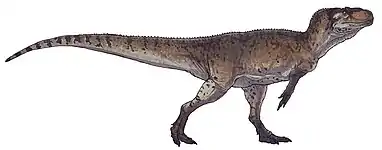
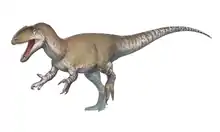Monolophosaurus
Monolophosaurus (/ˌmɒnoʊˌlɒfoʊˈsɔːrəs/ MON-oh-LOF-oh-SOR-əs;[1] meaning "single-crested lizard") is an extinct genus of tetanuran theropod dinosaur from the Middle Jurassic Shishugou Formation in what is now Xinjiang, China.[2][3] It was named for the single crest on top of its skull. Monolophosaurus was a mid-sized theropod at about 5–5.5 metres (16–18 ft) long and weighed 475 kilograms (1,047 lb).
| Monolophosaurus Temporal range: Middle Jurassic, | |
|---|---|
 | |
| Mounted skeleton in Japan | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Clade: | Tetanurae |
| Genus: | †Monolophosaurus Zhao & Currie 1993 |
| Species: | †M. jiangi |
| Binomial name | |
| †Monolophosaurus jiangi Zhao & Currie 1993 | |
Discovery and naming

A nearly complete skeleton of a theropod new to science was discovered by Dong Zhiming in 1981, during stratigraphic exploration for the benefit of the oil industry. The fossil was not unearthed until 1984. In 1987, before description in the scientific literature, it was referred to in the press as Jiangjunmiaosaurus, an invalid nomen nudum.[2][4] In 1992 it was mentioned by Dong Zhiming as Monolophosaurus jiangjunmiaoi,[5] and in 1993 by Wayne Grady as Monolophosaurus dongi.[6] These latter names also lacked a description and therefore were nomina nuda as well.
In 1993/1994, Zhao Xijin and Philip John Currie named and described the type species Monolophosaurus jiangi. The generic name is derived from Greek μόνος, monos, "single", and λόφος or λόφη, lophos/lophè, "crest", in reference to the single crest on the snout. The specific name refers to Jiangjunmiao, an abandoned desert inn near which the fossil was found.[2][7] Jiangjunmiao means "the temple (miao) of the general (jiangjun)"; local legend has it that a general was buried here.
The holotype IVPP 84019 was discovered in the Junggar Basin, in layers of the Wucaiwan Formation dating from the Bathonian-Callovian. It consists of a rather complete skeleton including the skull, lower jaws, vertebral column and pelvis. The rear of the tail, the shoulder girdle and the limbs are lacking. It represents an adult or subadult individual.[2] The type specimen was restored with plaster to be used in a travelling exhibit. Its left side was encased in foam which has hindered subsequent study. A reconstruction was made of the missing elements to create casts of complete skeletal mounts. In 2010, two studies by Stephen Brusatte, et al redescribed the holotype, at the time still the only specimen known, in detail.[7][8]
In 2006, Thomas Carr suggested that Guanlong, another theropod with a large, thin, and fenestrated midline crest and from the same formation, was in fact a subadult individual of Monolophosaurus. Usually Guanlong had been considered a proceratosaurid tyrannosauroid, but Carr had performed an analysis in which both specimens clustered and were allosauroids.[9] More conservatively, in 2010 Gregory S. Paul renamed Guanlong into a Monolophosaurus species, Monolophosaurus wucaii,[10] presuming the taxa might be sister species. In 2010, Brusatte et al rejected the identity, pointing out that the Guanlong holotype was actually a fully adult individual.[7]
Description

