط البلازمينوجين-1 | استقرار الأوعية الدموية القريبة. |
| مخلقة أكسيد النتريك |
عوامل نمو الأرومة الليفية
تتكون عائلة عوامل نمو الخلايا الليفية (FGF) مع أعضائها النموذجيين عامل نمو الأرومة الليفية -1 (عامل نمو الأرومة الليفية الحمضي) وعامل نمو الأرومة الليفية-2 (عامل نمو الأرومة الليفية القاعدي) حتى الآن من 22 عضوًا معروفًا على الأقل.[11] معظمها عبارة عن ببتيدات أحادية السلسلة تتراوح من 16 إلى 18 كيلو دالتون وتظهر تقاربًا كبيرًا مع الهيبارين وكبريتات الهيبارين. بشكل عام تحفز عوامل نمو الخلايا الليفية مجموعة متنوعة من الوظائف الخلوية من خلال الارتباط بمستقبلات عوامل نمو الخلايا الليفية على سطح الخلية في وجود بروتينات الهيبارين. تتكون عائلة مستقبلات عوامل نمو الخلايا الليفية من سبعة أعضاء وجميع بروتينات المستقبلات عبارة عن مستقبلات التيروزين كينازات أحادية السلسلة التي تُنشَّط من خلال الفسفرة الذاتية التي تحدثها آلية تضاعف مستقبلات عوامل نمو الخلايا الليفية بوساطة. يؤدي تنشيط المستقبلات إلى ظهور تسلسل نقل الإشارة الذي يؤدي إلى تنشيط الجينات والاستجابات البيولوجية المتنوعة، بما في ذلك تمايز الخلايا والتكاثر وحل المطرس وبالتالي بدء عملية النشاط الانقسامي الضروري لنمو الخلايا البطانية والأرومات الليفية وخلايا العضلات الملساء. يمكن لعوامل نمو الخلايا الليفية الفريدة من بين جميع أعضاء عائلة عوامل نمو الخلايا الليفية البالغ عددهم 22 الارتباط بجميع الأنواع الفرعية لمستقبلات عوامل نمو الخلايا الليفية السبعة مما يجعله العضو الأوسع مفعولًا في عائلة عوامل نمو الخلايا الليفية وميتوجين قوي لأنواع الخلايا المتنوعة اللازمة لتكوين الأوعية الدموية استجابة في الأنسجة التالفة (ناقصة التأكسج)، حيث يحدث انتظام في مستقبلات عوامل نمو الخلايا الليفية.[12] يحفز عامل نمو الخلايا الليفية-1 تكاثر وتمايز جميع أنواع الخلايا اللازمة لبناء الأوعية الدموية الشريانية بما في ذلك الخلايا البطانية وخلايا العضلات الملساء، هذه الحقيقة تميز عامل نمو الخلايا الليفية-1 عن عوامل النمو الأخرى المؤيدة لتولد الأوعية مثل عامل النمو البطاني الوعائي (VEGF) الذي يقود بشكل أساسي تكوين شعيرات دموية جديدة.[13][14]
إلى جانب عامل نمو الخلايا الليفية-1 تتمثل إحدى أهم وظائف عامل نمو الخلايا الليفية -2 تعزيز تكاثر الخلايا البطانية والتنظيم الفيزيائي للخلايا البطانية في هياكل تشبه الأنبوب، وبالتالي تعزيز تكوين الأوعية. عامل نمو الخلايا الليفية-2 هو عامل تولد الأوعية أكثر فاعلية من عامل النمو البطاني الوعائي أو عامل النمو المشتق من الصفائح الدموية، ومع ذلك فهو أقل فعالية من عامل نمو الخلايا الليفية-1. بالإضافة إلى تحفيز نمو الأوعية الدموية، فإن عامل نمو الأرومة الليفية -1 وعامل نمو الأرومة الليفية -2 يلعبان دورًا مهمًا في التئام الجروح. فهي تحفز تكاثر الخلايا الليفية والخلايا البطانية التي تؤدي إلى تكوين الأوعية الدموية وتطوير الأنسجة الحبيبية. كلاهما يزيد من إمداد الدم ويملأ مساحة / تجويف الجرح في وقت مبكر من عملية التئام الجروح.
