Asteraceae
The family Asteraceae (/ˌæstəˈreɪsi.iː, -si.aɪ/), with the original name Compositae,[5] consists of over 32,000 known species of flowering plants in over 1,900 genera within the order Asterales. Commonly referred to as the aster, daisy, composite, or sunflower family, Compositae were first described in the year 1740. The number of species in Asteraceae is rivaled only by the Orchidaceae, and which is the larger family is unclear as the quantity of extant species in each family is unknown.
| Asteraceae | |
|---|---|
 | |
| Twelve species of Asteraceae from the subfamilies Asteroideae, Carduoideae, and Cichorioideae | |
| Scientific classification | |
| Kingdom: | Plantae |
| Clade: | Tracheophytes |
| Clade: | Angiosperms |
| Clade: | Eudicots |
| Clade: | Asterids |
| Order: | Asterales |
| Family: | Asteraceae Bercht. & J.Presl[2] |
| Type genus | |
| Aster | |
| Subfamilies | |
| |
| Diversity[3] | |
| 1,911 genera | |
| Synonyms[4] | |
|
List
| |
Most species of Asteraceae are annual, biennial, or perennial herbaceous plants, but there are also shrubs, vines, and trees. The family has a widespread distribution, from subpolar to tropical regions, in a wide variety of habitats. Most occur in hot desert and cold or hot semi-desert climates, and they are found on every continent but Antarctica. Their primary common characteristic is flower heads, technically known as capitula, consisting of sometimes hundreds of tiny individual florets enclosed by a whorl of protective involucral bracts.
The oldest known fossils are pollen grains from the Late Cretaceous (Campanian to Maastrichtian) of Antarctica, dated to c. 76–66 million years ago (mya). It is estimated that the crown group of Asteraceae evolved at least 85.9 mya (Late Cretaceous, Santonian) with a stem node age of 88–89 mya (Late Cretaceous, Coniacian).
Asteraceae is an economically important family, providing food staples, garden plants, and herbal medicines. Species outside of their native ranges can be considered weedy or invasive.
Description
Members of the Asteraceae are mostly herbaceous plants, but some shrubs, vines, and trees (such as Lachanodes arborea) do exist. Asteraceae species are generally easy to distinguish from other plants because of their unique inflorescence and other shared characteristics, such as the joined anthers of the stamens.[6] Nonetheless, determining genera and species of some groups such as Hieracium is notoriously difficult (see "damned yellow composite" for example).[7]
Roots
Members of the family Asteraceae generally produce taproots, but sometimes they possess fibrous root systems. Some species have underground stems in the form of caudices or rhizomes. These can be fleshy or woody depending on the species.[5]
Stems
Stems are herbaceous, aerial, branched, and cylindrical with glandular hairs, generally erect, but can be prostrate to ascending. The stems can contain secretory canals with resin,[5] or latex, which is particularly common among the Cichorioideae.[8]
Leaves
Leaves can be alternate, opposite, or whorled. They may be simple, but are often deeply lobed or otherwise incised, often conduplicate or revolute. The margins also can be entire or toothed. Resin[5] or latex[8] also can be present in the leaves.
Inflorescences
Nearly all Asteraceae bear their flowers in dense flower heads called capitula. They are surrounded by involucral bracts, and when viewed from a distance, each capitulum may appear to be a single flower. Enlarged outer (peripheral) flowers in the capitulum may resemble petals, and the involucral bracts may look like a calyx.
