مراقبة الرنا الرسول
مراقبة جودة الرنا الرسول هي آليات تستخدمها الكائنات لضمان جودة ودقة جزيئات الرنا الرسول. توجد عدة آليات مراقبة في الخلية وتعمل هذه الآليات على عدة مراحل من حياة الرنا الرسول ويمكنها اكتشاف وتحديد جزيئات الرنا الرسول التي بها عيب في النسخ أو بها طفرة ثم تفكيكها.
ترجمة جزيئات الرنا الرسول المعيبة يمكن أن تؤدي إلى آثار ضارة على الكائن. تميل البروتينات غير الطبيعية الناتجة عن ترجمة جزيئات الرنا الرسول هذه إلى تكوين تكدساتٍ تؤدي إلى موت الخلية وإلى تكون الرقع النشوانية في الأنسجة.
نظرة عامة
ترجمة نسخ الرنا الرسول إلى بروتينات جزء حياتٌِّي في هدف علم الأحياء الجزيئي الرئيسي. مع ذلك، جزيئات الرنا الرسول عرضة للعديد من الأخطاء في دقة النسخ والتي تسبب بدورها أخطاءً في ترجمة الرنا إلى البروتين المطلوب.[1] آليات مراقبة الرنا هي طرق تستخدمها الخلية لضمان جودة ودقة جزيئات الرنا الرسول.[2] ويتم هذا عموما عبر وسم جزيئات الرنا الرسول المعيبة بطريقة محددة كي تتعرف عليها إنزيمات النوكليازات داخلية المنشأ وتقوم بتفكيكها.[3]
وُثِّقت مراقبة الرنا الرسول لدى البكتيريا والخميرة. لدى حقيقيات النوى، تعمل هذه الآليات في كل من النواة والسيتوبلازم.[4] ينتج عن فحوصات دقة جزيئات الرنا الرسول في النواة تفكيك النسخ المعالجة بشكل غير صحيح قبل تصديرها إلى السيتوبلازم. بعد ذلك تتعرض هذه النسخ إلى المزيد من المراقبة حين تصل السيتوبلازم. تفحص آليات المراقبة السيتوبلازمية نُسَخ الرنا بحثا عن تواجد أو غياب كودونات التوقف المبتسرة.[ملاحظة 1][3][4]
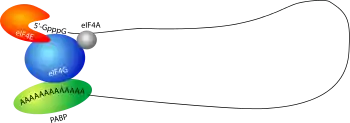
لدى حقيقيات النوى، توظيف الريبوسوم عند بداية الترجمة هو أول مستوى مراقبة جودة الرنا الرسول في السيتوبلازم. في سبيل توظيف الريبوسوم، من الضروري أن يكون الرنا الرسول على هيئة شبه دائرة ويكون ذيل عديد الأدينين 3' مرتبط بـPABP، وتكون القبعة 5' مرتبطة بعامل بدء الترجمة eIF4E، ثم يتآثر هاذين البروتينين مع عامل الترجمة eIF4G.[5] هذه الآلية تضمن -قبل بدء الترجمة- أن الرنا الرسول سليم وكامل ويملك قبعة 5' وذيل عديد الأدينين 3'، ومنه جزيئات الرنا التي لا تحتوي على هاتين البنيتين لا تتم ترجمتها.
توجد ثلاث طرق مراقبة معروفة حاليا تعمل بها الخلايا وهي: تفكك الرنا الرسول المتوسَّط بلا معنى (NMD)، تفكك الرنا الرسول المتوسَّط بلا توقف (NSD)، وتفكك الرنا الرسولال المتوسَّط بلا مُضِي (NGD).