عوامل النمو البطانية الوعائية
ثبت أن عامل النمو البطاني الوعائي (VEGF) مساهم رئيسي في تكوين الأوعية الدموية مما يزيد من عدد الشعيرات الدموية في شبكة معينة. أظهرت الدراسات الأولية في المختبر أن الخلايا البطانية الشعرية في الأبقار تتكاثر وتظهر علامات على هياكل الأنبوب عند التحفيز بواسطة عامل النمو البطاني الوعائي وعامل نمو الخلايا الليفية -2 على الرغم من أن النتائج كانت أكثر وضوحًا مع عامل النمو البطاني الوعائي.[15] يعد تنظيم عامل النمو البطاني الوعائي مكونًا رئيسيًا للاستجابة الفسيولوجية للتمرين ويشتبه في أن دوره في تكوين الأوعية هو علاج محتمل لإصابات الأوعية الدموية.[16][17][18][19] تظهر الدراسات في المختبر بوضوح أن عامل النمو البطاني الوعائي هو محفز قوي لتكوين الأوعية لأنه في وجود عامل النمو هذا سوف تتكاثر الخلايا البطانية المطلية وتهاجر وتشكل في النهاية هياكل أنبوبية تشبه الشعيرات الدموية.[10] يتسبب عامل النمو البطاني الوعائي في سلسلة إشارات ضخمة في الخلايا البطانية. يؤدي الارتباط بمستقبل عامل النمو البطاني الوعائي-2 إلى بدء سلسلة إشارات التيروزين كيناز التي تحفز إنتاج العوامل التي تحفز بشكل مختلف نفاذية الأوعية (إنتاج أكسيد النيروجين)، التكاثر/البقاء (عامل نمو الخلايا الليفية -2)، الهجرة (ICAMs / VCAMs / ميتالوبروتياز المادة الخلالية) وأخيرًا التمايز إلى أوعية دموية ناضجة. ميكانيكيًا يُنظَّم عامل النمو البطاني الوعائي مع تقلصات العضلات نتيجة لزيادة تدفق الدم إلى المناطق المصابة. يؤدي التدفق المتزايد أيضًا إلى زيادة كبيرة في إنتاج mRNA لمستقبلات عامل النمو البطاني الوعائي-1 و 2. الزيادة في إنتاج المستقبلات تعني أن تقلصات العضلات يمكن أن تؤدي إلى زيادة تنظيم سلسلة الإشارات المتعلقة بتكوين الأوعية. كجزء من سلسلة الإشارات المولدة للأوعية، يعتبر NO على نطاق واسع مساهماً رئيسياً في الاستجابة المولدة للأوعية لأن تثبيط أكسيد النيتروجين يقلل بشكل كبير من تأثيرات عوامل النمو الوعائية. ومع ذلك فإن تثبيط أكسيد النيتروجين أثناء التمرين لا يمنع تكوين الأوعية مما يشير إلى وجود عوامل أخرى تشارك في الاستجابة لتولد الأوعية.[10]
ميتالوبروتياز المادة الخلالية
يُعد ميتالوبروتياز المادة الخلالية مساهم رئيسي آخر في تولد الأوعية. يساعد ميتالوبروتياز المادة الخلالية في تحلل البروتينات التي تحافظ على صلابة جدران الأوعية. يسمح هذا التحلل البروتيني للخلايا البطانية بالهروب إلى المصفوفة الخلالية كما يظهر في تكوين الأوعية الدموية.[20] يمنع تثبيط ميتالوبروتياز المادة الخلالية تكوين شعيرات دموية جديدة. يتم تنظيم هذه الإنزيمات بشكل كبير أثناء عملية تكوين الأوعية لأن تدمير النسيج البيني خارج الخلية من شأنه أن يقلل من سلامة الأوعية الدموية الدقيقة.[10]
DLL4
DLL4 هو بروتين له تأثير تنظيمي سلبي على تكوين الأوعية.[21][22] Dll4 عبارة عن لجين عبر الغشاء لعائلة المستقبلات من الدرجة الأولى. كانت هناك العديد من الدراسات التي أجريت والتي عملت على تحديد عواقب مثل DLL4. إحدى الدراسات على وجه الخصوص قيمت آثار DII4 على الأوعية الدموية الورمية ونموها.[23] لكي ينمو الورم ويتطور يجب أن يكون لديه الأوعية الدموية المناسبة. يعد مسار عوامل النمو البطانية الوعائية أمرًا حيويًا لتطوير الأوعية الدموية التي بدورها تساعد الأورام على النمو. يؤدي الحصار المشترك لعوامل النمو البطانية الوعائية و DII4 إلى تثبيط تطور الورم وتكوين الأوعية في جميع أنحاء الورم. ويرجع ذلك إلى إعاقة إرسال الإشارات في إشارات الخلايا البطانية التي تمنع تكاثر وتكاثر هذه الخلايا البطانية. مع هذا التثبيط لا تنمو الخلايا بشكل لا يمكن السيطرة عليه لذلك يتوقف السرطان عند هذه النقطة. ومع ذلك فإنه إذا أُزيل التثبيط ستبدأ الخلايا في التكاثر مرة أخرى.[24]
سمافورين فئة 3
تنظم السمافورينات من الفئة 3 (SEMA3s) تولد الأوعية عن طريق تعديل التصاق الخلايا البطانية والهجرة والتكاثر والبقاء وتوظيف الحبيبات.[25] علاوة على ذلك يمكن أن يتداخل السيمافورين مع تكوين الأوعية الدموية بوساطة عوامل النمو البطانية الوعائية نظرًا لأن كل من SEMA3 وعوامل النمو البطانية الوعائية-أ يتنافسان على ارتباط مستقبلات نيوروبيلين في الخلايا البطانية.[26][27] لذلك قد تكون مستويات التعبير النسبي لـ SEMA3s وعوامل النمو البطانية الوعائية-أ مهمة لتولد الأوعية.[25]
التثبيط الكيميائي
يمكن أن يكون مثبط تكوّن الأوعية داخليًا أو يأتي من الخارج كعقار أو مكون غذائي.
