جسيم مركزي
في علم الأحياء الخلوي، الجسيم المركزي (بالإنجليزية: Centrosome) هو عضو هيولي صغير موجود في معظم وليس كل الخلايا الحيوانية، وهو في العادة قريب من النواة إلى جانب المركز الهندسي للخلية، ويظهر كمنطقة كروية بهيولي صافية تحتوي في مركزها على نويتين، لها ألفة خاصة هيماتوكسلين الحديد (هايدن هاين) وخلال الانقسام فإن الجسيم المركزي ينقسم إلى جزأين يهاجران إلى الأقطاب المتقابلة للمغزل اللالوني، وهناك يحاط كل جزء بمنطقة واسعة صافية تسمى الكرة المركزية التي بدورها تندمج خارجيا إلى الكرة النجمية، والمعرفة حول التركيب الكيماوي، أو الوظيفة الدقيقة لهذه الهياكل المختلفة ما زالت قليلة.[1][2]
| علم الأحياء الخلوي | |
|---|---|
| الخلية الحيوانية | |
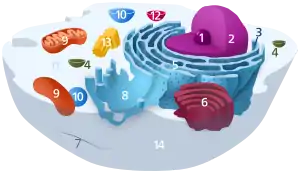 مكونات الخلية الحيوانية النموذجية:
| |
حيث يوجد في الخلية الحيوانية. وبالرغم من عدم وجود الجسيمات المركزية في الخلايا النباتيه الا انه يوجد مركز لتخليق النبيبات الدقيقة وايضا خيوط دقيقه كما ان الجسيمات المركزية متكونه من زوج من المريكزات عادة وكل من هذه المريكزات يمثل اسطوانه مكونه من تسع مجاميع وتحتوي كل مجموعه على ثلاثه نبيبات دقيقه.
التاريخ
اكتُشف الجسيم المركزي بالمشاركة بين والثر فليمينغ في 1875 وإدوارد فان بينيدين في 1876، ووصفه لاحقًا وسماه ثيودور بوفيري في 1888.[3][4]
الوظائف
يرتبط الجسيم المركزي بالغشاء النووي خلال المرحلة التمهيدية من الدورة الخلوية. خلال الانقسام، يتحطم الغشاء النووي، وتستطيع حينها الأنبيبات حول الجسيم المركزي التفاعل مع الصبغيات لتشكيل مغزل الانقسام.
للمُريكز الأقدم بين المريكزين (المريكز الأصل) دور محوري في صناعة الأهداب والسياط.[5]
يُنسخ الجسيم المركزي مرة واحدة في كل دورة خلوية، فترث كل خلية بنت جسيم مركزي واحد، يحوي بنيتين تدعيان مُرَيكزين. يتضاعف الجسيم المركزي خلال الطور إس من الدورة الخلوية. خلال الطور التمهيدي في عملية الانقسام الخلوي، يهاجر الجسيمان المركزيان إلى القطبين الخلويين المتقابلين من الخلية. يتشكل بعد ذلك مغزل الانقسام بين المريكزين. عند الانقسام، تأخذ كل خلية بنت جسيمًا مركزيًا واحدًا.[6] يرتبط وجود عدد شاذ من الجسيمات المركزية في الخلية بحدوث السرطانات. ويشبه تضاعف الجسيم المركزي تضاعف الحمض النووي في جانبين: كلا التضاعفين من النمط نصف المحافظ ويلعب سي دي كي 2 دورًا منظمًا للعملية. لكن العمليتين مختلفتان بشكل أساسي بأن تضاعف الجسيم المركزي لا يحدث من خلال قراءة قوالب وتجميعها. تساعد المريكزات الأم في تجميع المواد اللازمة لتكوين المريكزات البنات.[7]
لكن المريكزات غير ضرورية لتقدم الانقسام الخلوي. عندما تُشعَع المريكزات باستخدام الليزر، يستمر الانقسام عادة بشكل طبيعي مع تشكل مغزل طبيعي الشكل. تتطور ذبابة الفاكهة (دورسوفيلا) بشكل طبيعي عندما يغيب المريكز بسبب طفرة في الجين اللازم لتضاعف المريكزات. عند غياب المريكزات، تُجمع الأنبيبات الدقيقة من خلال المحركات الجزيئية، وهو ما يسمح بتشكيل المغزل ثنائي القطب. تستطيع الكثير من الخلايا إكمال الطور البيني دون وجود المريكزات. [5][8]
الجسيم المركزي ضروري لبقاء الكائن الحي. تفتقر الخلايا التي لا تحتوي على الجسيم المركزي التجمعات الشعاعية للأنبيبات الدقيقة النجمية. ويكون موضع المغزل فيها مَعيبًا ولا يتحقق التوضع المركزي للعضيات عند الانقسام الهيولي. وظيفة الجسيم المركزي المفترضة في هذا السياق التحقق من صلاحية الانقسام الخلوي، لأنه يزيد فعالية الانقسام بشكل كبير. تتوقف بعض الأنواع الخلوية في الانقسام الخلوي التالي عندما تغيب الجسيمات المركزية. وليست هذه الحالة ظاهرة شاملة لكل الخلايا.