Floral heads

In plants of the Asteraceae, what appears to be a single "daisy"-type flower is actually a composite of several much smaller flowers, known as the capitulum or head. By visually presenting as a single flower, the capitulum functions in attracting pollinators, in the same manner that other "showy" flowering plants in numerous other, older, plant families have evolved to attract pollinators. The previous name for the family, Compositae, reflects the fact that what appears to be a single floral entity is in fact a composite of much smaller flowers.[9]
The "petals" or "sunrays" in an "asteraceous" head are in fact individual strap-shaped[10] flowers called ray flowers or ray florets, and the "sun disk" is made up of smaller, radially symmetric, individual flowers called disc flowers or disc florets. The word aster means "star" in Greek, referring to the appearance of most family members as a "celestial body with rays". The capitulum, which often appears to be a single flower, is often referred to as a head.[11] In some species, the entire head is able to pivot its floral stem in the course of the day to track the sun (like a "smart" solar panel), thus maximizing the reflectivity of the entire floral unit and further attracting flying pollinators.[9]
Nearest to the flower stem lie a series of small, usually green, scale-like bracts . These are known as phyllaries; collectively, they form the involucre, which serves to protect the immature head of florets during its development.[9]: 29 The individual florets are arranged atop a dome-like structure called the receptacle.[9]
The individual florets in a head consist, developmentally, of five fused petals (rarely four); instead of sepals, they have threadlike, hairy, or bristly structures,[11] known collectively as a pappus, (plural pappi). The pappus surrounds the ovary and can, when mature and attached to a seed, adhere to animal fur or be carried by air currents, aiding in seed dispersal. The whitish, fluffy head of a dandelion, commonly blown on by children, consists of numerous seeds resting on the receptacle, each seed attached to its pappus. The pappi provide a parachute-like structure to help the seed travel from its point of origin to a more hospitable site.[9]
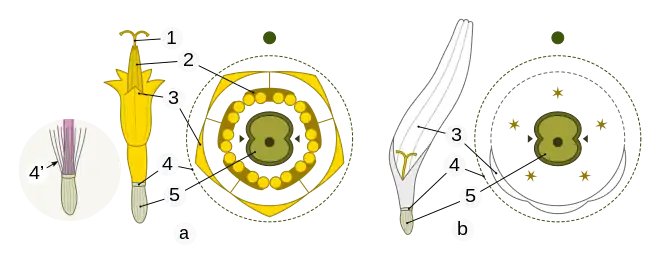
1 – style with stigmas
2 – anthers
3 – corolla (petals); typically, in the ray flower, three petals are joined to form a strap (in other species, five petals can fuse to form a ligule)
4 – reduced calyx
4’ – Carduus acanthoides (left shaded circle): pappus: in many Asteraceae species, the calyx develops as a fibrous or bristly pappus
5 – inferior ovary: fused ovary consisting of two carpels, containing one abaxial ovule (basal placentation).
A ray flower is a two- or three-lobed, strap-shaped, individual flower, found in the head of most members of the Asteraceae.[9][10] The corolla of the ray flower may have two tiny, vestigial teeth, opposite to the three-lobed strap, or tongue, indicating its evolution by fusion from an ancestral, five-part corolla. In some species, the 3:2 arrangement is reversed, with two lobes, and zero or three tiny teeth visible opposite the tongue.
A ligulate flower is a five-lobed, strap-shaped, individual flower found in the heads of certain other asteraceous species.[9] A ligule is the strap-shaped tongue of the corolla of either a ray flower or of a ligulate flower.[10] A disk flower (or disc flower) is a radially symmetric individual flower in the head, which is ringed by the ray flowers when both are present.[9][10] In some species, ray flowers may be arranged around the disc in irregular symmetry, or with a weakly bilaterally symmetric arrangement.[9]
Variations
A radiate head has disc flowers surrounded by ray flowers. A ligulate head has all ligulate flowers and no disc flowers. When an Asteraceae flower head has only disc flowers that are either sterile, male, or bisexual (but not female and fertile), it is a discoid head.
Disciform heads possess only disc flowers in their heads, but may produce two different sex types (male or female) within their disciform head.
Some other species produce two different head types: staminate (all-male), or pistillate (all-female). In a few unusual species, the "head" will consist of one single disc flower; alternatively, a few species will produce both single-flowered female heads, along with multi-flowered male heads, in their "pollination strategy".[9]
Floral structures
.jpg.webp)
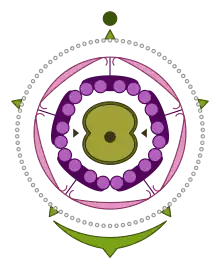
The distinguishing characteristic of Asteraceae is their inflorescence, a type of specialised, composite flower head or pseudanthium, technically called a calathium or capitulum,[12][13] that may look superficially like a single flower. The capitulum is a contracted raceme composed of numerous individual sessile flowers, called florets, all sharing the same receptacle.
A set of bracts forms an involucre surrounding the base of the capitulum. These are called "phyllaries", or "involucral bracts". They may simulate the sepals of the pseudanthium. These are mostly herbaceous but can also be brightly coloured (e.g. Helichrysum) or have a scarious (dry and membranous) texture. The phyllaries can be free or fused, and arranged in one to many rows, overlapping like the tiles of a roof (imbricate) or not (this variation is important in identification of tribes and genera).
Each floret may be subtended by a bract, called a "palea" or "receptacular bract". These bracts are often called "chaff". The presence or absence of these bracts, their distribution on the receptacle, and their size and shape are all important diagnostic characteristics for genera and tribes.
The florets have five petals fused at the base to form a corolla tube and they may be either actinomorphic or zygomorphic. Disc florets are usually actinomorphic, with five petal lips on the rim of the corolla tube. The petal lips may be either very short, or long, in which case they form deeply lobed petals. The latter is the only kind of floret in the Carduoideae, while the first kind is more widespread. Ray florets are always highly zygomorphic and are characterised by the presence of a ligule, a strap-shaped structure on the edge of the corolla tube consisting of fused petals. In the Asteroideae and other minor subfamilies these are usually borne only on florets at the circumference of the capitulum and have a 3+2 scheme – above the fused corolla tube, three very long fused petals form the ligule, with the other two petals being inconspicuously small. The Cichorioideae has only ray florets, with a 5+0 scheme – all five petals form the ligule. A 4+1 scheme is found in the Barnadesioideae. The tip of the ligule is often divided into teeth, each one representing a petal. Some marginal florets may have no petals at all (filiform floret).
The calyx of the florets may be absent, but when present is always modified into a pappus of two or more teeth, scales or bristles and this is often involved in the dispersion of the seeds. As with the bracts, the nature of the pappus is an important diagnostic feature.
There are usually four or five stamens.[11] The filaments are fused to the corolla, while the anthers are generally connate (syngenesious anthers), thus forming a sort of tube around the style (theca). They commonly have basal and/or apical appendages. Pollen is released inside the tube and is collected around the growing style, and then, as the style elongates, is pushed out of the tube (nüdelspritze).
The pistil consists of two connate carpels. The style has two lobes. Stigmatic tissue may be located in the interior surface or form two lateral lines. The ovary is inferior and has only one ovule, with basal placentation.
Fruits and seeds
In members of the Asteraceae the fruit is achene-like, and is called a cypsela (plural cypselae). Although there are two fused carpels, there is only one locule, and only one seed per fruit is formed.[11] It may sometimes be winged or spiny because the pappus, which is derived from calyx tissue often remains on the fruit (for example in dandelion). In some species, however, the pappus falls off (for example in Helianthus). Cypsela morphology is often used to help determine plant relationships at the genus and species level.[14] The mature seeds usually have little endosperm or none.[6]
Pollen
The pollen of composites is typically echinolophate, a morphological term meaning "with elaborate systems of ridges and spines dispersed around and between the apertures."[15]
Metabolites
In Asteraceae, the energy store is generally in the form of inulin rather than starch. They produce iso/chlorogenic acid, sesquiterpene lactones, pentacyclic triterpene alcohols, various alkaloids, acetylenes (cyclic, aromatic, with vinyl end groups), tannins. They have terpenoid essential oils that never contain iridoids.[16]
Asteraceae produce secondary metabolites, such as flavonoids and terpenoids. Some of these molecules can inhibit protozoan parasites such as Plasmodium, Trypanosoma, Leishmania and parasitic intestinal worms, and thus have potential in medicine.[17]
Taxonomy
History
Compositae, the original name for Asteraceae, were first described in 1740 by Dutch botanist Adriaan van Royen.[18]: 117–118 Traditionally, two subfamilies were recognised: Asteroideae (or Tubuliflorae) and Cichorioideae (or Liguliflorae).[19]: 242 The latter has been shown to be extensively paraphyletic, and has now been divided into 12 subfamilies, but the former still stands.[20] The study of this family is known as synantherology.
Phylogeny
The phylogenetic tree of subfamilies presented below is based on Panero & Funk (2002)[20] updated in 2014,[21] and now also includes the monotypic Famatinanthoideae.[21][22][23] The diamond (♦) denotes a very poorly supported node (<50% bootstrap support), the dot (•) a poorly supported node (<80%).[16]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The family includes over 32,000 currently accepted species, in over 1,900 genera (list) in 13 subfamilies.[3] The number of species in the family Asteraceae is rivaled only by Orchidaceae.[16][24] Which is the larger family is unclear, because of the uncertainty about how many extant species each family includes. The four subfamilies Asteroideae, Cichorioideae, Carduoideae and Mutisioideae contain 99% of the species diversity of the whole family (approximately 70%, 14%, 11% and 3% respectively).
Because of the morphological complexity exhibited by this family, agreeing on generic circumscriptions has often been difficult for taxonomists. As a result, several of these genera have required multiple revisions.[6]
Paleontology and evolutionary processes
The oldest known fossils of members of Asteraceae are pollen grains from the Late Cretaceous of Antarctica, dated to ~76–66 mya (Campanian to Maastrichtian) and assigned to the extant genus Dasyphyllum. Barreda, et al. (2015) estimated that the crown group of Asteraceae evolved at least 85.9 mya (Late Cretaceous, Santonian) with a stem node age of 88–89 mya (Late Cretaceous, Coniacian).[1]
It is not known whether the precise cause of their great success was the development of the highly specialised capitulum, their ability to store energy as fructans (mainly inulin), which is an advantage in relatively dry zones, or some combination of these and possibly other factors.[16] Heterocarpy, or the ability to produce different fruit morphs, has evolved and is common in Asteraceae. It allows seeds to be dispersed over varying distances and each is adapted to different environments, increasing chances of survival.[25]
Etymology and pronunciation
The original name Compositae is still valid under the International Code of Nomenclature for algae, fungi, and plants.[26] It refers to the "composite" nature of the capitula, which consist of a few or many individual flowers.
The alternative (as it came later) name Asteraceae (English: /ˌæstəˈreɪsi, -siˌaɪ, -siˌeɪ, -siˌiː/) comes to international scientific vocabulary from Neo-Latin, from Aster, the type genus, + -aceae,[27] a standardized suffix for plant family names in modern taxonomy. This genus name comes from the Classical Latin word aster, "star", which came from Ancient Greek ἀστήρ (astḗr), "star".[27] It refers to the star-like form of the inflorescence.
The vernacular name daisy, widely applied to members of this family, is derived from the Old English name of the daisy (Bellis perennis): dæġes ēaġe, meaning "day's eye". This is because the petals open at dawn and close at dusk.[28]
Distribution and habitat
Asteraceae species have a widespread distribution, from subpolar to tropical regions in a wide variety of habitats. Most occur in hot desert and cold or hot semi-desert climates, and they are found on every continent but Antarctica. They are especially numerous in tropical and subtropical regions (notably Central America, eastern Brazil, the Mediterranean, the Levant, southern Africa, central Asia, and southwestern China).[24] The largest proportion of the species occur in the arid and semi-arid regions of subtropical and lower temperate latitudes.[5] The Asteraceae family comprises 10% of all flowering plant species.[7]
Ecology


Asteraceae are especially common in open and dry environments.[6] Many members of Asteraceae are pollinated by insects, which explains their value in attracting beneficial insects, but anemophily is also present (e.g. Ambrosia, Artemisia). There are many apomictic species in the family.
Seeds are ordinarily dispersed intact with the fruiting body, the cypsela. Anemochory (wind dispersal) is common, assisted by a hairy pappus. Epizoochory is another common method, in which the dispersal unit, a single cypsela (e.g. Bidens) or entire capitulum (e.g. Arctium) has hooks, spines or some structure to attach to the fur or plumage (or even clothes, as in the photo) of an animal just to fall off later far from its mother plant.
Some members of Asteraceae are economically important as weeds. Notable in the United States are Senecio jacobaea (ragwort),[29] Senecio vulgaris (groundsel),[30] and Taraxacum (dandelion).[31] Some are invasive species in particular regions, often having been introduced by human agency. Examples include various tumbleweeds, Bidens, ragweeds, thistles, and dandelion.[32] Dandelion was introduced into North America by European settlers who used the young leaves as a salad green.[33] A number of species are toxic to grazing animals.[11]
Uses

Asteraceae is an economically important family, providing products such as cooking oils, leaf vegetables like lettuce, sunflower seeds, artichokes, sweetening agents, coffee substitutes and herbal teas. Several genera are of horticultural importance, including pot marigold (Calendula officinalis), Echinacea (coneflowers), various daisies, fleabane, chrysanthemums, dahlias, zinnias, and heleniums. Asteraceae are important in herbal medicine, including Grindelia, yarrow, and many others.[36]
Commercially important plants in Asteraceae include the food crops Lactuca sativa (lettuce), Cichorium (chicory), Cynara scolymus (globe artichoke), Helianthus annuus (sunflower), Smallanthus sonchifolius (yacón), Carthamus tinctorius (safflower) and Helianthus tuberosus (Jerusalem artichoke).[37]
Plants are used as herbs and in herbal teas and other beverages. Chamomile, for example, comes from two different species: the annual Matricaria chamomilla (German chamomile) and the perennial Chamaemelum nobile (Roman chamomile). Calendula (known as pot marigold) is grown commercially for herbal teas and potpourri. Echinacea is used as a medicinal tea. The wormwood genus Artemisia includes absinthe (A. absinthium) and tarragon (A. dracunculus). Winter tarragon (Tagetes lucida), is commonly grown and used as a tarragon substitute in climates where tarragon will not survive.[38]
Many members of the family are grown as ornamental plants for their flowers, and some are important ornamental crops for the cut flower industry. Some examples are Chrysanthemum, Gerbera, Calendula, Dendranthema, Argyranthemum, Dahlia, Tagetes, Zinnia, and many others.[39]
.jpg.webp)
Many species of this family possess medicinal properties and are used as traditional antiparasitic medicine.[17]
Members of the family are also commonly featured in medical and phytochemical journals because the sesquiterpene lactone compounds contained within them are an important cause of allergic contact dermatitis. Allergy to these compounds is the leading cause of allergic contact dermatitis in florists in the US.[41] Pollen from ragweed Ambrosia is among the main causes of so-called hay fever in the United States.[42]
Asteraceae are also used for some industrial purposes. French Marigold (Tagetes patula) is common in commercial poultry feeds and its oil is extracted for uses in cola and the cigarette industry. The genera Chrysanthemum, Pulicaria, Tagetes, and Tanacetum contain species with useful insecticidal properties. Parthenium argentatum (guayule) is a source of hypoallergenic latex.[39]
Several members of the family are copious nectar producers[39] and are useful for evaluating pollinator populations during their bloom. Centaurea (knapweed), Helianthus annuus (domestic sunflower), and some species of Solidago (goldenrod) are major "honey plants" for beekeepers. Solidago produces relatively high protein pollen, which helps honey bees over winter.[43]
References
- Barreda, Viviana D.; Palazzesi, Luis; Tellería, Maria C.; Olivero, Eduardo B.; Raine, J. Ian; Forest, Félix (2015). "Early evolution of the angiosperm clade Asteraceae in the Cretaceous of Antarctica". Proceedings of the National Academy of Sciences of the United States of America. 112 (35): 10989–10994. Bibcode:2015PNAS..11210989B. doi:10.1073/pnas.1423653112. PMC 4568267. PMID 26261324.
- "Asteraceae Bercht. & J.Presl". Tropicos. Missouri Botanical Garden. Retrieved 14 July 2017.
- "Compositae". The Plant List (www.theplantlist.org). Royal Botanic Gardens, Kew & Missouri Botanical Garden. 2013. Archived from the original on 11 February 2012. Retrieved 18 November 2016.
- "Family: Asteraceae Bercht. & J. Presl, nom. cons". Germplasm Resources Information Network (GRIN) (www.ars-grin.gov). Beltsville, Maryland: USDA, ARS, National Genetic Resources Program, National Germplasm Resources Laboratory. Archived from the original on 17 September 2008. Retrieved 12 June 2008.
- Barkley, T.M.; Brouillet, L.; Strother, J.L. (2006). "Asteraceae". In Flora of North America Editorial Committee (ed.). Flora of North America North of Mexico (FNA). Vol. 19. New York and Oxford: Oxford University Press. Retrieved 20 March 2022 – via eFloras.org, Missouri Botanical Garden, St. Louis, MO & Harvard University Herbaria, Cambridge, MA.
- Judd, W.S.; Campbell, C.S.; Kellogg, E.A.; Stevens, P.F.; Donaghue, M.J. (2007). Plant Systematics: A Phylogenetic Approach (3rd ed.). Sunderland, Massachusetts: Sinauer Associates. ISBN 978-0878934072.
- Mandel, Jennifer R.; Dikow, Rebecca B.; Siniscalchi, Carolina M.; Thapa, Ramhari; Watson, Linda E.; Funk, Vicki A. (9 July 2019). "A fully resolved backbone phylogeny reveals numerous dispersals and explosive diversifications throughout the history of Asteraceae". Proceedings of the National Academy of Sciences of the United States of America. National Academy of Sciences. 116 (28): 14083–14088. Bibcode:2019PNAS..11614083M. doi:10.1073/pnas.1903871116. ISSN 1091-6490. PMC 6628808. PMID 31209018.
- Kilian, Norbert; Gemeinholzer, Birgit; Lack, Hans Walter. "24. Cichorieae" (PDF). In Funk, V.A.; Susanna, A.; Stuessy, T.E.; Bayer, R.J. (eds.). Systematics, evolution and biogeography of Compositae. Vienna: International Association for Plant Taxonomy. Archived (PDF) from the original on 22 June 2021. Retrieved 20 February 2021.
- Morhardt, Sia; Morhardt, Emil (2004). California desert flowers: an introductions to families, genera, and species. Berkeley, Los Angeles, and London: University of California Press. pp. 29–32. ISBN 978-0520240032.
- MacKay, Pam (2013). Mojave Desert Wildflowers: A Field Guide To Wildflowers, Trees, And Shrubs Of The Mojave Desert, Including The Mojave National Preserve, Death Valley National Park, and Joshua Tree National Park (Wildflower Series). Guilford, Connecticut: FalconGuides. p. 35 (figure 5). ISBN 978-0762780334.
- Spellenberg, Richard (2001) [1979]. National Audubon Society Field Guide to North American Wildflowers: Western Region (rev ed.). Knopf. p. 351. ISBN 978-0-375-40233-3 – via Internet Archive.
- Beentje, Henk (2010). The Kew Plant Glossary, an illustrated dictionary of plant terms. Richmond, U.K.: Kew Publishing. ISBN 978-1842464229.
- Usher, G. (1966). A Dictionary of Botany, including terms used in biochemistry, soil science, and statistics. London: Constable and Company, Ltd. ISBN 0094504903. OCLC 959412625.
- McKenzie, R.J.; Samuel, J.; Muller, E.M.; Skinner, A.K.W.; Barker, N.P. (2005). "Morphology of cypselae in subtribe Arctotidinae (Compositae–Arctotideae) and its taxonomic implications". Annals of the Missouri Botanical Garden. Missouri Botanical Garden Press. 92 (4): 569–594. JSTOR 40035740. Retrieved 18 February 2021 – via ResearchGate.
- Tomb, A. Spencer; Larson, Donald A.; Skvarla, John J. (1 May 1974). "Pollen morphology and detailed structure of family Compositae, tribe Cichorieae. I. Subtribe Stephanomeriinae". American Journal of Botany. Wiley. 61 (5): 486–498. doi:10.1002/j.1537-2197.1974.tb10788.x. JSTOR 2442020.
- Stevens, Peter F. (2022) [2001]. "Angiosperm Phylogeny Website: Asterales". Angiosperm Phylogeny Website. Archived from the original on 29 November 2022. Retrieved 15 January 2023.
- Panda, Sujogya Kumar; Luyten, Walter (2018). "Antiparasitic activity in Asteraceae with special attention to ethnobotanical use by the tribes of Odisha, India". Parasite. 25: 10. doi:10.1051/parasite/2018008. ISSN 1776-1042. PMC 5847338. PMID 29528842.
- von Royen, Adriani (1740). Florae leydensis prodromus : exhibens plantas quae in Horto academico Lugduno-Batavo aluntur (in Latin). Lugduni Batavorum [Leiden]: Apud Samuelen Luchtmans academiae typographum. Archived from the original on 19 July 2021. Retrieved 18 February 2021 – via Botanicus.
- Bayer, Randall J.; Starr, Julian R. (1998). "Tribal phylogeny of the Asteraceae based on two non-coding chloroplast sequences, the trnL Intron and trnL/trnF Intergenic Spacer". Annals of the Missouri Botanical Garden. Missouri Botanical Garden Press. 85 (2): 242–256. doi:10.2307/2992008. JSTOR 2992008.
- Panero, J.L.; Funk, V.A. (2002). "Toward a phylogenetic subfamilial classification for the Compositae (Asteraceae)". Proceedings of the Biological Society of Washington. 115: 909–922. Archived from the original on 7 May 2021. Retrieved 16 February 2021 – via Biodiversity Heritage Library.
- Panéro, José L.; Freire, Susana E.; Ariza Espinar, Luis; Crozier, Bonnie S.; Barboza, Gloria E.; Cantero, Juan J. (2014). "Resolution of deep nodes yields an improved backbone phylogeny and a new basal lineage to study early evolution of Asteraceae". Molecular Phylogenetics and Evolution. 80 (1): 43–53. doi:10.1016/j.ympev.2014.07.012. hdl:11336/36507. PMID 25083940. S2CID 6745704 – via ResearchGate (abstract and introduction).
- Fu, Zhi-Xi; Jiao, Bo-Han; Nie, Bao; Zhang, Guo-Jin; Gao, Tian-Gang (22 July 2016). "A comprehensive generic‐level phylogeny of the sunflower family: Implications for the systematics of Chinese Asteraceae". Journal of Systematics and Evolution. 54 (4): 416–437. doi:10.1111/jse.12216. S2CID 89115861 – via ResearchGate.
- Funk, Vicki A.; Fragman-Sapir, Ori (2009). "22. Gymnarrheneae (Gymnarrhenoideae)" (PDF). In V.A. Funk; A. Susanna; T. Stuessy; R. Bayer (eds.). Systematics, Evolution, and Biogeography of Compositae. Vienna: International Association for Plant Taxonomy. pp. 327–332. ISBN 978-3950175431. Archived (PDF) from the original on 6 May 2021. Retrieved 16 February 2021.
- Panero, Jose L.; Crozier, Bonnie S. (27 January 2012). "Asteraceae. Sunflowers, daisies". The Tree of Life Web Project (tolweb.org). Archived from the original on 21 January 2022. Retrieved 16 February 2021.
- Gardocki, Mary E.; Zablocki, Heather; El-Keblawy, Ali; Freeman, D. Carl (2000). "Heterocarpy in Calendula micrantha (Asteraceae): the effects of competition and availability of water on the performance of offspring from different fruit morphs". Evolutionary Ecology Research. 2 (6): 701–718. Retrieved 16 February 2021 – via ResearchGate.
- "International Code of Nomenclature for algae, fungi, and plants – Article 18.5". iapt-taxon.org. Archived from the original on 27 September 2013. Retrieved 2 June 2015.
- Merriam-Webster. Merriam-Webster's Unabridged Dictionary. Merriam-Webster. Archived from the original on 25 May 2020. Retrieved 27 July 2016.
- "daisy, n.". Home : Oxford English Dictionary. OED Online. Oxford University Press. March 2021. Archived from the original on 15 January 2023. Retrieved 23 April 2021.
- "Tansy ragwort Jacobaea vulgaris". Washington State Noxious Weed Control Board (www.nwcb.wa.gov). Archived from the original on 4 December 2020. Retrieved 21 January 2021.
- "Common groundsel Senecio vulgaris". Washington State Noxious Weed Control Board (www.nwcb.wa.gov). Archived from the original on 30 January 2021. Retrieved 21 January 2021.
- González-Castejón, Marta; Visioli, Francesco; Rodriguez-Casado, Arantxa (17 August 2012). "Diverse biological activities of dandelion". Nutrition Reviews. 70 (9): 534–547. doi:10.1111/j.1753-4887.2012.00509.x. ISSN 0029-6643. PMID 22946853.
- Martín-Forés, Irene; Acosta-Gallo, Belén; Castro, Isabel; de Miguel, José M.; del Pozo, Alejandro; Casado, Miguel A. (14 June 2018). "The invasiveness of Hypochaeris glabra (Asteraceae): Responses in morphological and reproductive traits for exotic populations". PLOS ONE. 13 (6): e0198849. Bibcode:2018PLoSO..1398849M. doi:10.1371/journal.pone.0198849. ISSN 1932-6203. PMC 6002075. PMID 29902275.
- "Taraxacum officinale: dandelion". Invasive Plant Atlas of the United States (www.invasiveplantatlas.org). Archived from the original on 23 October 2012. Retrieved 10 September 2012.
- Talhouk, S.N.; Fabian, M.; Dagher, R. (2015). "Senecio angulatus (Cape ivy, Climbing groundsel, Creeping groundsel)". AUB Landscape Plant Database (landscapeplants.aub.edu.lb). Beirut, Lebanon: Department of Landscape Design & Ecosystem Management, American University of Beirut. Archived from the original on 15 May 2021. Retrieved 16 February 2021.
- "Senecio angulatus – climbing groundsel". Brisbane City Council weed identification tool (weeds.brisbane.qld.gov.au). Brisbane, Queensland, Australia: Brisbane City Council. Archived from the original on 2 March 2021. Retrieved 16 February 2021.
- Michel, Jennifer; Abd Rani, Nur Zahirah; Husain, Khairana (2020). "A review on the potential use of medicinal plants from Asteraceae and Lamiaceae plant family in cardiovascular diseases". Frontiers in Pharmacology. 11: 852. doi:10.3389/fphar.2020.00852. ISSN 1663-9812. PMC 7291392. PMID 32581807.
- Ezcurra, Exequiel (May 2020). "12. Asteraceae: The sunflower family" (PDF). Spring Wildflowers: An Introduction to the Native Flora of Southern California (Lecture notes by Exequiel Ezcurra). Archived (PDF) from the original on 15 January 2023. Retrieved 15 January 2022.
- "Winter Tarragon, Sweet Mace Tagetes lucida". Jekka's. Avon, Bristol, U.K. Archived from the original on 12 December 2021. Retrieved 2 May 2022.
- Singh, Rajendra; Singh, Garima; Tiwari, Ajeet; Patel, Shveta; Agrawal, Ruhi; Sharma, Akhilesh; Singh, B. (2015). "Diversity of host plants of aphids (Homoptera: Aphididae) infesting Asteraceae in India". International Journal of Zoological Investigations. 1 (2): 137–167. ISSN 2454-3055. Retrieved 16 February 2021 – via ResearchGate.
- Auld, B.A.; Meld, R.W. (1992). Weeds: An Illustrated Botanical Guide to the Weeds of Australia. Melbourne: Inkata Press. p. 264. ISBN 978-0909605377.
- Odom, R.B.; James, W.D.; Berger, T.G. (2000). Andrews' Diseases of the Skin: Clinical Dermatology. Philadelphia, Pennsylvania: W.B. Saunders Company. p. 1135. ISBN 978-0721658322.
- Asthma and Allergy Foundation of America. "Ragweed Allergy". www.aafa.org. Archived from the original on 7 October 2007. Retrieved 10 November 2007.
-
"Five Plants to Help Pollinators". www.storey.com. The Xerces Society. 2016. Archived from the original on 22 September 2020. Retrieved 2 June 2020.
Goldenrods are among the most important late-season pollinator plants.
Bibliography
- Funk, Vicki A.; Susanna, A.; Stuessy, T.F.; Bayer, R.J., eds. (2009). Systematics, Evolution, and Biogeography of Compositae. Vienna: International Association for Plant Taxonomy. ISBN 978-3-9501754-3-1. Archived from the original on 15 January 2023. Retrieved 12 November 2021 – via Google Books. (Also available here at Internet Archive)
External links
 Media related to Asteraceae at Wikimedia Commons
Media related to Asteraceae at Wikimedia Commons Data related to Asteraceae at Wikispecies
Data related to Asteraceae at Wikispecies
- Asteraceae at the Angiosperm Phylogeny Website
- Compositae.org – Compositae Working Group (CWG) and Global Compositae Database (GCD)