التطبيق في الطب
تولد الأوعية كهدف علاجي
قد يكون تولد الأوعية هدفًا لمكافحة الأمراض مثل أمراض القلب التي تتميز إما بضعف الأوعية الدموية أو الأوعية الدموية غير الطبيعية.[28] قد يساعد استخدام مركبات معينة قد تمنع أو تحفز تخليق أوعية دموية جديدة في الجسم في مكافحة مثل هذه الأمراض. قد يؤثر وجود الأوعية الدموية التي لا ينبغي أن تكون هناك على الخواص الميكانيكية للأنسجة مما يزيد من احتمالية الفشل. قد يؤدي عدم وجود أوعية دموية في نسيج صالح أو نشط التمثيل الغذائي إلى إعاقة الإصلاح أو الوظائف الأساسية الأخرى. تنجم العديد من الأمراض -مثل الجروح المزمنة الإقفارية- عن فشل أو نقص تكوين الأوعية الدموية، ويمكن علاجها عن طريق التوسيع الموضعي للأوعية الدموية، وبالتالي جلب مغذيات جديدة إلى المكان المصاب مما يسهل الإصلاح. قد تنشأ أمراض أخرى -مثل التنكس البقعي المرتبط بالعمر- عن طريق توسع محلي في الأوعية الدموية مما يتداخل مع العمليات الفسيولوجية الطبيعية.
يُقسَّم التطبيق السريري الحديث لمبدأ تولد الأوعية إلى مجالين رئيسيين: العلاجات المضادة لتولد الأوعية (والتي بدأ بها البحث في مجال تكوين الأوعية) والعلاجات المؤيدة لتولد الأوعية. في حين تُستخدم العلاجات المضادة لتولد الأوعية لمحاربة السرطان والأورام الخبيثة[29][30] والتي تتطلب وفرة من الأكسجين والمواد المغذية للتكاثر، تُستكشف العلاجات المؤيدة لتولد الأوعية كخيارات لعلاج أمراض القلب والأوعية الدموية -السبب الأول للوفاة في العالم الغربي-. كان أحد التطبيقات الأولى للطرق المؤيدة لتولد الأوعية في البشر تجربة ألمانية تستخدم عامل نمو الخلايا الليفية 1 (FGF-1) لعلاج مرض الشريان التاجي.[13][31][32]
فيما يتعلق بآلية العمل، يمكن تقسيم الطرق المؤيدة لتولد الأوعية إلى ثلاث فئات رئيسية: العلاج الجيني -أي استهداف الجينات ذات الأهمية للتضخيم أو التثبيط- والعلاج ببتبديل البروتين -والذي يعالج في المقام الأول عوامل النمو الوعائية مثل FGF-1 أو عامل النمو البطاني الوعائي VEGF- والعلاجات القائمة على الخلايا -والتي تنطوي على زرع أنواع معينة من الخلايا.
لا تزال هناك مشكلات خطيرة لم تُحَل تتعلق بالعلاج الجيني. تشمل الصعوبات التكامل الفعال للجينات العلاجية في جينوم الخلايا المستهدفة وتقليل مخاطر الاستجابة المناعية غير المرغوب فيها والسمية المحتملة والاستمناع والاستجابات الالتهابية والتسرطن المتعلق بالنواقل الفيروسية المستخدمة في زرع الجينات والتعقيد الهائل للجينات أساس تولد الأوعية. من المرجح أن تكون الاضطرابات الأكثر شيوعًا عند البشر -مثل أمراض القلب وارتفاع ضغط الدم والسكري ومرض آلزهايمر- ناجمة عن التأثيرات المجتمعة للاختلافات في العديد من الجينات، وبالتالي فإن حقن جين واحد قد لا يكون مفيدًا بشكل كبير في مثل هذه الأمراض.