تفكك الرنا الرسول المتوسَّط بلا معنى
نظرة عامة
التفكك المتوسَّط بلا معنى (بالإنجليزية: Nonsense-mediated decay) (NMD) هي عملية تحديد وتفكيك نسخ الرنا الرسول التي تحتوي على كودونات إنهاء مبْتسرة. يمكن أن تنشأ كودونات التوقف المبتسرة عبر عدة آليات: طفرات في دنا الخط الجنسي، طفرات جسدية، أخطاء في النسخ أو أخطاء في معالجة الرنا الرسول بعد النسخ.[6][7] الفشل في التعرف على هذه النسخ وتفكيكها يؤدي إلى إنتاج بروتينات مبتورة يمكن أن تكون ضارة للكائن. بإحداث تفكك النهاية الكربوكسيلية لعديدات الببتيد المبتورة، يمكن لآلية NMD حماية الخلايا ضد تأثيرات الطفرات المكسِبة للوظيفة والطفرات الحاذفة السائدة السلبية.[8] لكودونات التوقف المبتسرة دور في حوالي 30% من الأمراض الوراثية وعليه، تلعب آلية NMD دورا حياتيا في ضمان بقاء الكائن على قيد الحياة وفي صحته ولياقته.[9][10]
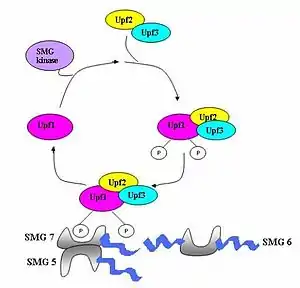
يُجمَّع مركب المراقبة المتكون من مختلف البروتينات (eRF1، eRF3، Upf1، Upf2 ،Upf3) ويقوم بفحص ومسح الرنا الرسول بحثا عن كودونات توقف مبتسرة.[6] يُستثار تجميع هذا المركب بواسطة إنهاء ترجمة مبْتسر. إذا تم تحديد كودون توقف مبْتسر، يتم تأشير نسخة الرنا الرسول للتفكيك ويحدث اقتران التحديد مع التفكيك.[3][11][12]
تم تحديد سبع جينات (smg-1 حتى smg-7) وثلاث جينات UPF (UPF-1 حتى UPF-3) في فطريات الخميرة والربداء الرشيقة على أنها عوامل أساسية عاملة على مفروق مساهمةٌ في نشاط التفكك المتوسط بلا معنى.[13][14] جميع هذه الجينات محفوظة في ذبابة الفاكهة الشائعة والعديد من الثدييات وتلعب دورا حاسما في هذا النوع من التفكك. أما في حقيقيات النوى فتوجد ثلاث مركبات محفوظة في هذا التفكك وهي مركبات Upf1/SMG-2 وUpf2/SMG-3 وUpf3/SMG-4.[15] Upf1/SMG-2 هو بروتين فوسفاتي في الكائنات متعددة الخلايا ويُعتقد أنه يساهم في التفكك المتوسط بلا معنى عبر نشاطه في الفسفرة. مع ذلك، تآثرات هذه البروتينات الدقيقة فيما بينها ودورها في هذا النوع من التفكك ما زال محل نقاش.[12][13][15][16][17]
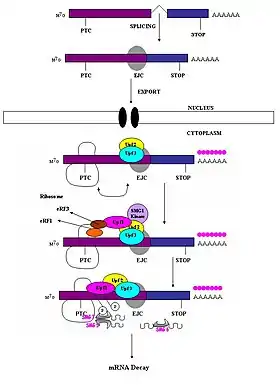
الآلية لدى الثدييات
يجب أن يُميَّز كودون التوقف المبْتسر على أنه مختلف عن كودون التوقف الطبيعي حتى تحدث استجابة لآلية التفكك بلا معنى. لقد لوحظ أن قدرة الكودون المبتسر الذي لا معنى له في إحداث تفكك الرنا الرسول يعتمد على مكانه النسبي بالنسبة لتسلسل عنصر مع المسار والبروتينات المتعلقة به.[1] أثبتت الدراسات أن الكودون المبتسر إذا كان يقع في النوكليوتيدات الأزيد من 50-54 نوكليوتيد عكس المسار (أي باتجاه النهاية 5') لموصل الإكسون-إكسون الأخير فإن بإمكانه استثارة استهداف الرنا الرسول وتفكيكه بهذه الآلية.[1][4][6][7][8][18] أما إن كان مكان الكودون المبتسر يقع في النوكليوتيدات مع المسار من هذه المنطقة فإنه يكون غير قادر على استثارة الآلية. وعليه تقع الكودونات المبتسرة 50-54 نوكليوتيد عكس المسار من آخر موصل لإكسونين في حين تتواجد كودونات التوقف الطبيعية داخل الأكسون الأخير.[19]