عند تخصيب بيضة دودة الربداء الرشيقة، تقدم الحيوانات المنوية زوجًا من المريكزات. ستشكل هذه المريكزات الجسيمات المركزية، التي ستوجه أول انقسام خلوي للبيضة الملقحة، وهو ما سيحدد قطبيتها. ليس واضحًا بعد ما إذا كان دور الجسيم المركزي في تحديد القطبية معتمدًا على الأنيبيبات الدقيقة أو أنه مستقل عنها. [9]
في التكاثر البشري، تقدم الحيوانات المنوية المريكزات التي تشكل الجسيم المركزي وجهاز الأنيبيبات الدقيقة في البيضة الملقحة.
التبدلات في الجسيم المركزي في الخلايا السرطانية
وصف ثيودور بوفيري في 1914 اضطرابات الجسيمات المركزية في الخلايا السرطانية. توسع نطاق هذه الملاحظة الأولية فشمل الكثير من الأورام البشرية.[10] يمكن توزيع اضطرابات الجسيمات المركزية في مجموعتين فرعيتين: الشذوذات البنيوية والشذوذات العددية، ويمكن العثور على النوعين في وقت واحد في الخلية الورمية.
الشذوذات البنيوية
تظهر التغيرات البنيوية عادة نتيجة التعبير عن مكونات الجسيم المركزي على نحو غير مضبوط، أو بسبب تعديلات تالية للترجمة (مثل الفسفرة) غير ملائمة للمكونات. يمكن أن تسبب هذه التعديلات تغيرات في حجم الجسيم المركزي (يكون عادة كبيرًا جدًا بسبب زيادة المادة حول المريكزات). وبالإضافة إلى ذلك، ولأن بروتينات الجسيم المركزي تميل للتجمع، تُشاهد أجسام مرتبطة بالجسيم المركزي في أماكن أخرى. يشبه الجسيمان المركزيان المتضخمان والأجسام المرتبطة بالجسيم المركزي بنى من الجسيمات المركزية موجودة في الخلايا الورمية. يمكن أن يُحرَض تكون هذه البنى في الزرع الخلوي من خلال فرط التعبير عن بروتينات محددة في الجسيم المركزي، مثل سي إن إيه بّي 1 وإن آي بّي. يمكن أن تبدو هذه البنى متشابهة جدًا لكن الدراسات المفصلة كشفت أنها تملك خصائص مختلفة جدًا، تبعًا للتكوين البروتيني. فمثلًا، تختلف قدرتها على ضم معقدات غاما تي يو آر سي، وبذلك تؤثر قدرتها على تنوية الأنيبيبات الدقيقة على شكل وقطبية وحركية خلايا الورم الناتج بطرق مختلفة.[11][12]
الشذوذات العددية
عندما توجد الجسيمات المركزية في الخلية بعدد غير كافي يظهر عدم استقرار الجينوم الخلوي وفقدان التمايز النسيجي. ولكن طريقة عد الجسيمات المركزية (بالاعتماد على أن كل جسيم مركزي يضم مريكزين) طريقة غير دقيقة غالبًا، إذ يستخدم المجهر الفلوري لتقييمها وهذا المجهر لا يتمتع بدقة بصرية قادرة على كشف المريكزات المتقاربة جدًا. من الواضح أن وجود فائض من الجسيمات المركزية أمر شائع في الأورام البشرية. لوحظ أن فقدان الجين الكابح للورم بي 53 يسبب زيادة في عدد الجسيمات المركزية، بالإضافة إلى الإخلال بتنظيم بعض البروتينات المتورطة في تشكل الأورام البشرية، مثل بي آر سي إيه 1 و2. يمكن أن تحدث زيادة في عدد الجسيمات المركزية بآليات مختلفة: مضاعفة الجسيم المركزي، فشل في الانقسام الهيولي خلال الانقسام الخلوي (وينتج عنه زيادة في عدد الصبغيات)، الالتحام الخلوي (كما في حالات العدوى بفيروسات محددة)، التشكل الجديد للجسيم المركزي. لا توجد حاليًا معلومات كافية لمعرفة مدى انتشار هذه الآليات في خلايا الكائن الحي، ويُعتقد أن زيادة عدد الجسيمات المركزية بسبب الأخطاء خلال الانقسام الخلوي أكثر شيوعًا مما نعرف. ولأن الكثير من العيوب الأولية في خلية واحدة (خلل تنظيم الدورة الخلوية، الدنا المَعيب أو الخلل في استقلاب الكروماتين، الفشل في نقاط التحقق في المغزل وغيرها) ستسبب فشلًا في انقسام الخلية، وزيادة في الصيغة الصبغية وزيادة عدد الجسيمات المركزية بوصفه أثرًا «ثانويًا».[13][14]