على النقيض من ذلك فإن العلاج بالبروتين المؤيد لتولد الأوعية يستخدم بروتينات محددة جيدًا ومنظمة بدقة مع جرعات مثلى محددة مسبقًا من البروتين الفردي للحالات المرضية مع تأثيرات بيولوجية معروفة.[2] من ناحية أخرى فإن عقبة العلاج بالبروتين هي طريقة التوصيل. لا تكون طرق إعطاء البروتين عن طريق الفم أو الوريد أو داخل الشرايين أو العضل فعالة دائمًا؛ حيث يمكن استقلاب البروتين العلاجي أو تصفيته قبل أن يتمكن من دخول النسيج المستهدف. لا تزال العلاجات المؤيدة لتولد الأوعية القائمة على الخلايا في المراحل الأولى من البحث مع وجود العديد من الأسئلة المفتوحة بشأن أفضل أنواع الخلايا والجرعات التي يجب استخدامها.
تولد الأوعية بالورم
الخلايا السرطانية هي الخلايا التي فقدت قدرتها على الانقسام بطريقة صحيحة. يتكون الورم الخبيث من مجموعة من الخلايا السرطانية سريعة الانقسام والنمو والتي تتراكم عليها الطفرات تدريجياً. ومع ذلك تحتاج الأورام إلى إمداد دم مخصص لتوفير الأكسجين والعناصر الغذائية الأساسية الأخرى التي تحتاجها من أجل النمو إلى ما بعد حجم معين (بشكل عام 1-2 مم).[33][34]
تحفز الأورام تولد الأوعية الدموية عن طريق إفراز عوامل النمو المختلفة (مثل عوامل النمو البطانية الوعائية) والبروتينات. يمكن لعوامل النمو مثل عوامل نمو الأرومة الليفية وعوامل النمو البطانية الوعائية أن تحفز نمو الشعيرات الدموية في الورم والذي يشك بعض الباحثين في توفير العناصر الغذائية المطلوبة مما يسمح بتوسع الورم. على عكس الأوعية الدموية الطبيعية تتوسع الأوعية الدموية للورم بشكل غير منتظم.[35] يعتقد أطباء آخرون أن تولد الأوعية يعمل حقًا كمسار للنفايات حيث يزيل المنتجات البيولوجية النهائية التي تفرز عن طريق الانقسام السريع للخلايا السرطانية. في كلتا الحالتين يعتبر تكوين الأوعية خطوة ضرورية ومطلوبة للانتقال من مجموعة صغيرة من الخلايا غير المؤذية -والتي يُقال غالبًا أنها بحجم الكرة المعدنية في نهاية قلم حبر جاف- إلى ورم كبير. تكون الأوعية الدموية ضرورية أيضًا لانتشار الورم أو ورم خبيث. يمكن للخلايا السرطانية المنفردة أن تنفصل عن الورم الصلب الثابت وتدخل الأوعية الدموية وتُنقَل إلى موقع بعيد حيث يمكنها الزرع والبدء في نمو ورم ثانوي. تشير الدلائل الآن إلى أن الأوعية الدموية في ورم صلب معين قد تكون في الواقع أوعية فسيفساء تتكون من خلايا بطانية وخلايا ورمية. تسمح هذه الفسيفساء بإلقاء كبير من الخلايا السرطانية في الأوعية الدموية مما قد يساهم في ظهور الخلايا السرطانية المنتشرة في الدم المحيطي للمرضى المصابين بالأورام الخبيثة.[36] سيتطلب النمو اللاحق لهذه الانبثاثات أيضًا إمدادًا بالمغذيات والأكسجين ومسارًا للتخلص من النفايات.
