فينكولين
يوجد الفينكولين في خلايا الثدييّات وهو بروتين هيكلي خلوي يتوضّع في لويحات الالتصاق البؤريّة الّتي تكتنِف ارتباط جزيئات الالتصاق الإنتغرين بأكتين الهيكل الخلوي. تمّ اكتشافه بشكل مستقل بواسطة Benny Geiger [1] و كيث بوريدج,[2] سلسلته مشابهة بنسبة 20%-30% للكاتينين-α, الذي يقوم بذات الوظيفة. يتغيّر شكل بروتين الفينكولين عند ارتباطه بالتناوب مع التالين أو الأكتينين-α, و كنتيجةٍ تتغيّر خواصه الارتباطيّة. تحدث مورثة الفينكولين كنسخة وحيدة ويبدو أنّها لا تقوم بوظائف أكثر في حال غيابها. يحتاج أيضاً متخالفه الوصلي الميتافينكولين (انظر في الأسفل) إلى الفينكولين ليكون مثنويّاً مُتخالِفاً ويعمل بشكلٍ مستقلّ. الفينكولين بروتين هيكلي خلوي يُرافِق المَواصِل خلية-خلية وخلية-نسيج خلالي، حيث يُعتقَد أنّ وظيفتَه كأحد البروتينات المتفاعلة المُكتنفة في تثبيت F-أكتين بالغشاء الخلوي. تمّ اكتشاف نسخ وصل متعاقبة متعدّدة لهذه المورثة, لكن لم يتم تحديد الثبوت البيولوجي لبعض الأشكال. يُبدي بروتين الفينكولين البشري تطابق تسلسلي بنسبة أكبر من 95% لبروتين الفينكولين لدى الدجاج.[3] }}
| Vinculin | |||||||
|---|---|---|---|---|---|---|---|
| المعرفات | |||||||
| الأسماء المستعارة | epididymis secretory sperm binding protein, epididymis luminal protein 114, vinculin, metavinculin, VCL, meta-vinculin, MV | ||||||
| معرفات خارجية | |||||||
| نمط التعبير عن الحمض النووي الريبوزي | |||||||
 | |||||||
| المزيد من بيانات التعبير المرجعية | |||||||
| أورثولوج | |||||||
| الأنواع | الإنسان | الفأر | |||||
| أنتريه | n/a | ||||||
| Ensembl | n/a | n/a | |||||
| يونيبروت |
|
| |||||
| RefSeq (مرسال ر.ن.ا.) |
|
| |||||
| RefSeq (بروتين) |
|
| |||||
| الموقع (UCSC | n/a | ||||||
| بحث ببمد | n/a | ||||||
| ويكي بيانات | |||||||
| |||||||
البنية
الفينكولين بروتين هيكلي خلوي بوزن 117-كيلو دالتون، يتكوّن من 1066 حمض أميني. يحتوي البروتين على منطقة مِطرافيّة نتروجينيّة حمضيّة ومنطقة مِطرافيّة كربونيّة قاعديّة، يفصل بينهُما قطعة متوسِّطة غنيّة بالبرولين. يتألّف الفينكولين من منطقة رأسيّة كرويّة تحتوي على مواقع الارتباط بالتالين و ال α-أكتينين بالإضافة لموقع فَسفتَة التيروزين, بينما تحتوي منطقة الذّيل على مواقع ارتباط ل F-أكتين والباكسيلين و اللبيدات (غولدمان . 2001). بشكل أساسي، يوجد رأس مِطرافي نتروجيني يحتوي على 835 حمض أميني, مُنقسِم إلى أربعة أجزاء. يُربَط بالرأس المِطرافي الكربوني بواسطة منطقة رابطة.