The type and only known individual has been estimated at five metres (16.4 ft).[3] In 2010, Paul estimated the length at 5.5 metres (18 ft) and the weight at 475 kilograms (1,047 lb).[10]
Several distinguishing traits have been established. The snout on its midline bears a large crest, the front of which is formed by the praemaxillan. It continues to behind over the nasals and lacrimals; its rear touches the frontals. The top of the crest runs parallel to the upper jaw edge. The ascending branches of the praemaxillae each have a forked rear. The side of the praemaxilla features a deep groove running from an opening in the ascending branch towards an opening below the nostril. Within the depression around the upper rear side of the nostril two pneumatic openings are present, of unequal size. The rear branch of the lacrimal, above the eye socket, has a distinctive hatchet-shaped process pointing upwards. The combined frontals are rectangular and elongated with a length:width ratio of 1.67.[7]
Skull

The holotype skull has a length of eighty centimetres. It is as such rather flat but this is obscured by the presence of a large snout crest occupying about three quarters of the skull length, reaching the level of the eye sockets. Sprouting from the praemaxillae at the snout tip, the crest is largely formed by the nasal bones. Transversely it has a triangular cross-section with a broad base and a more narrow top; this does not form a ridge, however, but has a flat upper surface. The nasal crest side is very rugose with a series of bosses and swellings. The nasal bone contributes to the upper rear part of the depression around the antorbital fenestra. This area shows a number of pneumatic openings or pneumatopores, where diverticula of the air sacks entered the bone. In the front two small foramina are present, more to the rear two large horizontal oval openings. CAT scans showed that internally the nasal bone is heavily pneumatised, with large air chambers. Also the jugal bone is pneumatised. The lacrimal is I-shaped. It has an ascending branch forming the vertical rear edge of the crest; due to the triangular cross-section this branch is inclined towards the midline of the skull. The upper outer side of this branch forms a rectangular boss. Behind the eye socket, on the postorbital another, smaller, horn is present. The frontal bones do not contribute to the crest; they are unique among Theropoda in having a combined rectangular instead of triangular shape, due to the posterior position of the crest rear.[7]
The praemaxilla has a narrow ascending branch, forming the front of the crest. The rear of this branch is forked and embraces a lateral point of the nasal, a feature not recognised in the original description of 1994. At the base of the branch a small opening is present. A larger opening is located below the nostril. Both openings are connected by a distinct groove, curving around the underside of the nostril. The function of this unique trait is unknown. The praemaxilla bears four teeth. The maxilla bears thirteen teeth. The maxilla has a short depression around the lower front of the antorbital fenestra. Within this area a smaller hollowing is located, closed at the inside, perhaps representing the fenestra promaxillaris, of which it has the usual position, or the fenestra maxillaris, the normal identity of a single opening.[7]
In the braincase, the channel of the nervus trigeminus, the fifth brain nerve, is not bifurcated. The palatine bone is pneumatised, as shown by the presence of a pneumatopore. In the lower jaw, the external mandibular fenestra is rather small for a basal tetanuran. The holotype shows eighteen teeth in the right dentary, seventeen in the left dentary; such an asymmetry is not rare among large theropods. A row of foramina is present below and on the outer side of the tooth row. These openings are relatively large below the first four teeth; more to behind, they become smaller and their row curves downwards. From the ninth tooth onwards, the foramina merge into a groove. A second row of openings runs parallel to the lower jaw edge and ends at the thirteenth tooth position, which is exceptionally far. At the inside of the dentary, the Meckelian groove at the level of the third tooth extends to the front into two superimposed narrow slits. The rear of the lower jaw shows a unique combination of a kinked suture between the angular and the surangular, and the basal trait of the surangular reaching the rear jaw edge. The rather small foramen surangulare posterior is not overhung by a thick bone shelf, which is rare among large theropods.[7]
Postcranial skeleton