لطالما اعتبرت الخلايا البطانية أكثر استقرارًا وراثيًا من الخلايا السرطانية. يمنح هذا الاستقرار الجيني ميزة لاستهداف الخلايا البطانية باستخدام العلاج المضاد لتكوُّن الأوعية مقارنةً بالعلاج الكيميائي الموجه إلى الخلايا السرطانية والتي تتحول بسرعة وتكتسب مقاومة الأدوية للعلاج. لهذا السبب يُعتقد أن الخلايا البطانية هدف مثالي للعلاجات الموجهة ضدها.[37]
تكون الأوعية الدموية بالورم
تبدأ آلية تكوين الأوعية الدموية عن طريق تولد الأوعية عن طريق الانقسام التلقائي للخلايا السرطانية بسبب طفرة. ثم تنطلق منبهات الأوعية الدموية بواسطة الخلايا السرطانية. ثم تنتقل بعد ذلك إلى الأوعية الدموية الموجودة بالفعل والقريبة وتنشط مستقبلات الخلايا البطانية. يؤدي هذا إلى إطلاق الإنزيمات المحللة للبروتين من الأوعية الدموية. تستهدف هذه الإنزيمات نقطة معينة في الأوعية الدموية وتتسبب في تكوين مسام. هذه هي النقطة التي سينمو منها الأوعية الدموية الجديدة. السبب في حاجة الخلايا السرطانية إلى إمداد الدم هو أنها لا تستطيع النمو أكثر من 2-3 ملليمترات في القطر دون وجود إمداد دم ثابت يعادل حوالي 50-100 خلية.[38]
تولد الأوعية بالمرض القلبي الوعائي
يمثل تولد الأوعية هدفًا علاجيًا ممتازًا لعلاج أمراض القلب والأوعية الدموية. فهي عملية فسيولوجية قوية تكمن وراء الطريقة الطبيعية التي تستجيب بها أجسامنا لتناقص إمداد الدم للأعضاء الحيوية، أي تكوين الأوعية الجديدة (إنتاج أوعية جانبية جديدة للتغلب على الأذية الإقفارية).[13] أُجرِيَ عدد كبير من الدراسات قبل السريرية باستخدام العلاجات القائمة على البروتين والجينات والخلايا في نماذج حيوانية لنقص تروية القلب بالإضافة إلى نماذج لأمراض الشرايين الطرفية. أدت النجاحات الموثوقة والقابلة للتكرار في هذه الدراسات التي أجريت على الحيوانات في وقت مبكر إلى حماس كبير بأن هذا النهج العلاجي الجديد يمكن ترجمته بسرعة إلى فائدة سريرية لملايين المرضى في العالم الغربي الذين يعانون من هذه الاضطرابات. ومع ذلك فإن عقدًا من الاختبارات السريرية لكل من العلاجات القائمة على الجينات والبروتينات المصممة لتحفيز تكوين الأوعية في الأنسجة والأعضاء ناقصة الاستخدام أدى من خيبة أمل إلى أخرى. على الرغم من أن كل هذه القراءات قبل السريرية -والتي قدمت وعودًا كبيرة لانتقال علاج تكوين الأوعية من الحيوانات إلى البشر- دُمِجَت بطريقة أو بأخرى في التجارب السريرية المبكرة إلا أن إدارة الغذاء والدواء الأمريكية، حتى عام 2007 أصرت على أن العلاج الأولي يجب أن تكون نقطة النهاية للموافقة على العامل المولد للأوعية تحسنًا في أداء التمارين للمرضى المعالجين.[39]
تشير هذه الإخفاقات إلى أن هذه هي الأهداف الجزيئية الخاطئة للحث على تكوين الأوعية الدموية وأنه لا يمكن استخدامها بشكل فعال إلا إذا صيغَت وأُديرَت بشكل صحيح أو أن عرضها في سياق البيئة المكروية الخلوية الشاملة قد يلعب دورًا حيويًا في فائدتها. قد يكون من الضروري تقديم هذه البروتينات بطريقة تحاكي أحداث الإشارات الطبيعية بما في ذلك التركيز والملامح المكانية والزمانية وعرضها المتزامن أو المتسلسل مع عوامل أخرى مناسبة.[40]
التمارين
يرتبط تولد الأوعية عمومًا بالتمارين الهوائية وتمارين التحمل. بينما ينتج عن تكوّن الشرايين تغييرات في الشبكة تسمح بزيادة كبيرة في كمية التدفق الكلي في الشبكة، يتسبب تولد الأوعية في حدوث تغييرات تسمح بإيصال قدر أكبر من المغذيات على مدى فترة زمنية طويلة. صُمِّمَت الشعيرات الدموية لتوفير أقصى قدر من كفاءة توصيل المغذيات، لذا فإن الزيادة في عدد الشعيرات الدموية تسمح للشبكة بتقديم المزيد من العناصر الغذائية في نفس الوقت. يسمح عدد أكبر من الشعيرات الدموية أيضًا بتبادل أكبر للأكسجين في الشبكة. هذا مهم للغاية لتدريب التحمل لأنه يسمح للشخص بمواصلة التدريب لفترة طويلة من الزمن. ومع ذلك فإنه لا يوجد دليل تجريبي يشير إلى أن زيادة الشعيرات الدموية مطلوبة في تمارين التحمل لزيادة الحد الأقصى لتوصيل الأكسجين.[10]
التنكس البقعي
يؤدي الإفراط في التعبير عن عامل النمو البطاني الوعائي إلى زيادة نفاذية الأوعية الدموية بالإضافة إلى تحفيز تكوين الأوعية. في حالة التنكس البقعي الرطب يتسبب عامل النمو البطاني الوعائي في تكاثر الشعيرات الدموية في شبكية العين. نظرًا لأن الزيادة في تولد الأوعية تؤدي أيضًا إلى الوذمة فإن الدم وسوائل الشبكية الأخرى تتسرب إلى شبكية العين مما يتسبب في فقدان الرؤية. تُستخدم الآن الأدوية المضادة لتولد الأوعية التي تستهدف مسارات عامل النمو البطاني الوعائي بنجاح لعلاج هذا النوع من الضمور البقعي.