التعديل
يُسلِّط الاكتشاف الحديث للبنية ثلاثية الأبعاد الضوء على الكيفية الّتي يعدِّل بها هذا البروتين شكله ليُنجِز وظائف متنوّعة. مثلاً, الفينكولين قادر على تنظيم حركة الخلية عبر تعديل بسيط في شكله من فعّال إلى غير فعّال. عندما يكون بحالته غير الفعّالة، يتم وصف تعديل الفينكولين بأنّه تفاعل بين منطقتي الرأس والذيل. و عندما يتحوّل إلى الشكل الفعّال، كما عندما يثير التالين الارتباط، يتم فصل التفاعل الجزيئي الداخلي بين الرأس والذيل. بكلمات أخرى، عندما ترتبط مواقع ربط التالين في الحلزونات-α بالحزمة الحلزونية في منطقة الرأس من الفينكولين، يتم بدء عمليّة ’تحويل الحزمة الحلزونية‘, التي تؤدّي إلى إعادة تنظيم الحلزونات-α (α1-α-4), منتجةً خمس حزم حلزونية جديدة كليّاً. تمتد هذه الوظيفة أيضاً إلى الخلايا السرطانيّة وتنظيم حركتها وتكاثر السرطان إلى أجزاء أخرى من الجسم.
الآلية والوظيفة
يحدث انتشار وحركة الخلية بالرغم من عملية ارتباط مستقبلات الإنتغرين على سطح الخلية مع جزيئات الالتصاق في المادة الخلاليّة. يتم ربط الفينكولين بالالتصاق البؤري والمُواصِل المُلتصِقة، وهي معقدّات مُنوّاة بخيوط الأكتين وروابط متصالبة بين الوسط الخارجي و الغشاء البلازمي و هيكل الخلية الأكتيني[4] (Xu. 1998). يتألّف المُعقد في الالتصاقات البؤريّة من بروتينات عديدة مثل الفينكولين و α-أكتين والباكسيلين والتالين، على السطح الخلوي الداخلي للغشاء البلازمي. بوصف أكثر نوعيّة، يرتبط المِطراف الأميني للفينكولين بالتالين، الذي بدوره يرتبط بالإنتغرين-β و يرتبط المِطراف الكربوكسيلي بالأكتين والشحوم الفوسفوريّة و المثنويّات المُتجانِسة المُكوِّنة للباكسيلين. يتم تنظيم ارتباط الفينكولين بالتالين والأكتين بواسطة عديدات الفوسفوإينوزيتيد ويُثبّط بواسطة الشحوم الفوسفوريّة الحمضيّة. ثُمّ يقوم المعقد بتثبيت خيوط الأكتين بالغشاء[5] (Ezzell. 1997). يؤثّر فقدان بروتين الفينكولين على وظائف خلوية متنوعة؛ فيعطّل تشكيل المعقد، ويمنع التصاق الخلية وانتشارها. يتظاهر غياب البروتين بنقص في انتشار الخلايا، مصحوب بانخفاض في تشكُّل ألياف التوتر وتشكُّل التصاقات بؤريّة قليلة وتثبيط امتداد الأرجل الكاذبة الصُفاحِيّة[6] (غولدمان . 2001). تم اكتشاف أنّ الخلايا مَعوزَة الفينكولين تمتلك مخاريط نمو متقدمة ببطء أكثر، بالإضافة إلى أرجل كاذبة خيطيّة وأرجل صُفاحِيّة أقل ثباتاً من النمط المُفرِط. بالاعتماد على البحث، تمّ افتراض أنّ عوز الفينكولين ربما ينقص التصاق الخلية عبر تثبيط تجمّع الالتصاق البؤري ومنع بلمرة الأكتين. من ناحية أخرى، ربما يعيد تعبير الفينكولين المُفرِط الالتصاق والانتشار عبر تعزيز إجلاب البروتينات الهيكلية الخلوية إلى معقد الالتصاق البؤري في موقع ارتباط الإنتغرين[5](Ezzell .1997). يبدو أنّ مقدرة الفينكولين على التفاعل مع الإنتغرين على الهيكل الخلوي في الالتصاق البؤري تكون حديّة من أجل التحكم بالميكانيكا الهيكلية الخلوية وانتشار الخلية وتشكل الأرجل الكاذبة الصُفاحيّة. إذاً, يبدو أنّ الفينكولين يلعب دور المفتاح في التحكّم الشكلي المُعتمِد على قدرته في تعديل وظيفة وبنية الالتصاق البؤري.