The vertebral column consists of nine cervical vertebrae, fourteen dorsals and five sacrals. The number of tail vertebrae is unknown. The cervical vertebrae of the neck are strongly pneumatised. They possess pleurocoels at their sides and their insides are hollowed out by large air chambers. The neural spines of the cervical vertebrae are narrow in side view and decreased in width towards the rear: those of the eighth and ninth vertebrae were rod-like. At least the first three dorsal vertebrae of the back have pleurocoels as well. The dorsals are connected by robust hyposphene-hypantrum complexes. From the sixth vertebra onwards the neural spines abruptly become wider. The neural spines of the sacral vertebrae are not fused into a supraneural plate. The tail base is slightly oriented downwards. The caudal vertebrae of the tail base also show hyposphene-hypantrum complexes.[8]
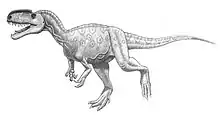
In the pelvis, the ilium has a slightly convex upper profile. Its front blade has a pending, hook-shaped point. The edge of the base of the front blade is incised by a groove. The ilium has some basal traits. The process to which the pubic bone is attached, has two facets, one directed to below, the other obliquely pointing to the front, instead of a single facet. Also basal is the fact that the hip joint is overhung by a hood-shaped extension of the antitrochanter; the front of this hood reaches further to below and to the outer side. There is no clear brevis shelf. The pubic bones and the ischia resemble each other in having a "foot" and being per pair connected via bony skirts, pierced by a foramen.[8]
Classification
Monolophosaurus was originally termed a "megalosaur" and has often since been suggested to be an allosauroid. Smith et al. (2007) was the first publication to find Monolophosaurus to be a non-neotetanuran tetanuran,[11] by noting many characters previously thought to be exclusive of Allosauroidea to have a more wider distribution. Also, Zhao et al. in 2010 noted various primitive features of the skeleton suggesting that Monolophosaurus could be one of the most basal tetanuran dinosaurs instead.[8] Benson (2008, 2010) placed Monolophosaurus in a clade with Chuandongocoelurus that is more basal than Megalosauridae and Spinosauridae in the Megalosauroidea.[12][13] Later, Benson et al. (2010) found the Chuandongocoelurus/Monolophosaurus clade to be outside of Megalosauroidea and Neotetanurae, near the base of Tetanurae.[7] A 2012 phylogeny found Monolophosaurus and Chuandongocoelurus, while not sister taxa, to form a group outside more derived groups at the base of Tetanurae.[14]


The following cladogram is based on the phylogenetic analysis conducted by Carrano in 2012, showing the relationships of Monolophosaurus:[14]
| Neotheropoda |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
In 2019, Rauhut found Monolophosaurus to be the most basal member of Carnosauria.[15]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||



A 2023 examination of Irritator challengeri found Monolophosaurus to be a sister taxon to Spinosauridae as part of a larger, monophyletic Carnosauria.[16]
Paleobiology
The type specimen (IVPP 84019) had its tenth and possibly eleventh neural spines fractured. They are fused together. A series of parallel ridges on one of the specimen's dentaries may represent tooth marks.[17]
References
- Creisler, Ben (July 7, 2003). "Dinosauria Translation and Pronunciation Guide M". Archived from the original on September 28, 2007. Retrieved August 23, 2010.
- Zhao, Xi-Jin; Currie, Philip J. (1993). "A large crested theropod from the Jurassic of Xinjiang, People's Republic of China" (PDF). Canadian Journal of Earth Sciences. 30 (10): 2027–2036. Bibcode:1993CaJES..30.2027Z. doi:10.1139/e93-178.
- Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2010 Appendix.
- Holley, David (October 23, 1987). "2nd creature was meat-eater: fossil remains of huge dinosaur found in China". Los Angeles Times. Retrieved August 23, 2010.
- Dong, Z., 1992, Dinosaurian Faunas of China, Ocean Press/Springer-Verlag, Beijing/Berlin. 188 pp
- Grady, W., 1993, The Dinosaur Project — The Story of the Greatest Dinosaur Expedition Ever Mounted, Edmonton: Ex Terra Foundation. Toronto: Macfarlane Walter & Ross. 61 pp
- Brusatte, Stephen L.; Benson, Roger B. J.; Currie, Philip J.; Zhao, Xijin (2010). "The skull of Monolophosaurus jiangi (Dinosauria: Theropoda) and its implications for early theropod phylogeny and evolution" (PDF). Zoological Journal of the Linnean Society. 158 (3): 573–607. doi:10.1111/j.1096-3642.2009.00563.x. hdl:20.500.11820/9db48bd7-0312-40ee-96a5-dccc897fd0cd. S2CID 83418757.
- Zhao, Xijin; Benson, Roger B. J.; Brusatte, Stephen L.; Currie, Philip J. (2010). "The postcranial skeleton of Monolophosaurus jiangi (Dinosauria: Theropoda) from the Middle Jurassic of Xinjiang, China, and a review of Middle Jurassic Chinese theropods". Geological Magazine. 147 (1): 13–27. Bibcode:2010GeoM..147...13X. doi:10.1017/S0016756809990240. hdl:20.500.11820/9593b624-85da-4359-9b7d-53dddcc4ecd9. S2CID 53374224.
- Carr, T. (2006). "Is Guanlong a tyrannosauroid or a subadult Monolophosaurus?". Journal of Vertebrate Paleontology. 26: 48A. doi:10.1080/02724634.2006.10010069.
- Paul, G.S., 2010, The Princeton Field Guide to Dinosaurs, Princeton University Press p. 93-94
- Smith ND, Makovicky PJ, Hammer WR, Currie PJ. 2007. Osteology of Cryolophosaurus ellioti (Dinosauria:Theropoda) from the Early Jurassic of Antarctica and implications for early theropod evolution. Zoological Journal of the Linnean Society 151: 377–421.
- Benson, Roger B. J. (2010). "A description of Megalosaurus bucklandii (Dinosauria: Theropoda) from the Bathonian of the UK and the relationships of Middle Jurassic theropods". Zoological Journal of the Linnean Society. 158 (4): 882–935. doi:10.1111/j.1096-3642.2009.00569.x.
- Benson, 2008. A new theropod phylogeny focussing on basal tetanurans, and its implications for European 'megalosaurs' and Middle Jurassic dinosaur endemism. Journal of Vertebrate Paleontology. 51A.
- Carrano, M. T.; Benson, R. B. J.; Sampson, S. D. (2012). "The phylogeny of Tetanurae (Dinosauria: Theropoda)". Journal of Systematic Palaeontology. 10 (2): 211–300. doi:10.1080/14772019.2011.630927. S2CID 85354215.
- Rauhut, Oliver W. M.; Pol, Diego (December 2019). "Probable basal allosauroid from the early Middle Jurassic Cañadón Asfalto Formation of Argentina highlights phylogenetic uncertainty in tetanuran theropod dinosaurs". Scientific Reports. 9 (1): 18826. Bibcode:2019NatSR...918826R. doi:10.1038/s41598-019-53672-7. ISSN 2045-2322. PMC 6906444. PMID 31827108.
- Schade, Marco; Rauhut, Oliver; Foth, Christian; Moleman, Olof; Evers, Serjoscha (2023). "A reappraisal of the cranial and mandibular osteology of the spinosaurid Irritator challengeri (Dinosauria: Theropoda)". Palaeontologia Electronica. doi:10.26879/1242.
- Molnar, R. E., 2001, Theropod paleopathology: a literature survey: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 337-363.
Further reading
- Dong Zhiming (1992). Dinosaurian Faunas of China. Beijing: China Ocean Press. ISBN 3-540-52084-8.
External links
- Carr, Thomas (2006). "Is Guanlong a tyrannosauroid or a subadult Monolophosaurus?" (PDF). Journal of Vertebrate Paleontology. 26 (3 Supplement): 48A. doi:10.1080/02724634.2006.10010069.
- Monolophosaurus in the Dino Directory, The Natural History Museum, London.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||

.jpg.webp)