أجزاء الأنسجة المعدلة الوراثية
من الضروري تولد الأوعية الدموية من الجسم المضيف إلى نسيج معدلة وراثيًا. غالبًا ما يعتمد التكامل الناجح على تكوين الأوعية الدموية الشامل للبنية لأنه يوفر الأكسجين والمغذيات ويمنع النخر في المناطق المركزية للزرع.[41] وقد ثبت أن عامل النمو المشتق من الصفيحات يعمل على استقرار الأوعية الدموية في سقالات الكولاجين والغليكوز أمينوغليكان.[42]
التقدير الكمي
قياس معلمات الأوعية الدموية مثل كثافة الأوعية الدموية الدقيقة له مضاعفات مختلفة بسبب التلوين التفضيلي أو التمثيل المحدود للأنسجة حسب الأقسام النسيجية. أظهرت الأبحاث الحديثة إعادة بناء كاملة ثلاثية الأبعاد لبنية الأوعية الدموية للورم وتقدير هياكل الأوعية الدموية في الأورام الكاملة في نماذج حيوانية.[43]
انظر أيضًا
المراجع
- "Al-Qamoos القاموس - English Arabic dictionary / قاموس إنجليزي عربي"، www.alqamoos.org، مؤرشف من الأصل في 21 ديسمبر 2019، اطلع عليه بتاريخ 21 ديسمبر 2019.
- Angiogenesis insights from a systematic overview، New York: Nova Science، 2013، ISBN 978-1-62618-114-4.
- "Pericytes at the intersection between tissue regeneration and pathology"، Clinical Science، 128 (2): 81–93، يناير 2015، doi:10.1042/CS20140278، PMC 4200531، PMID 25236972.
- "Type-2 pericytes participate in normal and tumoral angiogenesis"، American Journal of Physiology. Cell Physiology، 307 (1): C25-38، يوليو 2014، doi:10.1152/ajpcell.00084.2014، PMC 4080181، PMID 24788248.
- "Vasculogenesis"، Annual Review of Cell and Developmental Biology، 11: 73–91، 1995، doi:10.1146/annurev.cb.11.110195.000445، PMID 8689573.
- "Molecular mechanisms of vasculogenesis and embryonic angiogenesis"، Journal of Cellular Physiology، 173 (2): 206–10، نوفمبر 1997، doi:10.1002/(SICI)1097-4652(199711)173:2<206::AID-JCP22>3.0.CO;2-C، PMID 9365523.
- John S. Penn (11 مارس 2008)، Retinal and Choroidal Angiogenesis، Springer، ص. 119–، ISBN 978-1-4020-6779-2، مؤرشف من الأصل في 10 أكتوبر 2021، اطلع عليه بتاريخ 26 يونيو 2010.
- Adair TH, Montani JP. Angiogenesis. San Rafael (CA): Morgan & Claypool Life Sciences; 2010. Chapter 1, Overview of Angiogenesis. Available from: https://www.ncbi.nlm.nih.gov/books/NBK53238/ نسخة محفوظة 2 فبراير 2021 على موقع واي باك مشين.
- "Intussusceptive angiogenesis: its emergence, its characteristics, and its significance"، Developmental Dynamics، 231 (3): 474–88، نوفمبر 2004، doi:10.1002/dvdy.20184، PMID 15376313، S2CID 35018922.
- "What makes vessels grow with exercise training?"، Journal of Applied Physiology، 97 (3): 1119–28، سبتمبر 2004، doi:10.1152/japplphysiol.00035.2004، PMID 15333630.
- "Fibroblast growth factors"، Genome Biology، 2 (3): REVIEWS3005، 2001، doi:10.1186/gb-2001-2-3-reviews3005، PMC 138918، PMID 11276432.