تحدِّد مركبات موصل الإكسون (EJCs) مكان موصل الإكسون-إكسون، وهي مركبات عديدة البروتين يتم تجميعها أثناء الوصل في الموضع 20-24 نوكليوتيد عكس المسار من مكان الوصل.[20] مركب موصل الإكسون هو من يوفر المعلومة المطلوبة حول مكان كودون التوقف المبتسر لتمييزه عن كودونات التوقف الطبيعية. يَظهر أن التعرف على الكودونات المبتسرة يعتمد على تحديد مواصل الإكسون-إكسون، وهذا يُشير إلى وجود دور لجسيم الوصل في آلية NMD الخاصة بالثدييات.[18][21] تحرى الباحثون إمكانية وجود دور لجسيم الوصل في آلية NMD الخاصة بالثدييات واكتشفوا أن ذلك ممكن على الأرجح.[19] زيادة على ذلك، لوحِظ أن آلية التفكك المتوسط بلا معنى لا يتم تنشيطها في نُسَخِ الرنا الرسول التي تحتوي كودون مبتسر والتي تم إنتاجها من جينات لا تحتوي على إنترونات طبيعيا ولا تخضع لعملية الوصل (مثل الهستون H4، الشابرون Hsp70، مستقبل ميلانوكورتين 4).[8]
حين يصل الريبوسوم إلى كودن التوقف المبتسر، تتآثر عوامل الترجمة eRF1 وeRF3 مع مركبات موصل الإكسون غير المزالة عبر جسر عديد البروتين.[22] تآثرات UPF1 مع مركب الإنهاء ومع بروتيني UPF2/UPF3 الخاصين بمركبات موصل الإكسون غير المزالة حاسمةٌ وتؤدي إلى وسم الرنا الرسول للتفكيك السريع بواسطة النوكليازات داخلية المنشأ.[19][22]
الآلية لدى اللافقاريات
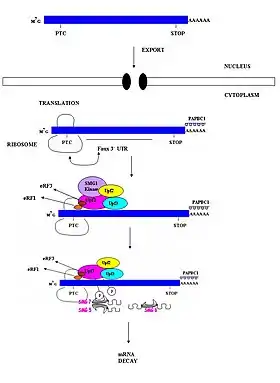
أظهرت الدراسات المتعلقة بكائناتٍ مثل: فطريات الخميرة، ذبابة الفاكهة الشائعة والربداء الرشيقة أن التعرف على كودونات التوقف المبتسرة لدى الكائنات اللافقارية لا يعتمد على مواصل الإكسون-إكسون.[21] واقترحت هذه الدراسات أن NMD لدى اللافقاريات يحدث بشكل مستقل عن الوصل. نتيجة لذلك، مركبات موصل الإكسون المسؤولة على تحديد ووسم مواصل الإكسون-إكسون لا حاجة إليها في NDM لدى اللافقاريات.[3] اقتُرحت العديد من النماذج لتفسير كيفية تمييز كودونات التوقف المبتسرة عن الطبيعية لدى اللافقريات، يقترح أحد هذه النماذج أنه ربما يوجد تسلسل عنصر مع المسار يقوم بوظيفة مماثلة لمركبات وصلة الإكسون لدى الثدييات.[12] نموذج آخر يقترح أن ذيل عديد الأدينين 3' الموجود بشكل واسع في جزيئات الرنا الرسول والمميز لها يمكن أن يوفر معلومةَ الموضعِ المطلوبة للتعرف على الكودون المبتسر.[23] نموذج آخر يسمى «نموذج المنطقة 3' غير المترجمة الخاطئة» يقترح أن إنهاء الترجمة المبتسر يمكن تمييزه عن الإنهاء الطبيعي بسبب الميزات الذاتية الجوهرية لهذه المنطقة التي تسمح لها بالتعرف على وجودها في موضع غير مناسب.[3] هذه النماذج لم يتم إثباتها بشكل حاسم بعد.