موقع ارتباط الفينكولين
| VBS | |
|---|---|
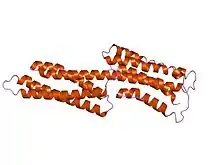 | |
| رأس الفينكولين البشري (1-258) في المعقد مع الموقع الثالث لارتباط الفينكولين على التالين (ثمالات 1944-1969) | |
| معرف | |
| رمز | VBS |
| قاعدة بيانات عوائل البروتينات | PF08913 |
| إنتربرو | IPR015009 |
توجد مواقع ارتباط الفينكولين بشكل سائد في بروتين التالين والجزيئات الشبيهة به، مُمَكِّنَةً الفينكولين من الارتباط بالتالين، ومثبِّتةً المَواصِل النسيجيّة الخلويّة المتواسطة بالإنتغرين. ثُم يَربِط التالين الإنتغرين بأكتين الهيكل الخلوي. التسلسل التوافقي لمواقع ارتباط الفينكولين هو LxxAAxxVAxxVxxLIxxA, مع التكهُّن بأربع حلزونات متقابلة الزُّمر كبنية ثانويّة. تحجُب الثمالات الكارهة للماء التي تُظهِر مواقع ارتباط الفينكولين نفسها وتُدفن في قلب سلسلة من الحزم الحلزونية التي تُكوِّن قضيب التالين.[7]
مُتخالف الوصل: ميتافينكولين
تُعبِّر العضلات الملساء والعضلات الهيكلية ( و ربما بمدى منخفض في العضلة القلبية) في حالة تمايزها الخلوي الجيد (قلوص) معاً (برفقة الفينكولين) المُتخالِف الوصلي حيث تحمل إكزون فائض في منطقة الترميز 3, بالتالي ترميز شكل مُماثل شكلي أطول ميتافينكولين بوزن جزيئي ~150 كيلو دالتون - و هو بروتين تم اكتشاف وجوده في ثمانينات القرن العشرين.[8] تُؤدّي ترجمة القطعة المُرمَّزة (exon) الفائضة إلى زيادةٍ في الحموض الأمينية بمقدار 68 إلى 79 حمض أميني يتم حجبها بين الحلزونين I و II في منطقة الذيل المطرافي الكربوني. يرتبط اعتلال العضلة القلبية المتوسع الوراثي مجهول السبّب بالطفرات الحادثة في المنطقة المحجوبة.[9] يُقدَّر طول المنطقة المحجوبة في الميتافينكولين ب 68 حمض أميني عند الثدييّات و 79 حمض أميني عند الضفادع. Strasser.[10] تم مقارنة سلاسل الميتافينكولين من الخنزير والرجل والدجاج والضفدع، وتم اكتشاف أنّ المنطقة المحجوبة ذات قسمين: القسم الأوّل مُتغيّر والثاني محفوظ بشدّة. تتوضع كل المماثلات الشكليّة للفينكولين في بُنى الالتصاق العضليّة، مثل اللويحات الكثيفة في العضلات الملساء والأقراص المُقحمة في الخلايا العضليّة القلبيّة وبروتينات كوستامر (costamere)في العضلات الهيكلية.[11] تمتلك منطقة ذيل الميتافينكولين ألفة منخفضة لمنطقة الرأس بالمقارنة مع ذيل الفينكولين. في حالة الميتافينكولين، يتجلى إضعاف عروة الدبوس الكارهة للماء للمطراف الكربوني في منطقة الذيل بواسطة الشحنات السالبة للحموض الأمينية المحجوبة (68 حمض أميني), بالتالي يتطلّب التفعيل الكامل لجزيء الميتافينكولين وجود المماثل الشكلي المُنتظم المُفعَّل بالشحوم الفوسفورية من الفينكولين.
التداخلات
يبدي الفينكولين تداخلاً مع البروتين المحتوي على منطقة SH3 و الصوربين (SORBS1),[12] و بروتين الكادرين 1 (CDH1) [13][14] و بروتين الباكسيلين.[15][16][17]
المراجع
- Geiger,B. 130K Protein from Chicken Gizzard - Its Localization at the Termini of Microfilament Bundles in Cultured Chicken-Cells. Cell 18, 193-205 (1979).