- "X-ray crystal structure of human acidic fibroblast growth factor"، Biochemistry، 35 (7): 2086–94، فبراير 1996، CiteSeerX 10.1.1.660.7607، doi:10.1021/bi9521755، PMID 8652550.
- "FGF-1: a human growth factor in the induction of neoangiogenesis"، Expert Opinion on Investigational Drugs، 7 (12): 2011–5، ديسمبر 1998، doi:10.1517/13543784.7.12.2011، PMID 15991943.
- "Insights from angiogenesis trials using fibroblast growth factor for advanced arteriosclerotic disease"، Trends in Cardiovascular Medicine، 13 (3): 116–22، أبريل 2003، doi:10.1016/S1050-1738(02)00259-1، PMID 12691676.
- "Synergistic effects of vascular endothelial growth factor and basic fibroblast growth factor on the proliferation and cord formation of bovine capillary endothelial cells within collagen gels"، Laboratory Investigation; A Journal of Technical Methods and Pathology، 69 (5): 508–17، نوفمبر 1993، PMID 8246443.
- "Exercise-induced overexpression of angiogenic factors and reduction of ischemia/reperfusion injury in stroke"، Current Neurovascular Research، 1 (5): 411–20، ديسمبر 2004، doi:10.2174/1567202043361875، PMID 16181089، مؤرشف من الأصل في 19 أبريل 2012.
- "Angiogenic growth factor response to acute systemic exercise in human skeletal muscle"، Journal of Applied Physiology، 96 (1): 19–24، يناير 2004، doi:10.1152/japplphysiol.00748.2003، PMID 12949011، S2CID 12750224.
- "Circulating plasma VEGF response to exercise in sedentary and endurance-trained men"، Journal of Applied Physiology، 96 (4): 1445–50، أبريل 2004، doi:10.1152/japplphysiol.01031.2003، PMID 14660505.
- "Angiogenic growth factor expression in rat skeletal muscle in response to exercise training"، American Journal of Physiology. Heart and Circulatory Physiology، 284 (5): H1668-78، مايو 2003، doi:10.1152/ajpheart.00743.2002، PMID 12543634.
- "Matrix metalloproteinase activity is required for activity-induced angiogenesis in rat skeletal muscle"، American Journal of Physiology. Heart and Circulatory Physiology، 279 (4): H1540-7، أكتوبر 2000، doi:10.1152/ajpheart.2000.279.4.H1540، PMID 11009439، S2CID 2543076.
- "Delta-like ligand 4 (Dll4) is induced by VEGF as a negative regulator of angiogenic sprouting"، Proceedings of the National Academy of Sciences of the United States of America، 104 (9): 3219–24، فبراير 2007، Bibcode:2007PNAS..104.3219L، doi:10.1073/pnas.0611206104، PMC 1805530، PMID 17296940.
- "Dll4 signalling through Notch1 regulates formation of tip cells during angiogenesis"، Nature، 445 (7129): 776–80، فبراير 2007، Bibcode:2007Natur.445..776H، doi:10.1038/nature05571، PMID 17259973، S2CID 4407198.
- Segarra M, Williams CK, Sierra ML, Bernarndo M, McCormick PJ, Meric D, Regino C, Choyke P, Tosato G. 2008. Dll4 activation of Notch signaling reduces tumore vascularity and inhibits tumor growth. Blood Journal. 112(5): 1904-1911
- "Simultaneous blockade of VEGF and Dll4 by HD105, a bispecific antibody, inhibits tumor progression and angiogenesis"، mAbs، 8 (5): 892–904، 2016، doi:10.1080/19420862.2016.1171432، PMC 4968104، PMID 27049350.
- "A perspective on the role of class III semaphorin signaling in central nervous system trauma"، Frontiers in Cellular Neuroscience، 8: 328، 2014، doi:10.3389/fncel.2014.00328، PMC 4209881، PMID 25386118.
- "Neuropilin-1 is expressed by endothelial and tumor cells as an isoform-specific receptor for vascular endothelial growth factor"، Cell، 92 (6): 735–45، مارس 1998، doi:10.1016/s0092-8674(00)81402-6، PMID 9529250، S2CID 547080.
- "VEGF binding to NRP1 is essential for VEGF stimulation of endothelial cell migration, complex formation between NRP1 and VEGFR2, and signaling via FAK Tyr407 phosphorylation"، Molecular Biology of the Cell، 22 (15): 2766–76، أغسطس 2011، doi:10.1091/mbc.E09-12-1061، PMC 3145551، PMID 21653826.
- "Angiogenesis as a therapeutic target"، Nature، 438 (7070): 967–74، ديسمبر 2005، Bibcode:2005Natur.438..967F، doi:10.1038/nature04483، PMID 16355214، S2CID 1183610.