الآلية لدى النبات
توجد آليتان للتعرف على كودونات التوقف المبتسرة لدى النبات: تبعا لبعدها عن مركب وصل الإكسون (كما هو الحال لدى الفقاريات) أو بعدها عن ذيل عديد الأدينين. آلية تفكك المتوسط بلا معنى لدى النبات تستحِثُّ تفكيك جزيئات الرنا الرسول التي تحتوي منطقة 3' غير مترجمة أطول من 300 نوكليوتيد، ولهذا نسبة جزيئات الرنا الرسول ذات المناطق 3' غير المترجة الطويلة أخفض بكثير في النباتات منها لدى الفقاريات.[24][25]
تجنب NMD
يُعتقد عموما أن جزيئات الرنا التي تحتوي طفرات لا معنى لها (طفرة هرائية [الإنجليزية]) يتم استهدافها لتفكيكها بواسطة مسارات NMD. يبدو أن تواجد كودونات التوقف المبتسرة حوالي 50-54 نوكليوتيد عكس المسار (باتجاه النهاية 5') من موصل إكسونين يستثير التفكيك السريع. مع ذلك، لوُحظ أن بعض جزيئات الرنا الرسول ذات كودونات توقف مبسترة قادرة على تجنب الاكتشاف والتفكيك.[18][26] بشكل عام، تملك جزيئات الرنا هذه كودون توقف في مكان مبكر جدا من إطار القراءة (بجوار كودون البدء AUG). هذا يبدو تناقضا مع نموذج NMD المقبول حاليا لكون هذا الموضع بعيد نسبيا عن موصل الإكسون-إكسون الأخير عكس المسار.[27]
وُضِّح هذا التجنب في غلوبولين بيتا. جزيئات الرنا الرسول الخاصة بغلوبولين بيتا التي تحتوي طفرة لا معنى لها مبكرا في الإكسون الأول من الجين هي أكثر استقرار من جزيئات الرنا الرسول الحساسة لـNMD. آلية تجنب اكتشافها بالضبط حاليا غير معروفة، واقتُرح أن للبروتين المرتبط بذيل عديد الأدينين (PABP) دور في هذا الاستقرار.[28] وتم في دراسات أخرى إثبات أن تواجد هذا البروتين بالقرب من الكودونات المبتسرة المجاورة لكودون البدء AUG يعزز استقرار جزيئات الرنا هذه التي كانت ستكون حساسة لـNMD من دون تواجده. لوحظ أن هذا التأثير الحامي ليس مقتصرا على معزز غلوبولين بيتا فحسب.[26] مما يقترح أن آلية تجنب NMD هذا يمكن أن تكون منتشرة في أنواع أخرى من الأنسجة لمجموعة متنوعة من الجينات. النموذج الحالي لـNMD ربما يحتاج مراجعة بعد المزيد من الدراسات.
تفكك الرنا الرسول المتوسَّط بلا توقف
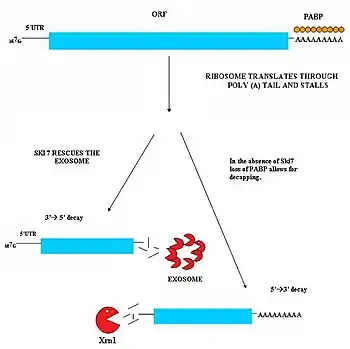
نظرة عامة
التفكك المتوسط بلا توقف (NSD) له دور في تحديد وتفكيك نسخ الرنا الرسول التي لا تملك كودون توقف.[30][31] يمكن أن تنتج نسخ الرنا هذه من عدة آليات مثل التذييل بعديد الأدينيلات السابق لأوانه، إشارات التذييل بعديد الأدينين المشفَّرة داخل المنطقة المشفِّرة من الجين.[32] انعدام كودون التوقف هو مشكل معتبر بالنسبة للخلايا، والريبوسومات المترجِمة للرنا الرسول تستمر في الترجمة وتقوم بترجمة ذيل عديد الأدينين ثم تعلق، وكنتيجة لذلك لا يستطيع الرنا الرسول الانفصال عنها.[33] هذه الريبوسومات تصبح عالقة ومرتبطة بالرنا الرسول الذي لا يملك كودون توقف ولا تكون متاحة لترجمة جزئات رنا أخرى إلى بروتينات. تقوم آلية التفكك المتوسط بلا توقف بحل المشكلة عبر تخليص الريبوسومات العالقة ووسم ذلك الرنا الرسول للتفكيك بواسطة النوكليازات. يوجد مساران لآلية التفكك المتوسط بلا توقف وهذان المساران يعتمدان على وجود أو غياب البروتين Ski7.[30][31]
مسار Ski7