- Burridge,K. & Feramisco,J.R. Microinjection and localization of a 130K protein in living fibroblasts: a relationship to actin and fibronectin. Cell 19, 587-595 (1980).
- "Entrez Gene: VCL vinculin"، مؤرشف من الأصل في 05 ديسمبر 2010.
- Xu؛ Baribault؛ Adamson (1998)، "vinculin knockout results in heart and brain defects during embryonic development"، Development، 125 (2): 327–337، PMID 9486805.
- Ezzell RM, Goldmann WH, Wang N, Parasharama N, Ingber DE. (1997). Vinclin promotes cell spreading by mechanically coupling integrins to the cytoskeleton. Experimental Cell Research. 231(1):14-26.
- Goldmann؛ Ingber (2001)، "Intact vinculin protein is Require for control of cell shape, cell mechanics, and rac-Dependent Lamellipodia Formation"، Biochemical and Biophysical Research Communications، 290 (2): 749–755، doi:10.1006/bbrc.2001.6243، PMID 11785963.
- Gingras AR, Vogel KP, Steinhoff HJ, Ziegler WH, Patel B, Emsley J, Critchley DR, Roberts GC, Barsukov IL (فبراير 2006)، "Structural and dynamic characterization of a vinculin binding site in the talin rod"، Biochemistry، 45 (6): 1805–17، doi:10.1021/bi052136l، PMID 16460027.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - JR Feramisco, JE Smart, K Burridge, DM Helfman, and GP Thomas Co-existence of vinculin and a vinculin-like protein of higher molecular weight in smooth muscle J. Biol. Chem. 257: 11024-11031.
- Sebastian Witt, Anke Zieseniss, Ulrike Fock, Brigitte M. Jockusch, and Susanne Illenberger Comparative Biochemical Analysis Suggests That Vinculin and Metavinculin Cooperate in Muscular Adhesion Sites J. Biol. Chem. 279: 31533-31543.
- Strasser, P.; Gimona, M.; Herzog, M.; Geiger, B.; Small, J. V. : Variable and constant regions in the C-terminus of vinculin and metavinculin: cloning and expression of fragments in E. coli. FEBS Lett. 317: 189-194, 1993. PubMed ID : 8425604
- Belkin, A. M., Ornatsky, O. I., Glukhova, M. A., and Koteliansky, V. E. (1988) J. Cell Biol. 107, 545–553
- Mandai, K (مارس 1999)، "Ponsin/SH3P12: an l-afadin- and vinculin-binding protein localized at cell-cell and cell-matrix adherens junctions"، J. Cell Biol.، UNITED STATES، 144 (5): 1001–17، doi:10.1083/jcb.144.5.1001، ISSN 0021-9525، PMC 2148189، PMID 10085297.
{{استشهاد بدورية محكمة}}: الاستشهاد يستخدم وسيط مهمل|lay-url=(مساعدة)، الوسيط author-name-list parameters تكرر أكثر من مرة (مساعدة) - Hazan, R B (ديسمبر 1997)، "Vinculin is associated with the E-cadherin adhesion complex"، J. Biol. Chem.، UNITED STATES، 272 (51): 32448–53، doi:10.1074/jbc.272.51.32448، ISSN 0021-9258، PMID 9405455.
{{استشهاد بدورية محكمة}}: الاستشهاد يستخدم وسيط مهمل|lay-source=(مساعدة)، الوسيط author-name-list parameters تكرر أكثر من مرة (مساعدة) - Hazan, R B (أبريل 1998)، "The epidermal growth factor receptor modulates the interaction of E-cadherin with the actin cytoskeleton"، J. Biol. Chem.، UNITED STATES، 273 (15): 9078–84، doi:10.1074/jbc.273.15.9078، ISSN 0021-9258، PMID 9535896.
{{استشهاد بدورية محكمة}}: الاستشهاد يستخدم وسيط مهمل|lay-source=(مساعدة)، الوسيط author-name-list parameters تكرر أكثر من مرة (مساعدة) - Turner, C E (1999)، "Paxillin LD4 motif binds PAK and PIX through a novel 95-kD ankyrin repeat, ARF-GAP protein: A role in cytoskeletal remodeling"، J. Cell Biol.، UNITED STATES، 145 (4): 851–63، doi:10.1083/jcb.145.4.851، ISSN 0021-9525، PMC 2133183، PMID 10330411.
{{استشهاد بدورية محكمة}}: الاستشهاد يستخدم وسيط مهمل|lay-source=(مساعدة)، الوسيط author-name-list parameters تكرر أكثر من مرة (مساعدة)، الوسيط غير المعروف|شهر=تم تجاهله (مساعدة) - Mazaki, Y (مارس 1997)، "Monocyte cells and cancer cells express novel paxillin isoforms with different binding properties to focal adhesion proteins"، J. Biol. Chem.، UNITED STATES، 272 (11): 7437–44، doi:10.1074/jbc.272.11.7437، ISSN 0021-9258، PMID 9054445.
{{استشهاد بدورية محكمة}}: الاستشهاد يستخدم وسيط مهمل|lay-source=(مساعدة)، الوسيط author-name-list parameters تكرر أكثر من مرة (مساعدة) - Brown, M C (نوفمبر 1996)، "Identification of LIM3 as the principal determinant of paxillin focal adhesion localization and characterization of a novel motif on paxillin directing vinculin and focal adhesion kinase binding"، J. Cell Biol.، UNITED STATES، 135 (4): 1109–23، doi:10.1083/jcb.135.4.1109، ISSN 0021-9525، PMC 2133378، PMID 8922390.
{{استشهاد بدورية محكمة}}: الاستشهاد يستخدم وسيط مهمل|lay-url=(مساعدة)، الوسيط author-name-list parameters تكرر أكثر من مرة (مساعدة)
قراءة إضافيّة
{{PBB_Further_reading | citations =
- Critchley DR (2005)، "Cytoskeletal proteins talin and vinculin in integrin-mediated adhesion"، Biochem. Soc. Trans.، 32 (Pt 5): 831–6، doi:10.1042/BST0320831، PMID 15494027.
- Koteliansky VE, Ogryzko EP, Zhidkova NI؛ وآخرون (1992)، "An additional exon in the human vinculin gene specifically encodes meta-vinculin-specific difference peptide. Cross-species comparison reveals variable and conserved motifs in the meta-vinculin insert"، Eur. J. Biochem.، 204 (2): 767–72، doi:10.1111/j.1432-1033.1992.tb16692.x، PMID 1339348.
{{استشهاد بدورية محكمة}}: Explicit use of et al. in:|مؤلف=(مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Mulligan LM, Gardner E, Telenius H, Ponder BA (1992)، "Complementary physical and genetic techniques map the vinculin (VCL) gene on chromosome 10q"، Genomics، 13 (4): 1347–9، doi:10.1016/0888-7543(92)90066-2، PMID 1505973.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Weller PA, Ogryzko EP, Corben EB؛ وآخرون (1990)، "Complete sequence of human vinculin and assignment of the gene to chromosome 10"، Proc. Natl. Acad. Sci. U.S.A.، 87 (15): 5667–71، doi:10.1073/pnas.87.15.5667، PMC 54388، PMID 2116004.
{{استشهاد بدورية محكمة}}: Explicit use of et al. in:|مؤلف=(مساعدة)صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Turner CE, Burridge K (1989)، "Detection of metavinculin in human platelets using a modified talin overlay assay"، Eur. J. Cell Biol.، 49 (1): 202–6، PMID 2503380.
- Turner CE, Miller JT (1994)، "Primary sequence of paxillin contains putative SH2 and SH3 domain binding motifs and multiple LIM domains: identification of a vinculin and pp125Fak-binding region"، J. Cell. Sci.، 107 (6): 1583–91، PMID 7525621.
- {{استشهاد بدورية محكمة | مؤلف=Salgia R,
روابط خارجية
- MBInfo - Vinculin
- http://www.stjude.org/media/0,2561,453_5297_9660,00.html
- http://www.medicalnewstoday.com/medicalnews.php?newsid=5196
- http://www.olympusmicro.com/primer/techniques/fluorescence/gallery/cells/bpae/images/bpaelarge.jpg
- Vinculin Info with links in the Cell Migration Gateway
{kind=link}
قالب:Cytoskeletal Proteins
 بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية