سوط (أحياء)
السوط (ج.: سياط) (باللاتينية: flagellum) بنية سطحية لخلايا حقيقيات النوى أو بدائيات النوى تساعدها على الحركة[1][2] في الوسط السائل أو على سطح وسط جامد. توجد فروق هامة بين السياط عند حقيقيات النوى وبدائيات النوى من حيث تركيبها البروتيني وآلية الدفع والبنية، حيث ثخن السوط عند البكتيريا من 10 إلى 20 نم، ولذلك لا يمكن رؤيته بواسطة المجهر الضوئي، وطوله 3-15 ميكرون، بينما قد يصل ثخن السوط عند حقيقيات النوى إلى 200 نم وطوله إلى 200 ميكرون. وتتشابه بنية السياط والأهداب عند حقيقيات النوى، وقد تختلف بالوظيفة أو الطول.[3]
| سوط | |
|---|---|
 تركيب سوط بكتيري | |
.jpg.webp) صورة بمجهر إلكتروني ماسح لكلاميدوموناس (10000×) صورة بمجهر إلكتروني ماسح لكلاميدوموناس (10000×) | |
| تفاصيل | |
| نوع من | عضية خلوية |
| ترمينولوجيا هستولوجيكا | H1.00.01.1.01032 |
| FMA | 67472 |
| ن.ف.م.ط. | A11.284.180.290 |
| ن.ف.م.ط. | D005407 |
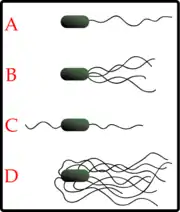
النطفة مثال على خلية حقيقية النواة تستعمل سوطها للتحرك نحو الجهاز التناسلي الأنثوي وضمنه[4]، والملويات البوابية مثال على بكتيريا ذات سياط متعددة تساعدها على الحركة عبر المخاط للوصول إلى ظهارة جهاز الهضم.[5]
تتكون من مواد بروتينية، ويسمى هذا الآحين (البروتين) فلاجيلين ويكون على شكل سلك أو سوط. وتقوم بمساعدة البكتيريا وغيرها من الخلايا على الحركة. ففي عالم البكتيريا يطلق على البكتيريا التي تمتلك سياطا بأنها متحركة. و وتعتبر السياط من الصفات المميزة للبكتيريا والتي يمكن استخدامها في عملية التصنيف.
أنواع
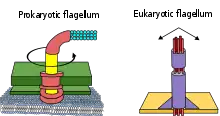
تم حتى الآن تمييز ثلاثة أنواع من الأسواط: البكتيرية، البدائية، وحقيقية النواة.
الاختلافات الرئيسية بين هذه الأنواع الثلاثة هي:
- الأسواط البكتيرية عبارة عن خيوط حلزونية، لكل منهامحرك دوار في قاعدتها يمكن أن تدور في اتجاه عقارب الساعة أو عكس اتجاه عقارب الساعة[7][8] [9][10] أنها توفر نوعين من عدة أنواع من الحركة البكتيرية.
- تتشابه الأسواط الأثرية (الأركيلا) بشكل سطحي مع الأسواط البكتيرية، ولكنها تختلف في كثير من التفاصيل وتعتبر غير متجانسة.[11][12][13]
- سوط حقيقيات النوى - تلك الخاصة بالخلايا الحيوانية، والنباتية، والخلايا الأولية - هي نتوءات خلوية معقدة تتقلب ذهابًا وإيابًا. يتم تصنيف سوط حقيقيات النوى جنبًا إلى جنب مع أهداب متحركة حقيقية النواة على أنها undulipodia للتأكيد على دورها الملحقالمتموج في الوظيفة الخلوية أو الحركة. الأهداب الأولية غير قابلة للحركة، وليست نواة؛ لديهم محور عصبي 9 + 0 مختلف هيكليًا بدلاً من محور عصبي 9 + 2 الموجود في كل من السوط والأهداب المتحركة.
جرثومي

الهيكل والتكوين
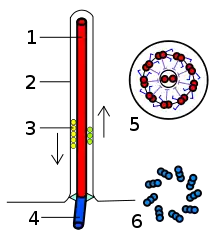
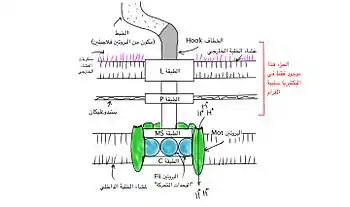
يتكون السوط البكتيري من بروتين فلاجيلين. شكله عبارة عن أنبوب مجوف بسمك 20 نانومتر. إنه حلزوني وله منحنى حاد خارج الغشاء الخارجي؛ يسمح هذا «الخطاف» لمحور اللولب بالإشارة بعيدًا عن الخلية مباشرةً. يمتد عمود بين الخطاف والجسم القاعدي، ويمر عبر حلقات البروتين في غشاء الخلية التي تعمل كمحامل. الكائنات موجبة الجرام لها اثنتان من حلقات الجسم القاعدية هذه، واحدة في طبقة الببتيدوغليكان والأخرى في غشاء البلازما.الكائنات سالبة الجرام لها أربع حلقات من هذا القبيل: الحلقة L ترتبط بـعديدات السكاريد الدهنية، ترتبط الحلقة Pبطبقة الببتيدوغليكان، والحلقة M مضمنة في غشاء البلازما، والحلقة S متصلة مباشرة بغشاء البلازما. ينتهي الخيط ببروتين متدرج.[14]
الخيط السوطي هو المسمار اللولبي الطويل الذي يدفع البكتيريا عند تدويرها بواسطة المحرك، من خلال الخطاف. في معظم البكتيريا التي تمت دراستها، بما في ذلك الإشريكية القولونية سالبة الجرام ، السالمونيلا التيفية، Caulobacter crescentus ، و Vibrio alginolyticus ، يتكون الخيط من 11 خيطًا أوليًا متوازيًا تقريبًا مع محور الشعيرة. كل خيوط أولية عبارة عن سلسلة من سلاسل البروتين الترادفية. ومع ذلك، تحتوي العطيفة الصائميةعلى سبعة خيوط أولية.