يَنشط هذا المسار حين يكون بروتين Ski7 متوفرا في الخلية. يُعتقد أن البروتين Ski7 يرتبط بالموقع A الفارغ من الريبوسوم، ويسمح له هذا الارتباط بفصل وإخراج الرنا الرسول العالق الذي لا يحتوي كودون توقف مما يحرر الريبوسوم ويسمح له بترجمة نسخ أخرى. بعد الانفصال يكون بروتين Ski7 مرتبطا بالرنا الرسول وهذا الارتباط هو الإشارة التي تجعل الإكسوسوم في العصارة الخلوية يستهدف الرنا الرسول. يقوم مركب Ski7-إكسوسوم بإزالة ذيل عديد الأدينين بسرعة وهذا يسمح للإكسوسوم بتفكيك الرنا الرسول من الاتجاه 3' إلى 5'.[30][31]
مسار غير Ski7
لوحظ نوع ثانٍ من التفكك المتوسط بلا توقف لدى الخميرة. في هذا المسار، ينتج عن غياب بروتين Ski7 فقدانٌ لبروتينات PABP التي ترتبط بذيل عديد الأدينين بسبب الريبوسوم الذي يواصل الترجمة. إزالة بروتينات PABP يتسبب في فقدان القبعة 5'm7G التي تحمي النهاية 5'، وينتج عن هذا الفقدان تفكيك نسخة الرنا بواسطة النوكليازات الخارجية 5'-3' داخلية المنشأ مثل XrnI.[31]
تفكك الرنا الرسول المتوسَّط بلا مضي
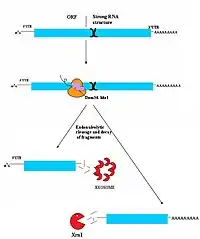
تفكك الرنا الرسول المتوسَّط بلا مضي (NGD) هو أحدث آليات المراقبة اكتشافا،[34] ولهذا فالآلية غير مفهومة جيدا حاليا. مع كون الأهداف الحقيقية للتفكك المتوسَّط بلا مضي مفهومة بشكل ضعيف، يبدو أنها تتكون إلى حد كبير من جزيئات الرنا الرسول التي علقت الريبوسومات بها أثناء ترجمتها. يمكن أن يحدث هذا العلْق والتوقف بسبب عدة عوامل منها بنُى ثانوية قوية، والتي يمكن أن تسد وتمنع ماكينة الترجمة جسديا من التحرك على طول النسخة.[34] يرتبط بروتينا Dom34/Hbs1 على الأرجح بالقرب من الموقع A للريبوسومات العالقة ويساهمان في تخليصها وإتاحتها لترجمات أخرى.[35] في بعض الحالات، تُقص نسخة الرنا كذلك بالقرب من موقع العلق بطريقة النوكليازات الداخلية، لكن هوية النوكلياز الداخلي المسؤول عن ذلك مازالت محل نقاش. تُفكك جزيئات الرنا المقطَّع بشكل كامل بواسطة الإكسوسوم بطريقة 3' إلى 5' وبواسطة Xrn1 بطريقة 5' إلى 3'.[34]
حاليا طريقة تخليص الرنا الرسول من الريبوسومات مازلت غير معروفة، مع ذلك، البروتين Hbs1 ذو صلة قريبة من البروتين Ski7 الذي يلعب دورا واضحا في تخليص وتحرير الريبوسوم في التفكك المتوسط بلا توقف الذي يتم بواسطته. تم افتراض أن Hbs1 ربما يلعب دورا مماثلا في التفكك المتوسط بلا مضي.[6][36]
تطور
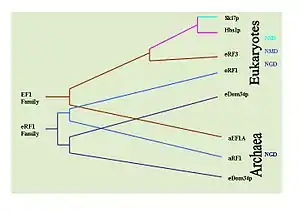
من الممكن تحديد التاريخ التطوري لهذه الآليات عبر ملاحظة انحفاظ البروتينات المفتاحية التي تقوم بكل آلية. على سبيل المثال: Dom34/Hbs1 لهما دور في آلية بلا مضي، Ski7 له دور في آلية بلا توقف،[30] و بروتينات eRF لها دور في آلية بلا معنى.[7] لهذه الغاية تم القيام بأبحاث مكثفة بواسطة برنامج بلاست لتحديد تواجد هذه البروتينات في مختلف الأنواع من الكائنات. تم تحديد أن Hbs1 الخاص بآلية بلا مضي وeRF3 الخاص بآلية بلا معنى يتواجدان فقط لدى حقيقيات النوى. مع ذلك، Dom34 الخاص بآلية بلا مضي متواجد لدى حقيقيات النوى والعتائق، وهذا يوحي بأن آلية بلا مضي هي أول آليات مراقبة الرنا تطورا. يبدو أن Ski7 الخاص بآلية دون توقف محصور بشكل خاص لدى أجناس الخميرة وهذا يوحي بأن آلية بلا توقف هي أحدث آليات المراقبة تطورا، وهذا يترك آلية بلا معنى تلقائيا لتكون ثاني آليات المراقبة في التطور.[37]
ملاحظات
- ( بالإنجليزية premature termination codons (PTCs) وهي كودونات توقف سابقة لأوانها ولا معنى لوجودها في تلك المواضع، ونتجت بسبب خطأ معين.)