التطور
دُرس التاريخ التطوري للجسيم المركزي والمريكز من خلال بعض الجينات المميزة مثل السينترينات. تشارك السينترينات في إطلاق إشارة الكالسيوم وتلزم في تضاعف المريكزات. توجد عائلتان فرعيتان رئيسيتان من هذه الجينات توجدان عند الجيارديا المعوية، وهي أحد الكائنات حقيقية النوى مبكرة التفرع. ما يعني أن السينترينات كانت موجودة في الخلية حقيقية النوى المشتركة السلف. وعلى النقيض، لا يوجه تركيب متناد قابل للكشف عند العتائق أو الجراثيم وبذلك تكون السينترينات جزءًا من الجينات المميزة لحقيقيات النوى. على الرغم من وجود دراسات على تطور السينترينات والمريكزات، لم تنشر أي دراسة عن تطور المادة حول المريكزات.[15]
من الواضح أن بعض أجزاء الجسيم المركزي مختلفة للغاية عند ذبابة الفاكهة الشائعة وديدان الربداء الرشيقة. فمثلًا، يفتقد النوعان إحدى العائلات الفرعية للسينترينات المتعلقة بتضاعف المريكز. يمكن أن تتطور الأفراد الطافرة من ذبابة الفاكهة الشائعة التي تفتقد الجسيم المركزي إلى ذبابات بالغة طبيعية الشكل، لكنها تموت سريعًا بعد ذلك، لأن خلاياها العصبية الحسية غير مزودة بالأهداب. وبالتالي، طورت هذه الذبابة آلية إضافية مستقلة عن الجسيمات المركزية.
النكيلوتيدات المرتبطة بالجسيم المركزي
أشارت الأبحاث في 2006 إلى أن الجسيمات المركزية في بيوض بطلينوس الأمواج الأطلسي تحوي تسلسلات من الحمض النووي رنا. تبين أن هذه التسلسلات غير موجودة تقريبًا في أي جزء آخر من الخلية، ولا توجد في قاعدة بيانات الجينوم. يحتوي أحد تسلسلات الرنا التي جرى التعرف عليها بوليميراز رنا محتمل، وهو ما دفع إلى افتراض وجود جينوم من الرنا في الجسيم المركزي. بينت الأبحاث اللاحقة أن الجسيم المركزي لا يحتوي على جينوم من الدنا. ورغم التأكد من وجود جزيئات الرنا في الجسيم المركزي، تبين أن التسلسلات نفسها موجودة في نواة الخلية. يمكن أن تتشكل جسيمات مركزية جديدة بعد إزالة الجسيمات المركزية (مثلما يحدث عند إزالتها بالتشعيع الليزري).[16]
المراجع
- "معلومات عن جسيم مركزي على موقع wikiskripta.eu"، wikiskripta.eu، مؤرشف من الأصل في 3 نوفمبر 2020.
- "معلومات عن جسيم مركزي على موقع amigo.geneontology.org"، amigo.geneontology.org، مؤرشف من الأصل في 5 أكتوبر 2018.
- Boveri, Theodor (1888)، Zellen-Studien II: Die Befruchtung und Teilung des Eies von Ascaris megalocephala.، Jena: Gustav Fischer Verlag، مؤرشف من الأصل في 26 أغسطس 2021.
- Bloodgood, RA (2009)، "From central to rudimentary to primary: the history of an underappreciated organelle whose time has come. The primary cilium."، Methods in Cell Biology، 94: 3–52، doi:10.1016/S0091-679X(08)94001-2، ISBN 9780123750242، PMID 20362083.
- Rieder, C. L.؛ Faruki, S.؛ Khodjakov, A. (أكتوبر 2001)، "The centrosome in vertebrates: more than a microtubule-organizing center"، Trends in Cell Biology، 11 (10): 413–419، doi:10.1016/S0962-8924(01)02085-2، ISSN 0962-8924، PMID 11567874.
- Stearns, T. (مايو 2001)، "Centrosome duplication. A centriolar pas de deux"، Cell، 105 (4): 417–420، doi:10.1016/S0092-8674(01)00366-X، ISSN 0092-8674، PMID 11371338، S2CID 1622118.
- Rodrigues-martins, A.؛ Riparbelli, M.؛ Callaini, G.؛ Glover, D. M.؛ Bettencourt-dias, M. (2007)، "Revisiting the Role of the Mother Centriole in Centriole Biogenesis"، Science، 316 (5827): 1046–50، Bibcode:2007Sci...316.1046R، doi:10.1126/science.1142950، hdl:10400.7/955، PMID 17463247، S2CID 6965044.
- Basto, R.؛ Lau, J.؛ Vinogradova, T.؛ Gardiol, A.؛ Woods, G.؛ Khodjakov, A.؛ Raff, W. (يونيو 2006)، "Flies without centrioles"، Cell، 125 (7): 1375–1386، doi:10.1016/j.cell.2006.05.025، ISSN 0092-8674، PMID 16814722، S2CID 2080684.
- Hewitson, Laura & Schatten, Gerald P. (2003)، "The biology of fertilization in humans"، في Patrizio, Pasquale؛ وآخرون (المحررون)، A color atlas for human assisted reproduction: laboratory and clinical insights، Lippincott Williams & Wilkins، ص. 3، ISBN 978-0-7817-3769-2، اطلع عليه بتاريخ 09 نوفمبر 2013.
- Nigg, E.A. (2002)، "Centrosome aberrations: cause or consequence of cancer progression? and taking little boys and girls away"، Nat Rev Cancer، 2 (11): 815–821، doi:10.1038/nrc924، PMID 12415252، S2CID 9038803.
- Lingle, W.L.؛ Barrett, S.L.؛ Negron, V.C.؛ D'assoro, A.B.؛ Boeneman, K.؛ Liu, W.؛ Whitehead, C.M.؛ Reynolds, C.؛ Salisbury, J.L. (2002)، "Centrosome amplification drives chromosomal instability in breast tumor development"، Proc Natl Acad Sci USA، 99 (4): 1978–1983، Bibcode:2002PNAS...99.1978L، doi:10.1073/pnas.032479999، PMC 122305، PMID 11830638.
- Fry, A.M.؛ Mayor, T.؛ Meraldi, P.؛ Stierhof, Y.D.؛ Tanaka, K.؛ Nigg, E.A. (1998)، "C-Nap1, a Novel Centrosomal Coiled-Coil Protein and Candidate Substrate of the Cell Cycle–regulated Protein Kinase Nek2"، J Cell Biol، 141 (7): 1563–1574، doi:10.1083/jcb.141.7.1563، PMC 2133000، PMID 9647649.
- Meraldi, P.؛ Honda, R.؛ Nigg, E.A. (2002)، "Aurora-A overexpression reveals tetraploidization as a major route to centrosome amplification in p53–/– cells"، EMBO J، 21 (4): 483–492، doi:10.1093/emboj/21.4.483، PMC 125866، PMID 11847097.
- Storchova, Z.؛ Pellman, D. (2004)، "From polyploidy to aneuploidy, genome instability and cancer"، Nat Rev Mol Cell Biol، 5 (1): 45–54، doi:10.1038/nrm1276، PMID 14708009، S2CID 11985415.
- Marshall, W. F. (2009)، "Centriole evolution"، Current Opinion in Cell Biology، 21 (1): 14–15، doi:10.1016/j.ceb.2009.01.008، PMC 2835302، PMID 19196504.
- Alliegro, M. C.؛ Alliegro, M. A.؛ Palazzo, R. E. (2006)، "Centrosome-associated RNA in surf clam oocytes"، Proceedings of the National Academy of Sciences، 103 (24): 9034–9038، Bibcode:2006PNAS..103.9034A، doi:10.1073/pnas.0602859103، PMC 1482561، PMID 16754862.
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة علم الأحياء
بوابة علم الأحياء