- "Angiogenic factors"، Science، 235 (4787): 442–7، يناير 1987، Bibcode:1987Sci...235..442F، doi:10.1126/science.2432664، PMID 2432664.
- "Fighting cancer by attacking its blood supply"، Scientific American، 275 (3): 150–4، سبتمبر 1996، Bibcode:1996SciAm.275c.150F، doi:10.1038/scientificamerican0996-150، PMID 8701285.
- "[Induction of myocardial neoangiogenesis by human growth factors. A new therapeutic approach in coronary heart disease]"، Herz (باللغة الألمانية)، 25 (6): 589–99، سبتمبر 2000، doi:10.1007/PL00001972، PMID 11076317.
- "Angiogenic therapy of the human heart"، Circulation، 97 (7): 628–9، فبراير 1998، doi:10.1161/01.CIR.97.7.628، PMID 9495294، مؤرشف من الأصل في 30 مارس 2019.
- "Mathematical modelling of dynamic adaptive tumour-induced angiogenesis: clinical implications and therapeutic targeting strategies"، Journal of Theoretical Biology، 241 (3): 564–89، أغسطس 2006، Bibcode:2006JThBi.241..564M، doi:10.1016/j.jtbi.2005.12.022، PMID 16487543.
- "Mesoscopic and continuum modelling of angiogenesis"، Journal of Mathematical Biology، 70 (3): 485–532، فبراير 2015، arXiv:1401.5701، doi:10.1007/s00285-014-0771-1، PMC 5320864، PMID 24615007.
- Gonzalez-Perez RR, Rueda BR (2013)، Tumor angiogenesis regulators (ط. first)، Boca Raton: Taylor & Francis، ص. 347، ISBN 978-1-4665-8097-8، مؤرشف من الأصل في 30 ديسمبر 2021، اطلع عليه بتاريخ 02 أكتوبر 2014.
- "Tumor cells circulate in the peripheral blood of all major carcinomas but not in healthy subjects or patients with nonmalignant diseases"، Clinical Cancer Research، 10 (20): 6897–904، أكتوبر 2004، doi:10.1158/1078-0432.CCR-04-0378، PMID 15501967.
- "Use of anti-VEGF adjuvant therapy in cancer: challenges and rationale"، Trends in Molecular Medicine، 16 (3): 122–32، مارس 2010، doi:10.1016/j.molmed.2010.01.004، PMID 20189876.
- "Angiogenesis in cancer"، Vascular Health and Risk Management، 2 (3): 213–9، سبتمبر 2006، doi:10.2147/vhrm.2006.2.3.213، PMC 1993983، PMID 17326328.
- "Angiogenesis and the heart: therapeutic implications"، Journal of the Royal Society of Medicine، 90 (6): 307–11، يونيو 1997، doi:10.1177/014107689709000604، PMC 1296305، PMID 9227376.
- "Spatiotemporal control over growth factor signaling for therapeutic neovascularization"، Advanced Drug Delivery Reviews، 59 (13): 1340–50، نوفمبر 2007، doi:10.1016/j.addr.2007.08.012، PMC 2581871، PMID 17868951.
- Rouwkema, Jeroen؛ Khademhosseini, Ali (سبتمبر 2016)، "Vascularization and Angiogenesis in Tissue Engineering: Beyond Creating Static Networks"، Trends in Biotechnology، 34 (9): 733–745، doi:10.1016/j.tibtech.2016.03.002، PMID 27032730.
- Amaral, Ronaldo Jose Farias Correa؛ Cavanagh, Brenton؛ O'Brien, Fergal Joseph؛ Kearney, Cathal John (16 ديسمبر 2018)، "Platelet‐derived growth factor stabilises vascularisation in collagen‐glycosaminoglycan scaffolds"، Journal of Tissue Engineering and Regenerative Medicine، 13 (2): 261–273، doi:10.1002/term.2789، PMID 30554484، S2CID 58767660.
- Chia-Chi Chien؛ Ivan M. Kempson؛ Cheng Liang Wang؛ H. H. Chen؛ Yeukuang Hwua؛ N. Y. Chen؛ T. K. Lee؛ Kelvin K.-C. Tsai؛ Ming-Sheng Liu؛ Kwang-Yu Change؛ C. S. Yang؛ G. Margaritondo (مايو–يونيو 2013)، "Complete microscale profiling of tumor microangiogenesis"، Biotechnology Advances، 31 (3): 396–401، doi:10.1016/j.biotechadv.2011.12.001، PMID 22193280.
 بوابة طب
بوابة طب