المراجع
- Amrani N, Sachs MS, Jacobson A (يونيو 2006)، "Early nonsense: mRNA decay solves a translational problem"، Nature Reviews. Molecular Cell Biology، 7 (6): 415–25، doi:10.1038/nrm1942، PMID 16723977.
- Moore MJ (سبتمبر 2005)، "From birth to death: the complex lives of eukaryotic mRNAs"، Science، 309 (5740): 1514–8، Bibcode:2005Sci...309.1514M، doi:10.1126/science.1111443، PMID 16141059.
- Amrani N, Ganesan R, Kervestin S, Mangus DA, Ghosh S, Jacobson A (نوفمبر 2004)، "A faux 3'-UTR promotes aberrant termination and triggers nonsense-mediated mRNA decay"، Nature، 432 (7013): 112–8، Bibcode:2004Natur.432..112A، doi:10.1038/nature03060، PMID 15525991.
- Fasken MB, Corbett AH (يونيو 2005)، "Process or perish: quality control in mRNA biogenesis"، Nature Structural & Molecular Biology، 12 (6): 482–8، doi:10.1038/nsmb945، PMID 15933735.
- S. Z. Tarun et A. B. Sachs, « Association of the yeast poly(A) tail binding protein with translation initiation factor eIF-4G », في The EMBO journal, vol. 15, 1996-12-16, ص. 7168–7177 ISSN 0261-4189 [النص الكامل, lien PMID, lien PMCID (pages consultées le 2016-10-31)] نسخة محفوظة 14 أكتوبر 2019 على موقع واي باك مشين.
- Chang YF, Imam JS, Wilkinson MF (2007)، "The nonsense-mediated decay RNA surveillance pathway"، Annual Review of Biochemistry، 76: 51–74، doi:10.1146/annurev.biochem.76.050106.093909، PMID 17352659.
- Rehwinkel J, Raes J, Izaurralde E (نوفمبر 2006)، "Nonsense-mediated mRNA decay: Target genes and functional diversification of effectors"، Trends in Biochemical Sciences، 31 (11): 639–46، doi:10.1016/j.tibs.2006.09.005، PMID 17010613.
- Maquat LE (فبراير 2004)، "Nonsense-mediated mRNA decay: splicing, translation and mRNP dynamics"، Nature Reviews. Molecular Cell Biology، 5 (2): 89–99، doi:10.1038/nrm1310، PMID 15040442.
- Holbrook JA, Neu-Yilik G, Hentze MW, Kulozik AE (أغسطس 2004)، "Nonsense-mediated decay approaches the clinic"، Nature Genetics، 36 (8): 801–8، doi:10.1038/ng1403، PMID 15284851.
- Mendell JT, Sharifi NA, Meyers JL, Martinez-Murillo F, Dietz HC (أكتوبر 2004)، "Nonsense surveillance regulates expression of diverse classes of mammalian transcripts and mutes genomic noise"، Nature Genetics، 36 (10): 1073–8، doi:10.1038/ng1429، PMID 15448691.
- Lejeune F, Maquat LE (يونيو 2005)، "Mechanistic links between nonsense-mediated mRNA decay and pre-mRNA splicing in mammalian cells"، Current Opinion in Cell Biology، 17 (3): 309–15، doi:10.1016/j.ceb.2005.03.002، PMID 15901502.
- Conti E, Izaurralde E (يونيو 2005)، "Nonsense-mediated mRNA decay: molecular insights and mechanistic variations across species"، Current Opinion in Cell Biology، 17 (3): 316–25، doi:10.1016/j.ceb.2005.04.005، PMID 15901503.