يمتلك الجسم القاعدي العديد من السمات المشتركة مع بعض أنواع المسام الإفرازية، مثل «السدادة» المجوفة التي تشبه القضيب في مراكزها الممتدة عبر غشاء البلازما. تقدم أوجه التشابه بين الأسواط البكتيرية وبنى النظام الإفرازي البكتيري والبروتينات أدلة علمية تدعم النظرية القائلة بأن الأسواط البكتيرية تطورت من نظام الإفراز من النوع الثالث.
محرك
مزيد من المعلومات: الحركة الدوارة في الأنظمة الحية
يتم تشغيل السوط البكتيري بواسطة محرك دوار (مجمع Mot) يتكون من البروتين، الموجود عند نقطة تثبيت السوط على غشاء الخلية الداخلي. يتم تشغيل المحرك بواسطة القوة المحركة للبروتون، أي عن طريق تدفق البروتونات (أيونات الهيدروجين) عبر غشاء الخلية البكتيرية بسبب تدرج تركيز تم إعداده بواسطة عملية التمثيل الغذائي للخلية (أنواع Vibrio لها نوعان من السوط، الجانبي والقطبي، ويتم تشغيل بعضها بواسطة مضخة أيونات الصوديوم بدلاً من مضخة البروتون ).ينقل الجزء المتحرك البروتونات عبر الغشاء، ويتم قلبه في هذه العملية. يمكن أن يعمل الدوار وحده من 6000 إلى 17000 دورة في الدقيقة، ولكن مع الخيوط السوطية المرفقة عادة ما تصل فقط إلى 200 إلى 1000 دورة في الدقيقة. يمكن تغيير اتجاه الدوران بواسطة مفتاح المحرك السوطي بشكل فوري تقريبًا، بسبب تغيير طفيف في موضع البروتين، FliG ، في الدوار. السوط ذو كفاءة عالية في استخدام الطاقة ويستخدم القليل جدًا من الطاقة. لا تزال الآلية الدقيقة لتوليد عزم الدوران غير مفهومة جيدًا. نظرًا لعدم وجود مفتاح تشغيل وإيقاف للمحرك السوطي، يتم استخدام البروتين epsE كقابض ميكانيكي لفصل المحرك عن الدوار، وبالتالي إيقاف السوط والسماح للبكتيريا بالبقاء في مكان واحد.
يناسب الشكل الأسطواني للأسواط حركة الكائنات المجهرية؛ تعمل هذه الكائنات الحية بعدد رينولدز المنخفض، حيث تكون لزوجة المياه المحيطة أكثر أهمية من كتلتها أو قصورها.
تختلف سرعة دوران الأسواط وفقًا لشدة القوة الدافعة للبروتون، مما يسمح بأشكال معينة من التحكم في السرعة، كما يسمح لبعض أنواع البكتيريا بالحصول على سرعات ملحوظة تتناسب مع حجمها؛ يصل بعضها إلى 60 خلية في الثانية تقريبًا. بهذه السرعة، تستغرق البكتيريا حوالي 245 يومًا لتغطي مسافة كيلومتر واحد؛ على الرغم من أن هذا قد يبدو بطيئًا، إلا أن المنظور يتغير عندما يتم تقديم مفهوم المقياس. بالمقارنة مع أشكال الحياة المجهرية، فهي سريعة جدًا بالفعل عند التعبير عنها من حيث عدد أطوال الجسم في الثانية. على سبيل المثال، يبلغ طول الفهد حوالي 25 طولًا للجسم في الثانية.[15]
من خلال استخدام سياط، و كولاي هي قادرة على التحرك بسرعة نحو جاذبة وبعيدا عن المواد الطاردة، عن طريقالمشي العشوائي منحازة، مع «يعمل» و «السقوط» التي أحدثتها الدورية لها السوط عكس اتجاه عقارب الساعة وعقارب الساعة، على التوالي. اتجاهي الدوران غير متطابقين (فيما يتعلق بحركة السوط) ويتم تحديدهما بواسطة مفتاح جزيئي.[16]
المجسم
أثناء تجميع السوط، تمر مكونات السوط عبر النوى المجوفة للجسم القاعدي والخيوط الناشئة. أثناء التجميع، تضاف مكونات البروتين عند الطرف السوطي بدلاً من القاعدة. في المختبر ، تتجمع الخيوط السوطية تلقائيًا في محلول يحتوي على سوط نقي كبروتين وحيد.[17]
تطور
تشترك 10 مكونات بروتينية على الأقل في السوط البكتيري في البروتينات المتماثلة مع نظام الإفراز من النوع الثالث (T3SS)، ومن ثم من المحتمل أن أحدهما قد تطور عن الآخر. نظرًا لأن T3SS يحتوي على عدد مماثل من المكونات مثل الجهاز السوطي (حوالي 25 بروتينًا)، فمن الصعب تحديد أي واحد تم تطويره أولاً. ومع ذلك، يبدو أن النظام السوطي يشتمل على المزيد من البروتينات بشكل عام، بما في ذلك العديد من المنظمين والمرافقين، ومن ثم فقد قيل أن الأسواط تطورت من T3SS. ومع ذلك، فقد تم اقتراح أن السوط ربما يكون قد تطور أولاً أو أن الهيكلين قد تطور بشكل متوازٍ. حاجة الكائنات الحية وحيدة الخلية المبكرة إلى الحركة يدعم (التنقل) أنه سيتم اختيار الأسواط الأكثر حركة من خلال التطور أولاً، ولكن يمكن اعتبار T3SS المتطور من السوط «تطورًا اختزاليًا»، ولا يتلقى أي دعم طوبولوجي من الأشجارالتطورية . الفرضية القائلة بأن الهيكلين تطورتا بشكل منفصل عن سلف مشترك تفسر أوجه التشابه بين البروتينات بين الهيكلين، بالإضافة إلى تنوعها الوظيفي.
فلاجيلا ومناقشة التصميم الذكي
جادل بعض المؤلفين بأن الأسواط لا يمكن أن تتطور، بافتراض أنها لا تستطيع العمل بشكل صحيح إلا عندما تكون جميع البروتينات في مكانها. وبعبارة أخرى، فإن الجهاز السوطي «معقد بشكل غير قابل للاختزال». ومع ذلك، يمكن حذف العديد من البروتينات أو تحويرها ولا يزال الجلد يعمل، وإن كان في بعض الأحيان بكفاءة منخفضة. بالإضافة إلى ذلك، يتنوع تكوين الأسواط بشكل مدهش عبر البكتيريا، حيث توجد العديد من البروتينات فقط في بعض الأنواع، ولكن ليس في البعض الآخر. ومن ثم، فمن الواضح أن الجهاز السوطي مرن جدًا من الناحية التطورية وقادرًا تمامًا على فقدان أو اكتساب مكونات البروتين. على سبيل المثال، تم العثور على عدد من الطفرات التي تزيد من حركية الإشريكية القولونية. دليل إضافي على تطور الأسواط البكتيرية يتضمن وجود سوط أثري، وأشكال وسيطة من الأسواط وأنماط من التشابه بين متواليات البروتين السوطي، بما في ذلك ملاحظة أن جميع البروتينات السوطية الأساسية تقريبًا قد عرفت تماثلات مع بروتينات غير سوطية . علاوة على ذلك، تم تحديد العديد من العمليات على أنها تلعب أدوارًا مهمة في تطور السوط، بما في ذلك التجميع الذاتي للوحدات الفرعية المتكررة البسيطة، والازدواجية الجينية مع الاختلاف اللاحق، وتجنيد عناصر من أنظمة أخرى («bricolage الجزيئي») وإعادة التركيب.[18]
مخططات ترتيب فلاجيلار
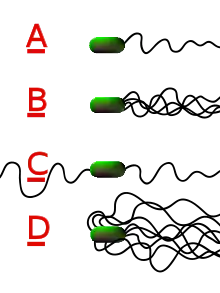
الأنواع المختلفة من البكتيريا لها أعداد وترتيبات مختلفة من الأسواط.
- تحتوي البكتيريا أحادية الشعر على سوط واحد (على سبيل المثال، ضمة الكوليرا).
- تحتوي البكتيريا Lophotrichous على العديد من الأسواط الموجودة في نفس المكان على الأسطح البكتيرية والتي تعمل في تناسق لدفع البكتيريا في اتجاه واحد. في كثير من الحالات، تكون قواعد الأسواط المتعددة محاطة بمنطقة متخصصة من غشاء الخلية، تسمى العضية القطبية .
- تمتلك البكتيريا البرمائية سوطًا واحدًا على كل من الطرفين المتقابلين (يعمل سوط واحد فقط في كل مرة، مما يسمح للبكتيريا بعكس مسارها بسرعة عن طريق تبديل السوط النشط).
- للبكتيريا peritrichous سوط بارزة في جميع الاتجاهات (على سبيل المثال، E. coli).
في أشكال معينة كبيرة من السلينوموناس ، يتم تنظيم أكثر من 30 سوطًا فرديًا خارج جسم الخلية، حيث يتم التواء حلزونيًا حول بعضها البعض لتشكيل بنية سميكة (يمكن رؤيتها بسهولة بواسطة المجهر الضوئي) تسمى «حافظة».
على النقيض من ذلك، تحتوي اللولبيات على سوط ينشأ من أقطاب متقابلة للخلية، وتقع داخل الفضاء المحيطي كما يتضح من كسر الغشاء الخارجي ومؤخرًا بواسطة الفحص المجهري الإلكتروني . يؤدي دوران الخيوط بالنسبة لجسم الخلية إلى تحرك البكتيريا بأكملها للأمام في حركة تشبه المفتاح، حتى من خلال مادة لزجة بدرجة كافية لمنع مرور البكتيريا ذات الجلد الطبيعي.
يؤدي الدوران بعكس اتجاه عقارب الساعة لسوط قطبي أحادي إلى دفع الخلية إلى الأمام مع تأخر السوط خلفها، تمامًا مثل المفتاح الذي يتحرك داخل الفلين. في الواقع، الماء على النطاق المجهري شديد اللزوجة، ويختلف تمامًا عن تجربتنا اليومية للمياه.
فلاجيللا عبارة عن حلزونات أعسر، ويتم تجميعها وتدويرها معًا فقط عند الدوران عكس اتجاه عقارب الساعة. عندما تنعكس بعض الدوارات في الاتجاه، تنفصل السوط وتبدأ الخلية في «الانقلاب». حتى لو كانت جميع الأسواط تدور في اتجاه عقارب الساعة، فمن المحتمل ألا تشكل حزمة، بسبب أسباب هندسية، وكذلك لأسباب هيدروديناميكية. قد يحدث مثل هذا «التقلب» في بعض الأحيان، مما يؤدي إلى تحطم الخلية على ما يبدو في مكانها، مما يؤدي إلى إعادة توجيه الخلية. يتم منع دوران السوط في اتجاه عقارب الساعة بواسطة مركبات كيميائية مواتية للخلية (مثل الطعام)، ولكن المحرك شديد التكيف مع هذا. لذلك، عند التحرك في اتجاه مواتٍ، يزداد تركيز الجاذب الكيميائي ويتم قمع «السقوط» باستمرار؛ ومع ذلك، عندما يكون اتجاه حركة الخلية غير مواتٍ (على سبيل المثال، بعيدًا عن جاذب كيميائي)، لا يتم قمع الانهيارات وتحدث كثيرًا، مع احتمال إعادة توجيه الخلية في الاتجاه الصحيح.
في بعض Vibrio spp. (بشكل خاص Vibrio parahaemolyticus ) والبكتيريا البروتينية ذات الصلة مثل Aeromonas ، يتعايش نظامان سوطيان، باستخدام مجموعات مختلفة من الجينات وتدرجات أيونية مختلفة للطاقة. يتم التعبير عن الأسواط القطبية بشكل أساسي وتوفر الحركة في سائل الكتلة، في حين يتم التعبير عن السوط الجانبي عندما يواجه السوط القطبي مقاومة كبيرة للغاية. توفر هذه الحشود الحركية على الأسطح أو في السوائل اللزجة.
عتيق
إن البدائية التي يمتلكها بعض الأرخبيل تشبه بشكل سطحي السوط البكتيري. في الثمانينيات، كان يُعتقد أنهم متماثلون على أساس التشكل والسلوك الإجمالي. يتكون كل من الأسواط والأركيلا من خيوط تمتد خارج الخلية وتدور لدفع الخلية. السوط الأثري له هيكل فريد يفتقر إلى قناة مركزية.على غرار البلينات البكتيرية من النوع الرابع، فإن السوطات الأثرية (الأركايلينات) مصنوعة من ببتيدات الإشارة من الفئة 3 وتتم معالجتها بواسطة إنزيم يشبه الببتيداز من النوع الرابع. وعادة ما يتم تعديل archaellins بإضافة المرتبطة N glycans والتي هي ضرورية لتجميع السليم أو وظيفة.
كشفت الاكتشافات في التسعينيات عن العديد من الاختلافات التفصيلية بين الأسواط البدائية والبكتيرية. وتشمل هذه:
- مزودة بمحركات سياط البكتيرية عن طريق تدفق H +أيونات (أو أحيانا نا + الأيونات)؛ يكاد يكون من المؤكد تقريبا أن السوط الأثري مدعوم من ATP . وعزم الدوران لم يتم التعرف -generating الحركية التي القوى دوران السوط archaeal.
- في حين أن الخلايا البكتيرية غالبًا ما تحتوي على العديد من الخيوط السوطية، والتي يدور كل منها بشكل مستقل، فإن السوط البدائي يتكون من حزمة من العديد من الخيوط التي تدور كتجمع واحد.
- تنمو الأسواط البكتيرية عن طريق إضافة وحدات فرعية من السوط في الطرف ؛ تنمو الأسواط الأثرية عن طريق إضافة وحدات فرعية إلى القاعدة.
- الأسواط البكتيرية أكثر سمكًا من الأركيلا، والخيوط البكتيرية بها «أنبوب» مجوف كبير بالداخل بحيث يمكن للوحدات الفرعية السوطية أن تتدفق إلى داخل الشعيرة وتضاف عند الطرف ؛ الأركيلوم رقيق جدًا (12-15 نانومتر) للسماح بذلك.
- تشترك العديد من مكونات الأسواط البكتيرية في تشابه التسلسل مع مكونات أنظمة الإفراز من النوع الثالث، لكن مكونات الأسواط البكتيرية والعرقية لا تشترك في أي تشابه في التسلسل. بدلاً من ذلك، تشترك بعض مكونات الأركيلا في التسلسل والتشابه المورفولوجي مع مكوناتالنوع الرابع من الشعيرات، والتي يتم تجميعها من خلال عمل أنظمة إفراز النوع الثاني (لا تتسق تسمية أنظمة إفراز البروتين والبروتين).
ويمكن لهذه الخلافات يعني أن سياط البكتيرية وarchaella يمكن أن يكون حالة كلاسيكية من البيولوجي القياس، أو تطور متقاربة ، وليس التماثل . ومع ذلك ، بالمقارنة مع عقود من الدراسة التيحظيت بتغطية إعلامية جيدة للأسواط البكتيرية (على سبيل المثال بواسطة هوارد بيرج)، لم تظهر الأركيلا إلا مؤخرًا بدأت في جذب الاهتمام العلمي.
حقيقيات النوى
المصطلح
بهدف التأكيد على التمييز بين الأسواط البكتيرية والأهدابحقيقية النواة والأسواط ، حاول بعض المؤلفين استبدال اسم هاتين البُنى حقيقية النواة بـ " undulipodia " (على سبيل المثال ، جميع الأوراق التي كتبها Margulis منذ السبعينيات) أو "cilia" لكليهما (على سبيل المثال ، Hülsmann ، 1992 ؛ Adl et al. 2012 ؛ معظم أوراق Cavalier-Smith)، والحفاظ على «الأسواط» للبنية البكتيرية. ومع ذلك ، فإن الاستخدام التمييزي لمصطلحي «أهداب» و «سوط» لحقيقيات النوى المعتمدين في هذه المقالة لا يزال شائعًا (على سبيل المثال ، أندرسن وآخرون ، 1991 ؛ ليدبيتر وآخرون ، 2000).[19]
الهيكل الداخلي
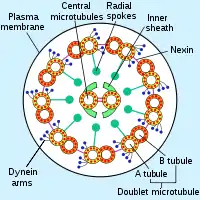
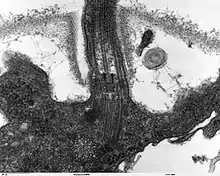
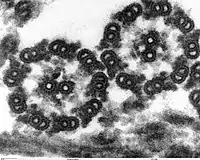
السوط حقيقيات النوى عبارة عن حزمة من تسعة أزواجمدمجة من مضاعفات الأنابيب الدقيقة المحيطة بنبيبيين مركزيين منفردين. البنية المسماة "9 + 2" هي سمة من سمات جوهر سوط حقيقيات النوى المسمى محور عصبي .يوجد في قاعدة سوط حقيقيات النوى جسم قاعدي ، «الجفن» أو الجسيم الحركي ، وهو مركز تنظيم الأنابيب الدقيقة للأنابيب الدقيقة السوطية ويبلغ طوله حوالي 500 نانومتر. الأجسام القاعدية متطابقة هيكليا مع المريكزات .يتم تغليف السوط داخل غشاء البلازما للخلية ، بحيث يمكن الوصول إلى الجزء الداخلي من السوط من خلال سيتوبلازمالخلية .
إلى جانب الجسم المحوري والجسم القاعدي ، الثابت نسبيًا في التشكل ، فإن الهياكل الداخلية الأخرى للجهاز السوطي هي منطقة الانتقال (حيث يلتقي الجسم المحوري والجسم القاعدي) ونظام الجذر (الهياكل الدقيقة أو الليفية التي تمتد من الأجسام القاعدية إلى السيتوبلازم)، أكثر تنوعًا وفائدة كمؤشرات على العلاقات التطورية لحقيقيات النوى. الهياكل الأخرى ، الأكثر شيوعًا ، هي قضيب paraflagellar (أو paraxonemal)، والألياف R ، والألياف S. السطحية ، انظر أدناه للتراكيب.[20]:63–84
آلية
يمتد كل من الأنابيب الدقيقة الخارجية التسعة المزدوجة زوجًا من أذرع داينين (ذراع «داخلي» و «خارجي») إلى الأنابيب الدقيقة المجاورة ؛ هذه تنتج القوة من خلال التحلل المائي ATP. يحتوي المحوار السوطي أيضًا على مكبرات صوت شعاعية ، ومجمعات متعددة الببتيد تمتد من كل من المضاعفات التسعة للأنابيب الخارجية نحو الزوج المركزي ، مع «رأس» المتحدث مواجهًا للداخل. يُعتقد أن المتحدث الشعاعي متورط في تنظيم حركة السوط ، على الرغم من أن وظيفته وطريقة عمله لم يتم فهمهما بعد.
فلاجيلا مقابل أهداب
نمط الضرب من «سوط» و «سيلوم» حقيقيات النوى ، وهو تمييز تقليدي قبل معرفة بنية الاثنين.
تولد أنماط الضرب المنتظمة لأهداب حقيقية النواة والأسواط حركة على المستوى الخلوي. تتراوح الأمثلة من دفع الخلايا المفردة مثل سباحة الحيوانات المنوية إلى نقل السائل على طول طبقة ثابتة من الخلايا كما هو الحال في الجهاز التنفسي .
على الرغم من أن أهداب وسوط حقيقيات النوى متماثلان في النهاية ، إلا أنهما يُصنفان أحيانًا حسب نمط حركتهما ، وهو تقليد يعود إلى ما قبل معرفة هياكلها. في حالة السوط ، غالبًا ما تكون الحركة مستوية وشبيهة بالموجة ، بينما تؤدي الأهداب المتحركة غالبًا حركة ثلاثية الأبعاد أكثر تعقيدًا بضربة طاقة واسترداد. شكل تقليدي آخر للتمييز هو عدد 9 + 2 عضيات على الخلية.
النقل
النقل داخل السوط ، العملية التي يتم من خلالها نقل الوحدات الفرعية المحورية ، والمستقبلات عبر الغشاء ، والبروتينات الأخرى لأعلى ولأسفل على طول السوط ، ضرورية لأداء السوط بشكل صحيح ، في كل من الحركة ونقل الإشارة.
التطور والوقوع
سياط حقيقية النواة أو أهداب، وربما سمة الأجداد، وعلى نطاق واسع في جميع الفئات تقريبا من حقيقيات النوى، كشرط الدائمة نسبيا، أو كمرحلة دورة الحياة سوطي (على سبيل المثال، zoids ، الأمشاج ، الأبواغ الحيوانية السابحة ، والتي يمكن أن تنتج باستمرار أو ليس).
وجدت على الحالة الأولى إما في خلايا متخصصة من الكائنات متعددة الخلايا (على سبيل المثال، choanocytes من الإسفنج أو مهدبة ظهائر منmetazoans)، كما هو الحال في ciliates والعديد من حقيقيات النوى مع «حالة سوطية» (أو «monadoidمستوى التنظيم»، انظر فلاجيلاتا ، مجموعة اصطناعية).
توجد مراحل دورة حياة سوطي في العديد من المجموعات، على سبيل المثال، العديد من الطحالب الخضراء (الأبواغ الحيوانية السابحة والأمشاج الذكرية)، الطحلبيات (الأمشاج الذكرية)، البتيريدوفيت (الأمشاج الذكور)، وبعض عاريات البذور (السيكاسيات و الجنكة ، والأمشاج الذكرية)، تتمحور الدياتومات (الأمشاج الذكور)، الطحالب البني (الأبواغ الحيوانية السابحة والأمشاج)، الفطريات البيضية (الأبواغ الحيوانية السابحة assexual والأمشاج)، hyphochytrids (الأبواغ الحيوانية السابحة)، labyrinthulomycetes (الأبواغ الحيوانية السابحة)، وبعض apicomplexans (الأمشاج)، وبعض الراديولاريا (ربما الأمشاج)، المنخريات (الأمشاج)، plasmodiophoromycetes (الأبواغ الحيوانية السابحة والأمشاج)، myxogastrids (الأبواغ الحيوانية السابحة)، metazoans (الأمشاج الذكور)، وحيدة الخلية الفطريات (الأبواغ الحيوانية السابحة والأمشاج).
لا توجد الأسواط أو الأهداب تمامًا في بعض المجموعات ، ربما بسبب الخسارة بدلاً من كونها حالة بدائية. وقعت فقدان أهداب في الطحالب الحمراء ، وبعض الطحالب الخضراء (طحالب زيغنيمية)، وعاريات البذور باستثناء السيكاسيات و الجنكة ، كاسيات ، pennate الدياتومات ، وبعض apicomplexans ، بعض amoebozoans ، في الحيوانات المنوية من بعض metazoans ، والفطريات (باستثناء chytrids).
التصنيف
يتم استخدام عدد من المصطلحات المتعلقة بالسوط أو الأهداب لوصف حقيقيات النوى. وفقًا السطحية الموجودة ، قد يكون الأسواط:
- السوط المصع (= سوط أملس ، لا كرونماتي): بدون شعر ، على سبيل المثال ، في Opisthokonta
- سوط مشعر (= بهرج ، وميض ، سوط الجنبة): مع الشعر (= mastigonemes sensu lato)، مقسم إلى:
- ذات الشعر الناعم (= الشعر غير الأنبوبي أو البسيط): يحدث في Euglenophyceae ، Dinoflagellata ، بعض Haptophyceae (Pavlovales)
- ذات الشعر القاسي (= الشعر الأنبوبي ، الريترونيمات ، الضرع بالمعنى الضيق)، مقسم إلى:
- الشعر ثنائي الجزء: ذو منطقتين. يحدث فيCryptophyceae ، Prasinophyceae ، وبعض سوطيات متغايرة
- شعر ثلاثي (= سترامينيبيلوس): ذو ثلاث مناطق (قاعدة ، عمود أنبوبي ، وشعر طرفي واحد أو أكثر). يحدث في معظمHeterokonta
- سوط عظمي: مع صف واحد من الشعر
- سوط بانتونيماتيك: مع صفين من الشعر
- acronematic: سوط بشعر واحد أو ضرع طرفي أو شعر سوطي (على سبيل المثال ، bodonids)؛ يستخدم بعض المؤلفين هذا المصطلح كمرادف للجلد
- مع المقاييس: على سبيل المثال ، Prasinophyceae
- مع أشواك: على سبيل المثال ، بعض الطحالب البنية
- مع غشاء متموجة: على سبيل المثال، بعض ذوات منشأ الحركة ، وبعض parabasalids
- مع خرطوم (نتوء يشبه الجذع للخلية): على سبيل المثال ، apusomonads ، بعض bodonids
وفقًا لعدد الأسواط ، قد تكون الخلايا (تذكر أن بعض المؤلفين يستخدمون «مهدب» بدلاً من «جلد»:
- uniflagellated: على سبيل المثال ، معظم Opisthokonta
- biflagellated: على سبيل المثال، كل دواميات السياط، الأمشاج من طحالب كارية ، معظم النباتات الطحلبية وبعض metazoans
- triflagellated: على سبيل المثال ، الأمشاج لبعض فورامينيفيرا
- quadriflagellated: على سبيل المثال ، بعض Prasinophyceae ، Collodictyonidae
- octoflagellated: على سبيل المثال ، بعض Diplomonada ، بعض Prasinophyceae
- multiflagellated: على سبيل المثال ، Opalinata ، Ciliophora ، Stephanopogon ، Parabasalida ، Hemimastigophora ، Caryoblastea ، Multicilia ، الأمشاج (أو zoids) من Oedogoniales (Chlorophyta)، وبعض النباتاتpteridophytes وبعض عاريات البذور
وفقًا لمكان إدخال السوط:
- opisthokont: الخلايا ذات السوط الذي تم إدخاله في الخلف ، على سبيل المثال ، في Opisthokonta (Vischer ، 1945). في Haptophyceae ، يتم إدخال الأسواط بشكل جانبي إلى النهاية ، ولكن يتم توجيهها للخلف أثناء السباحة السريعة.
- akrokont: خلايا مع سوط يتم إدخالها بشكل قمي
- subakrokont: خلايا ذات سوط مُدرج تحت السوط
- pleurokont: خلايا ذات سوط يتم إدخالها جانبياً
حسب نمط الضرب:
- مزلق: سوط يتدحرج على الركيزة
- ديناميكية غير متجانسة: سوط ذو أنماط مختلفة من الضرب (عادة مع سوط يعمل في التقاط الطعام والآخر يعمل في الانزلاق أو التثبيت أو الدفع أو «التوجيه»)
- إيزوديناميكي: الضرب بالسوط بنفس الأنماط
المصطلحات الأخرى المتعلقة بنوع السوط:
- isokont: خلايا ذات أسواط متساوية الطول. كما تم استخدامه سابقًا للإشارة إلى الكلوروفيتا
- anisokont: الخلايا مع سياط من عدم المساواة في طول، على سبيل المثال، بعض من الطحالب وPrasinophyceae
- heterokont: مصطلح قدمه لوثر (1899) للإشارة إلى Xanthophyceae ، بسبب زوج من الأسواط ذات الطول غير المتكافئ. لقد اتخذ معنى محددًا في الإشارة إلى الخلايا ذات السوط الأمامي من سترامينيبلوس (مع أضلاع الضرع الثلاثية ، في صف واحد أو صفين) وسوط خلفي عادة ما يكون أملس. كما أنها تستخدم للإشارة إلى الصنف Heterokonta
- stephanokont: الخلايا ذات تاج السوط بالقرب من نهايتها الأمامية ، على سبيل المثال ، الأمشاج وجراثيم Oedogoniales ، جراثيم بعض Bryopsidales . المصطلح الذي قدمه Blackman & Tansley (1902) للإشارة إلى Oedogoniales
- akont: خلايا بدون أسواط. كما كان يستخدم للإشارة إلى مجموعات تصنيفية، كما Aconta أو Akonta: وطحالب زيغنيمية والعصوية (أولتمان، 1904)، أوRhodophyceae (كريستنسن، 1962)
تركيب السوط الأساسي
وظيفة السياط
تساعد الخلية على الحركة حيث تدفعها للانتقال. فهي على سبيل المثال توجد في النطفة، كما توجد لدى الخلية البكتيرية.
السياط والبكتيريا
في الخلية البكتيرية توجد السياط بأنماط متعددة:
- فقد تحيط السياط بالخلية من كل اتجاه وفي هذه الحالة تسمى مُحيطِيَّةُ السِّياط (بالإنجليزية: peritrichous bacteria)
- أو قد تكون السياط متجمعة في مكان واحد من الخلية البكتيرية وتوصف بهذه الحالة بأنها عُفْرَيَّةِ السِّياط (بالإنجليزية: lophotrichous bacterium)
- وقد يتصل بخلية البكتيريا سوت واحد فقط على اتجاهين مختلفين وفي هذه الخلية تسمى مُتَقابِلُ السِّياطِ (بالإنجليزية: amphitrichous)
- وقد يوجد على خلية البكتيريا سوت واحد فقط وفي جهة واحدة فقط وفي هذه الحالة تسمى أُحادِيُّ السَّوط (بالإنجليزية: monotrichous bacterium.)
المراجع
- Bardy S.L., Ng S.Y., Jarrell K.F. (2003)، "Prokaryotic motility structures"، Microbiology، 149 (Pt 2): 295–304، doi:10.1099/mic.0.25948-0، PMID 12624192.
{{استشهاد بدورية محكمة}}: الوسيط غير المعروف|quotes=تم تجاهله (مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Lefebvre P.A. (2001)، "Assembly and motility of eukaryotic cilia and flagella. Lessons from Chlamydomonas reinhardtii"، Plant Physiology، 127 (4): 1500–1507، doi:10.1104/pp.010807، PMID 11743094.
{{استشهاد بدورية محكمة}}: الوسيط غير المعروف|quotes=تم تجاهله (مساعدة) - Haimo L.T., Rosenbaum J.L. (1981)، "Cilia, flagella, and microtubules"، Journal of Cell Biology، 91 (3 Pt 2): 125s–130s، doi:10.1083/jcb.91.3.125s، PMID 6459327.
{{استشهاد بدورية محكمة}}: الوسيط غير المعروف|quotes=تم تجاهله (مساعدة) - Malo A.F.؛ وآخرون (2006)، "Sperm design and sperm function"، Biology Letters، 2 (2): 246–9، doi:10.1098/rsbl.2006.0449، PMID 17148374، مؤرشف من الأصل في 7 فبراير 2020.
{{استشهاد بدورية محكمة}}: Explicit use of et al. in:|مؤلف=(مساعدة)، الوسيط غير المعروف|quotes=تم تجاهله (مساعدة) - Lacy B.E., Rosemore J. (2001)، "Helicobacter pylori: ulcers and more: the beginning of an era"، Journal of Nutrition، 131 (10): 2789S–93S، PMID 11584108، مؤرشف من الأصل في 03 أبريل 2010.
{{استشهاد بدورية محكمة}}: الوسيط غير المعروف|quotes=تم تجاهله (مساعدة) - Streif S, Staudinger WF, Marwan W, Oesterhelt D (2008)، "Flagellar rotation in the archaeon Halobacterium salinarum depends on ATP"، Journal of Molecular Biology، 384 (1): 1–8، doi:10.1016/j.jmb.2008.08.057، PMID 18786541.
- Meister GL, Berg HC (1987)، "Rapid rotation of flagellar bundles in swimming bacteria"، Nature، 325 (6105): 637–640، Bibcode:1987Natur.325..637L، doi:10.1038/325637a0، S2CID 4242129.
- Berg HC, Anderson RA (أكتوبر 1973)، "Bacteria swim by rotating their flagellar filaments"، Nature، 245 (5425): 380–2، Bibcode:1973Natur.245..380B، doi:10.1038/245380a0، PMID 4593496، S2CID 4173914.
- Jahn TL, Bovee EC (1965)، "Movement and locomotion of microorganisms"، Annual Review of Microbiology، 19: 21–58، doi:10.1146/annurev.mi.19.100165.000321، PMID 5318439.
- Harshey RM (2003)، "Bacterial motility on a surface: many ways to a common goal"، Annual Review of Microbiology، 57: 249–73، doi:10.1146/annurev.micro.57.030502.091014، PMID 14527279.
- Ng SY, Chaban B, Jarrell KF (2006)، "Archaeal flagella, bacterial flagella and type IV pili: a comparison of genes and posttranslational modifications"، Journal of Molecular Microbiology and Biotechnology، 11 (3–5): 167–91، doi:10.1159/000094053، PMID 16983194، S2CID 30386932.
- Metlina AL (نوفمبر 2004)، "Bacterial and archaeal flagella as prokaryotic motility organelles"، Biochemistry. Biokhimiia، 69 (11): 1203–12، doi:10.1007/s10541-005-0065-8، PMID 15627373، S2CID 632440.
- Jarrell K (2009)، "Archaeal Flagella and Pili"، Pili and Flagella: Current Research and Future Trends، Caister Academic Press، ISBN 978-1-904455-48-6.
- Diószeghy Z, Závodszky P, Namba K, Vonderviszt F (يونيو 2004)، "Stabilization of flagellar filaments by HAP2 capping"، FEBS Letters، 568 (1–3): 105–9، doi:10.1016/j.febslet.2004.05.029، PMID 15196929، S2CID 33886010.
- Hildebrand M (نوفمبر 1959)، "Motions of the running Cheetah and Horse"، Journal of Mammalogy، 44 (4): 481–495، doi:10.2307/1376265، JSTOR 1376265. Although according to Hunter, Luke؛ Hamman, Dave (2003)، Cheetah، Struik Publishers، ص. 37–38،
the cheetah's fastest recorded speed was 110 كم/س (68 ميل/س)
- Meadows R (مايو 2011)، "How bacteria shift gears"، PLOS Biology، 9 (5): e1001061، doi:10.1371/journal.pbio.1001061، PMC 3091840، PMID 21572986.
- Asakura S, Eguchi G, Iino T (أكتوبر 1964)، "Reconstitution of Bacterial Flagella In Vitro"، Journal of Molecular Biology، 10: 42–56، doi:10.1016/S0022-2836(64)80026-7، PMID 14222895.
- Pallen MJ, Gophna U (2007)، "Bacterial flagella and Type III secretion: case studies in the evolution of complexity"، Genome Dynamics، 3: 30–47، doi:10.1159/000107602، ISBN 978-3-8055-8340-4، PMID 18753783.
- Leadbeater, Barry S. C.؛ Green, John C., المحررون (2000)، Flagellates: Unity, Diversity and Evolution، The Systematics Association Special Volume، Taylor and Francis، ج. 59، ISBN 978-1-4822-6822-5، مؤرشف من الأصل في 13 أغسطس 2020.
- Barsanti L, Gualtieri P (2006)، Algae: Anatomy, Biochemistry, and Biotechnology، Florida, USA: CRC Press، ISBN 9780203492598، مؤرشف من الأصل في 30 ديسمبر 2020.
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي.svg.png.webp) بوابة علم الأحياء الدقيقة
بوابة علم الأحياء الدقيقة
| عن عوالق |
|  | |||
|---|---|---|---|---|---|
| عن الحجم |
| ||||
| عوالق بكتيرية |
| ||||
| عوالق نباتية |
| ||||
| عوالق حيوانية |
| ||||
| related topics |
| ||||