- Cali BM, Kuchma SL, Latham J, Anderson P (فبراير 1999)، "smg-7 is required for mRNA surveillance in Caenorhabditis elegans"، Genetics، 151 (2): 605–16، PMC 1460488، PMID 9927455.
- Yamashita A, Kashima I, Ohno S (ديسمبر 2005)، "The role of SMG-1 in nonsense-mediated mRNA decay"، Biochimica et Biophysica Acta، 1754 (1–2): 305–15، doi:10.1016/j.bbapap.2005.10.002، PMID 16289965.
- Kim YK, Furic L, Desgroseillers L, Maquat LE (يناير 2005)، "Mammalian Staufen1 recruits Upf1 to specific mRNA 3'UTRs so as to elicit mRNA decay"، Cell، 120 (2): 195–208، doi:10.1016/j.cell.2004.11.050، PMID 15680326.
- Longman D, Plasterk RH, Johnstone IL, Cáceres JF (مايو 2007)، "Mechanistic insights and identification of two novel factors in the C. elegans NMD pathway"، Genes & Development، 21 (9): 1075–85، doi:10.1101/gad.417707، PMC 1855233، PMID 17437990.
- Gatfield D, Unterholzner L, Ciccarelli FD, Bork P, Izaurralde E (أغسطس 2003)، "Nonsense-mediated mRNA decay in Drosophila: at the intersection of the yeast and mammalian pathways"، The EMBO Journal، 22 (15): 3960–70، doi:10.1093/emboj/cdg371، PMC 169044، PMID 12881430.
- Nagy E, Maquat LE (يونيو 1998)، "A rule for termination-codon position within intron-containing genes: when nonsense affects RNA abundance"، Trends in Biochemical Sciences، 23 (6): 198–9، doi:10.1016/S0968-0004(98)01208-0، PMID 9644970.
- Player TJ, Mills DJ, Horton AA (يونيو 1979)، "Lipid peroxidation of the microsomal fraction and extracted microsomal lipids from DAB-induced hepatomas"، British Journal of Cancer، 39 (6): 773–8، doi:10.1128/mcb.18.9.5272، PMC 109113، PMID 9710612.
- Neu-Yilik G, Gehring NH, Thermann R, Frede U, Hentze MW, Kulozik AE (فبراير 2001)، "Splicing and 3' end formation in the definition of nonsense-mediated decay-competent human beta-globin mRNPs"، The EMBO Journal، 20 (3): 532–40، doi:10.1093/emboj/20.3.532، PMC 133467، PMID 11157759.
- Behm-Ansmant I, Gatfield D, Rehwinkel J, Hilgers V, Izaurralde E (مارس 2007)، "A conserved role for cytoplasmic poly(A)-binding protein 1 (PABPC1) in nonsense-mediated mRNA decay"، The EMBO Journal، 26 (6): 1591–601، doi:10.1038/sj.emboj.7601588، PMC 1829367، PMID 17318186.
- Kashima I, Yamashita A, Izumi N, Kataoka N, Morishita R, Hoshino S, Ohno M, Dreyfuss G, Ohno S (فبراير 2006)، "Binding of a novel SMG-1-Upf1-eRF1-eRF3 complex (SURF) to the exon junction complex triggers Upf1 phosphorylation and nonsense-mediated mRNA decay"، Genes & Development، 20 (3): 355–67، doi:10.1101/gad.1389006، PMC 1361706، PMID 16452507.
- Palaniswamy V, Moraes KC, Wilusz CJ, Wilusz J (مايو 2006)، "Nucleophosmin is selectively deposited on mRNA during polyadenylation"، Nature Structural & Molecular Biology، 13 (5): 429–35، doi:10.1038/nsmb1080، PMC 2811576، PMID 16604083.
- Schwartz AM, Komarova TV, Skulachev MV, Zvereva AS, Dorokhov I, Atabekov JG (ديسمبر 2006)، "Stability of plant mRNAs depends on the length of the 3'-untranslated region"، Biochemistry. Biokhimiia، 71 (12): 1377–84، doi:10.1134/s0006297906120145، PMID 17223792.
- Nyikó T, Kerényi F, Szabadkai L, Benkovics AH, Major P, Sonkoly B, Mérai Z, Barta E, Niemiec E, Kufel J, Silhavy D (يوليو 2013)، "Plant nonsense-mediated mRNA decay is controlled by different autoregulatory circuits and can be induced by an EJC-like complex"، Nucleic Acids Research، 41 (13): 6715–28، doi:10.1093/nar/gkt366، PMC 3711448، PMID 23666629.
- Inácio A, Silva AL, Pinto J, Ji X, Morgado A, Almeida F, Faustino P, Lavinha J, Liebhaber SA, Romão L (يوليو 2004)، "Nonsense mutations in close proximity to the initiation codon fail to trigger full nonsense-mediated mRNA decay"، The Journal of Biological Chemistry، 279 (31): 32170–80، doi:10.1074/jbc.m405024200، PMID 15161914.
- Silva AL, Pereira FJ, Morgado A, Kong J, Martins R, Faustino P, Liebhaber SA, Romão L (ديسمبر 2006)، "The canonical UPF1-dependent nonsense-mediated mRNA decay is inhibited in transcripts carrying a short open reading frame independent of sequence context"، RNA، 12 (12): 2160–70، doi:10.1261/rna.201406، PMC 1664719، PMID 17077274.
- Silva AL, Ribeiro P, Inácio A, Liebhaber SA, Romão L (مارس 2008)، "Proximity of the poly(A)-binding protein to a premature termination codon inhibits mammalian nonsense-mediated mRNA decay"، RNA، 14 (3): 563–76، doi:10.1261/rna.815108، PMC 2248256، PMID 18230761.
- Garneau NL, Wilusz J, Wilusz CJ (فبراير 2007)، "The highways and byways of mRNA decay"، Nature Reviews. Molecular Cell Biology، 8 (2): 113–26، doi:10.1038/nrm2104، PMID 17245413.
- van Hoof A, Frischmeyer PA, Dietz HC, Parker R (مارس 2002)، "Exosome-mediated recognition and degradation of mRNAs lacking a termination codon"، Science، 295 (5563): 2262–4، doi:10.1126/science.1067272، PMID 11910110.
- Frischmeyer PA, van Hoof A, O'Donnell K, Guerrerio AL, Parker R, Dietz HC (مارس 2002)، "An mRNA surveillance mechanism that eliminates transcripts lacking termination codons"، Science، 295 (5563): 2258–61، Bibcode:2002Sci...295.2258F، doi:10.1126/science.1067338، PMID 11910109.
- Temperley RJ, Seneca SH, Tonska K, Bartnik E, Bindoff LA, Lightowlers RN, Chrzanowska-Lightowlers ZM (سبتمبر 2003)، "Investigation of a pathogenic mtDNA microdeletion reveals a translation-dependent deadenylation decay pathway in human mitochondria"، Human Molecular Genetics، 12 (18): 2341–8، doi:10.1093/hmg/ddg238، PMID 12915481.
- Karzai AW, Roche ED, Sauer RT (يونيو 2000)، "The SsrA-SmpB system for protein tagging, directed degradation and ribosome rescue"، Nature Structural Biology، 7 (6): 449–55، doi:10.1038/75843، PMID 10881189.
- Doma MK, Parker R (مارس 2006)، "Endonucleolytic cleavage of eukaryotic mRNAs with stalls in translation elongation"، Nature، 440 (7083): 561–4، Bibcode:2006Natur.440..561D، doi:10.1038/nature04530، PMC 1839849، PMID 16554824.
- Kobayashi K, Kikuno I, Kuroha K, Saito K, Ito K, Ishitani R, Inada T, Nureki O (أكتوبر 2010)، "Structural basis for mRNA surveillance by archaeal Pelota and GTP-bound EF1α complex"، Proceedings of the National Academy of Sciences of the United States of America، 107 (41): 17575–9، Bibcode:2010PNAS..10717575K، doi:10.1073/pnas.1009598107، PMC 2955123، PMID 20876129.
- Graille M, Chaillet M, van Tilbeurgh H (مارس 2008)، "Structure of yeast Dom34: a protein related to translation termination factor Erf1 and involved in No-Go decay"، The Journal of Biological Chemistry، 283 (11): 7145–54، doi:10.1074/jbc.M708224200، PMID 18180287.
- Atkinson GC, Baldauf SL, Hauryliuk V (أكتوبر 2008)، "Evolution of nonstop, no-go and nonsense-mediated mRNA decay and their termination factor-derived components"، BMC Evolutionary Biology، 8: 290، doi:10.1186/1471-2148-8-290، PMC 2613156، PMID 18947425.
 بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي