Torvosaurus
Torvosaurus (/ˌtɔːrvoʊˈsɔːrəs/) is a genus of large megalosaurine theropod dinosaur that lived approximately 165 to 148 million years ago during the Callovian to Tithonian ages of the late Middle and Late Jurassic period in what is now Colorado, Portugal, Germany, and possibly England, Spain, Tanzania, and Uruguay. It contains two currently recognized species, Torvosaurus tanneri and Torvosaurus gurneyi, plus a third unnamed species from Germany.[1]
| Torvosaurus | |
|---|---|
 | |
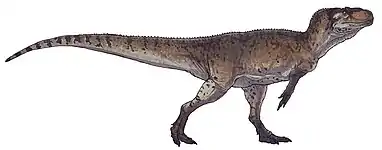
| Mounted T. tanneri skeletal reconstruction, Museum of Ancient Life | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Family: | †Megalosauridae |
| Subfamily: | †Megalosaurinae |
| Genus: | †Torvosaurus Galton & Jensen, 1979 |
| Type species | |
| †Torvosaurus tanneri Galton & Jensen, 1979 | |
| Other species | |
| |
| Synonyms | |
| |
In 1979, the type species Torvosaurus tanneri was named. It is a large, heavily built, bipedal carnivore that could grow to a length of about 9 meters (30 ft) and weigh approximately 2 metric tons (2.2 short tons). T. tanneri was among the largest terrestrial carnivores of its time, alongside Epanterias and Saurophaganax (which could both be synonymous with Allosaurus). Specimens of Torvosaurus gurneyi were measured up to 10–11 meters (33–36 ft) in length and 4–5 metric tons (4.4–5.5 short tons) in body mass,[2][3] suggesting that it was much larger than T. tanneri and was the largest terrestrial carnivore in Europe during the late Jurassic. Based on bone morphology, Torvosaurus is thought to have had very powerful short arms.
Discovery

Fossilized remains of Torvosaurus have been found in North America, Portugal, Germany, and possibly in England, Spain, Tanzania, and Uruguay. The first discovered remains referable to Torvosaurus were discovered in 1899 by Elmer Riggs in the "Freeze-out Hills" of southeastern Wyoming, 18 kilometers (11 mi) northwest of Medicine Bow. The material consisted of part of the left foot and right hand and they were taken to the Field Museum of Natural History in Chicago, where they were stored until being re-discovered around 2010. The specimen was assigned to Torvosaurus tanneri after being described in 2014.[4]
More remains of a large theropod that is now believed to have been Torvosaurus were discovered in the Tendaguru Formation of Tanzania and was named "Megalosaurus" ingens by Werner Janensch in 1920, based on the specimen MB R 1050, a 12-centimeter (4.7 in) long tooth from German East Africa (now Tanzania).[5] It was eventually reclassified as a probable member of Carcharodontosauridae before being reclassified as a probable member of the Torvosaurus genus in 2020.[6] Although it was only referred to as Torvosaurus sp., one commentator has noted it could potentially be called Torvosaurus ingens.[7][8] Soto et al. described teeth of a member of the genus Torvosaurus from the Tacuarembó Formation of Uruguay. The authors noted that some of the specimens of “Megalosaurus” ingens figured by Werner Janensch share the features of the Uruguayan material and stated that the materials from Tanzania and Uruguay may represent the same taxon, due to geographical proximity, but ultimately concluded that, based on only teeth, they do not share any derived characteristics to distinguish them from the described species of the genus, T. tanneri and T. gurneyi.[6][9] However, Rauhut et al. consider the teeth to be undiagnostic, being coherent in size and shape with a variety of other theropods (including carcharodontosaurids), thus considering their attribution to the genus to be problematic.[1]
In 1971, Vivian Jones of Delta, Colorado, in the Calico Gulch Quarry in Moffat County, discovered a single gigantic thumb claw of a theropod. This was shown to James Alvin Jensen, a collector who was working for Brigham Young University. In an effort to discover comparable fossils, Vivian's husband Daniel Eddie Jones directed Jensen to the Dry Mesa Quarry, where abundant gigantic theropod bones, together with Supersaurus remains, proved present in rocks of the Morrison Formation. From 1972 onward, the site was excavated by Jensen and Kenneth Stadtman. The type species Torvosaurus tanneri was named and described in 1979 by Peter Malcolm Galton and Jensen.[10] The genus name Torvosaurus derives from the Latin word torvus, meaning "savage", and the Greek word sauros (σαυρος), meaning "lizard".[11] The specific name tanneri comes from the, first counselor in the First Presidency of the Church of Jesus Christ of Latter-day Saints, Nathan Eldon Tanner.
In 1985, Jensen could report a considerable amount of additional material, among it being the first skull elements.[12] The fossils from Colorado were further described by Brooks Britt in 1991.[13] The holotype, BYU 2002, originally consisted of upper and lower arm bones. The paratypes included some back bones, hip bones, and hand bones.[10] When the material described in 1985 is added, the main missing elements are the shoulder girdle and the thighbone.[13] The original thumb claw, specimen BYUVP 2020, was only provisionally referred, as it had been found in a site 195 kilometers (121 mi) away from the Dry Mesa Quarry.[10] The holotype and paratypes represented at least three individuals, these being two adults and a juvenile.[13] In 1991, Britt concluded that there was no proof that the front limbs of the holotype were associated and chose the left humerus as the lectotype.[13] Several single bones and teeth found in other American sites have been referred to Torvosaurus.[13]

In 1992, fossils of a large theropod found at Como Bluff in Wyoming contained skull, shoulder girdle, pelvic, and rib elements. They were named by Robert T. Bakker et al. as the species Edmarka rex. Bakker et al were impressed with the size of Edmarka, noting that it "would rival T. rex in total length," and viewing this approximate size as "a natural ceiling for dinosaurian meat-eaters."[14] This was often considered a junior synonym of Torvosaurus,[15] but a detailed analysis has not been carried out yet.[16] The same site has rendered comparable remains for which the nomen nudum Brontoraptor has been used.[17][18] Most researchers now regard both specimens as belonging to Torvosaurus tanneri.[2] However, Edmarka rex and Brontoraptor require reclassification to determine whether or not they actually belong to T. tanneri, as all the specimens described prior to their discovery indicate that they reached their adult size and both incomplete specimens lack detailed osteological descriptions.[19]
In 2012, a still undescribed, 55% complete specimen was discovered in Colorado in the Skull Creek Quarry, which is an exposure of the Morrison Formation. The specimen, nicknamed "Elvis", included the pelvic, spine, and hind limb bones, a complete, associated backbone, as well as cranial elements. It is the most complete specimen of Torvosaurus found to date.[20] A mounted skeleton of the specimen, with missing parts reconstructed with casts from other Torvosaurus specimens, is currently on display in the Museum of Natural History & Science in Cincinnati.[20][21]
In 2000, material from Portugal was referred to a Torvosaurus sp. by Octávio Mateus and Miguel Telles Antunes.[22] In 2006, fossils from the Portuguese Lourinhã Formation were referred to Torvosaurus tanneri.[3] In 2012, however, Matthew Carrano et al. concluded that this material could not be more precisely determined than a Torvosaurus sp.[23] In 2013 and 2014, eggs with and without embryos were reported from Portugal and referred to Torvosaurus.[24][25] The species from Portugal was named T. gurneyi in honor of James Gurney in 2014, the creator of the Dinotopia series of books. It is the largest named theropod known from Europe, although an isolated anterior caudal vertebra from the Vega Formation in Spain, which may belong to Torvosaurus or a closely related taxon, is about 15% larger than the one found on T. gurneyi.[2][26] Furthermore, remains of a large spinosaurid from the Vectis Formation in the Isle of Wight include vertebrae of size comparable to those of Spinosaurus, indicating an animal exceeding 10 m in length, making it the largest reported European theropod.[27][28] It was the morphological distinctiveness of the holotype maxilla ML1100 that led to the naming of the Portuguese species.[2] In 2017, a set of Portuguese cranial material assigned to Torvosaurus was described, including a specimen interpreted as belonging to the same individual as the holotype of Torvosaurus gurneyi.[29]
In 2020, Soto et al. described FC-DPV 2971, a tooth from Uruguay, as belonging to a new unnamed species of Torvosaurus.[6] They also assigned Megalosaurus/Ceratosaurus ingens (specimen MB R 1050) from Tanzania to Torvosaurus.[6] Also in 2020, a fragmentary maxilla referable to Torvosaurus was described from the middle Callovian Ornatenton Formation of Germany. This is the oldest record of the genus and suggests that megalosaurines originated in Europe, or at least that Europe was a biogeographical turntable for them from the Middle to the early Late Jurassic. Other possible Torvosaurus instances in Europe include fragmentary remains from the Kimmeridge Clay of England that possibly belong to the genus.[1] These consist of a tibia (OUMNH J.29886) and a maxilla fragment that were collected separately from each other.[23]
Description

Torvosaurus was a very large and robust predatory dinosaur, with an estimated maximum body length of 10–11 meters (33–36 ft) and mass of 4–5 metric tons (4.4–5.5 short tons) for the Portuguese species T. gurneyi,[2][3] making T. gurneyi among the largest terrestrial carnivores of the Jurassic. Claims have been made indicating even larger sizes for the American species T. tanneri up to 12 m (39 ft) in length and more than 4 t (4.4 short tons) based on incomplete remains of Edmarka rex and Brontoraptor.[30][16][31] However, these claims regarding T. tanneri are likely erroneous, as the discovered remains of T. tanneri indicate that they reached adult size at 9 meters (30 ft) in length and approximately 2 metric tons (2.2 short tons) in body mass.[13][32][33][34] Furthermore, Edmarka rex and Brontoraptor lack detailed analyses to verify whether or not they actually belong to T. tanneri.[19][16]
Among the differentiating features originally recognized between T. gurneyi and T. tanneri are the number of teeth, alongside the size and shape of the mouth. While the upper jaw of T. tanneri has more than 11 teeth, that of T. gurneyi has less.[2] However, later examination of a new right maxilla, probably belonging to the same individual as the holotype of T. gurneyi, has determined that, while the two species can be distinguished based on the morphology of the maxillary medial wall and interdental plates, the supposedly lower number of maxillary teeth in the Portuguese form may be an artifact of preservation, since it is not possible to know the exact number of teeth in the complete maxilla at the moment.[29]
The material from Germany is further distinguished by the other two species by a temporal difference of c. 10 Ma and a few morphological differences which indicate that it was the third species outside a sister taxon relationship between T. tanneri and T. gurneyi. The material is only 10% smaller than the maxilla of T. tanneri, although the ontogenetic stage of the specimen is unknown. This indicates that derived megalosaurines were already among the largest terrestrial predators of the late Middle Jurassic, with only a moderate increase in size in the genus by the Late Jurassic.[1]

Torvosaurus had an elongated, narrow snout, with a kink in its profile just above the large nostrils. The frontmost snout bone, the premaxilla, bore three rather flat teeth oriented somewhat outwards with the front edge of the teeth crown overlapping the outer side of the rear edge of the preceding crown. The maxilla was tall and bore at least eleven rather long teeth. The antorbital fenestra was relatively short. The lacrimal bone had a distinctive lacrimal horn on top. Its lower end was broad in side view. The eye socket was tall with a pointed lower end. The jugal was long and transversely thin. The lower front side of the quadrate bone was hollowed out by a tear-shaped depression, the contact surface with the quadratojugal. Both the neck vertebrae and the front dorsal vertebrae had relatively flexible ball-in-socket joints. The balls on the front side of the vertebral centra had a wide rim, a condition by Britt likened to a Derby hat. The tail base was stiffened in the vertical plane by high and in side view wide neural spines. The whole of the arm was very strong, but somewhat short. Whether the thumb claw was especially enlarged is uncertain. In the pelvis, the ilium resembled that of Megalosaurus and had a tall, short, front blade and a longer pointed rear blade. The pelvis as a whole was massively built, with the bone skirts between the pubic bones and the ischia contacting each other and forming a vaulted closed underside.[13]
Systematics and classification

When first described in 1979 by Galton and Jensen,[10] Torvosaurus was classified as a megalosaurid, which is the current consensus.[15] It was later assigned to Carnosauria by Ralph Molnar et al. in 1990,[35] then to a basal position in Spinosauroidea by Oliver Walter Mischa Rauhut in 2003,[36] and to a very basal position in Tetanurae by Thomas Holtz in 1994.[37] All these assignments are not supported by present phylogenetic analysis.[15] In 1985, Jensen assigned Torvosaurus to a family of its own, Torvosauridae.[12] Despite support for this concept by Paul Sereno[38] and Mateus,[3] it seems redundant because Torvosaurus is closely related to, and perhaps the sister species of, the earlier Megalosaurus within Megalosaurinae.[15] However, Torvosauridae may be used as an alternative name for Megalosauridae if Megalosaurus is considered an indeterminable nomen dubium.[39] Though a close relative of Megalosaurus, Torvosaurus is seemingly more advanced, or apomorphic. Torvosaurus's larger clade, Megalosauridae, is most commonly held as a basal branch of Tetanurae, considered to be less derived than carnosaurs or coelurosaurs and likely related to the spinosaurids.[15]
The following is a cladogram based on the phylogenetic analysis conducted by Carrano, Benson, and Sampson (2012) showing the relationships of Torvosaurus:[15]
| Megalosauroidea |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Distinguishing anatomical features

According to Carrano et al. (2012), Torvosaurus can be distinguished based on the following characteristics:[40]
- The presence of a very shallow maxillary fossa (it lacks a fenestra maxillaris piercing the bone wall)
- The presence of fused interdental plates
- The pneumatic fossae in the posterior dorsal and the anterior caudal vertebrae centra being expanded to form enlarged, deep openings
- The puboischiadic plate being highly ossified (the paired bony plates of both sides connect and close off the entire underside of the pelvis, a very basal trait that Galton & Jensen saw as an indication that Theropoda was polyphyletic, with Carnosauria having independently evolved from carnivorous Prosauropoda)[10]
- A distal expansion of the ischium shaft with a prominent lateral midline crest and an ovalur outline when examined in lateral view
- The cervical vertebrae are opisthocoelous with a pronounced flat rim around the anterior ball (according to Rauhut, 2000)
- A transverse fenestra is situated in the neural arch of the dorsal vertebrae in front of the hyposphene (according to Rauhut, 2000)[41]
Paleobiology

The study of fossilized embryos of Torvosaurus provides researchers with information about the transformation of the embryo over time, the different developmental pathways present in dinosaur lineages, dinosaur reproductive behavior, and dinosaur parental care.[42][43][44] In 2013, Araújo et al. announced the discovery of specimen ML1188, a clutch of crushed dinosaur eggs and embryonic material attributed to Torvosaurus.[45] This discovery further supports the hypothesis that large theropods were oviparous, meaning that they laid eggs and hence that embryonic development occurred outside the body of female dinosaurs. This discovery was made in 2005 by the Dutch amateur fossil-hunter Aart Walen at the Lourinhã Formation in Western Portugal in fluvial overbank sediments that are considered to be from the Tithonian age of the Jurassic, approximately 152 to 145 million years ago. This discovery is paleontologically significant for a number of reasons: (a) these are the most primitive dinosaur embryos known; (b) these are the only basal theropod embryos known; (c) fossilized eggs and embryos are rarely found together; (d) it represents the first evidence of a one-layered eggshell for theropod dinosaurs; and (e) it allows researchers to link a new eggshell morphology to the osteology of a particular group of theropod dinosaurs.[24] The specimen is housed at the Museu da Lourinhã in Portugal. As the eggs were abandoned due to unknown circumstances, it is not known if Torvosaurus provided parental care to its eggs and young or abandoned them shortly after laying.[46] However, the eggshells are highly porous, allowing efficient gaseous exchange between the external and internal media, thus indicating the eggs were buried for incubation within substrate in a manner similar to modern seaturtles. This is also corroborated by the undisturbed taphonomic setting and low-energy geological context.[24]
All documented Torvosaurus specimens from the Morrison Formation are from similarly sized, likely adult individuals and the lack of immature individuals may be explained by many factors, none of which are mutually exclusive. For one thing, the formation is known to preserve large vertebrates better than smaller ones. Immature individuals may also have occupied a different ecological niche from adults in habitats where their remains were likely to preserve as fossils and they may have been the prey of choice of larger predators as well. Torvosaurus may also have experienced Type B1 population survivorship, as has been found in other dinosaurs, with mortality increasing after sexual maturity was achieved, leading to an abundance of mature individuals in the fossil record. A final possibility is that immature Torvosaurus remains could be misidentified due to having different proportions compared to the very large and robust adults.[4]
Paleoecology

Provenance and occurrence
The type specimen of Torvosaurus tanneri, BYU 2002, was recovered in the Dry Mesa Quarry of the Brushy Basin Member of the Morrison Formation in Montrose County, Colorado. The specimen was collected by James A. Jensen and Kenneth Stadtman in 1972 in medium-grained, coarse sandstone that was deposited during the Tithonian and Kimmeridgian ages of the Jurassic, approximately 153 to 148 million years ago.[47] This specimen is housed in the collection of Brigham Young University in Provo, Utah.
Fauna and habitat in North America
Studies suggest that the paleoenvironment of this section of the Morrison Formation included rivers that flowed from the west into a basin that contained a giant, saline alkaline lake and there were extensive wetlands in the vicinity. The Dry Mesa Dinosaur Quarry of western Colorado yields one of the most diverse Upper Jurassic vertebrate assemblages in the world.[48] The Dry Mesa Quarry has produced the remains of the sauropods Apatosaurus, Brachiosaurus, Diplodocus, Barosaurus, Supersaurus, and Camarasaurus, the iguanodonts Camptosaurus and Dryosaurus, and the theropods Allosaurus, Tanycolagreus, Koparion, Stokesosaurus, Ceratosaurus, and Ornitholestes, as well as Othnielosaurus, Gargoyleosaurus, and Stegosaurus.[49]
The flora of the period has been revealed by fossils of green algae, fungi, mosses, horsetails, ferns, cycads, ginkgoes, and several families of conifers. Other animal fossils discovered include bivalves, snails, ray-finned fishes, frogs, salamanders, amphibians, turtles, sphenodonts, lizards, terrestrial (like Hoplosuchus) and aquatic crocodylomorphs, cotylosaurs, several species of pterosaurs, like Harpactognathus, and early mammals, such as multituberculates, symmetrodonts, and triconodonts.[49]
Fauna and habitat in Europe
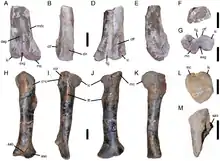
The Ornatenton Formation is a Callovian aged shallow marine deposit, within the formation Torvosaurus was sympatric with the closely related and also large Wiehenvenator. The Lourinhã Formation is Kimmeridgian-Tithonian in age. The environment is coastal and therefore has a strong marine influence. Its flora and fauna are very similar to that of the Morrison. Torvosaurus appears to be the top predator here. It lived alongside the European species of Allosaurus (A. europaeus), Ceratosaurus, Stegosaurus, and presumably Camptosaurus. The theropod Lourinhanosaurus also stalked the area. Lusotitan was the largest sauropod in the region, while the diplodocids Dinheirosaurus and Lourinhasaurus were also present. Dacentrurus and Miragaia were both stegosaurs, while Dracopelta was an ankylosaurian. Draconyx was an iguanodontid related to Camptosaurus. Due to the marine nature of the Lourinhã Formation, sharks, plesiochelyid turtles, and teleosaurid crocodyliforms are also present.[50]
Fauna and habitat in Africa
The small-scale trough and ripple cross-bedded fine-grained sandstone at the base of the Upper Dinosaur Member of the Tendaguru Formation, of which possible Torvosaurus material is known from, is interpreted as tidal flat deposits. Stagnant water bodies, such as small lakes and ponds, were present and a freshwater depositional environment close to the sea was also probably present.
The possible unnamed Torvosaurus species from the Tendaguru Formation would have shared its habitat with many species of sauropods, such as Australodocus, Dicraeosaurus, Giraffatitan, Janenschia, Tornieria, Wamweracaudia, three unnamed species of diplodocine sauropods, an unnamed species of flagellicaudatan, and "The Archbishop". The theropods it coexisted with were Allosaurus tendagurensis, Ceratosaurus roechlingi, Elaphrosaurus, Labrosaurus stechowi, Ostafrikasaurus, Veterupristisaurus (of which it would have possibly competed with to be top predator), a possible abelisauroid, and an indeterminate megalosauroid, while the ornithischians it coexisted with were Dysalotosaurus and Kentrosaurus. As far as pterosaurs are concerned, it coexisted with Tendaguripterus, an indeterminate archaeopterodactyloid, an indeterminate azhdarchid, two indeterminate dsungaripteroids, an indeterminate rhamphorynchoid, and an indeterminate pterosaur of unknown classification. Due to the coastal environment of the Tendaguru Formation, crocodyliformes, such as Bernissartia, amphibians, including an unnamed lissamphibian, and sharks are also present.[51]
Fauna and habitat in South America
Within South America, possible Torvosaurus remains are only present within the Tacuarembó Formation of Uruguay. Because the formation was laid down in fluvial to lacustrine sandstones, siltstones, and mudstones, this indicates that the environment of the formation would have been dominated by rivers, streams, and lakes.
If present, Torvosaurus would probably have been the apex predator in the Tacuarembó Formation, although it may have been rivalled by cf. Ceratosaurus.[52] It shared its habitat with two unnamed theropods, an unnamed coelurosaur, an unnamed sauropod known solely from footprints, an unnamed ornithopod also known from footprints, an indeterminate mesoeucrocodylian, an indeterminate turtle species that lived alongside the named turtle species Tacuarembemys kusterae, and the possible pholidosaur Meridiosaurus vallisparadisi. Fish, such as Arganodus tiguidiensis, Asiatoceratodus cf. tiguidensis, Neoceratodus africanus, and Priohybodus arambourgi, and bivalves, such as Diplodon, are also present.[53][54]
Coexistence with other large carnivores
Torvosaurus coexisted with other large theropods such as Allosaurus, Ceratosaurus, and Saurophaganax in the United States, Allosaurus, Ceratosaurus, and Lourinhanosaurus in Portugal, possibly Veterupristisaurus in Tanzania, and possibly cf. Ceratosaurus (?) in Uruguay. The three, possibly four, species appear to have had different ecological niches, based on anatomy and the location of fossils. Torvosaurus and Ceratosaurus may have preferred to be active around waterways and had lower, more sinuous, bodies that would have given them an advantage in forest and underbrush terrains, whereas Allosaurus had shorter bodies, longer legs, were faster and less maneuverable, and seem to have preferred dry floodplains.[55] Also, Rauhut et al. (2016) proposed that allosaurids and megalosaurids would have had different environmental preferences, the former being more common in inland areas, while the latter being dominant in marine and coastal environments.[56]
On the other hand, the majority of Torvosaurus remains from the Morrison Formation have been found in localities preserving multiple taxa, including Allosaurus, with Torvosaurus itself being a minor component of the bonebeds. This pattern has been interpreted as indicative of Torvosaurus sharing habitats with other predators, most notably Allosaurus, but at much lower abundances.[4] The three may also have had different dietary preferences, with Allosaurus being more suited for bone slicing (thanks to its short and stout serrated teeth, deep and narrow skull, and powerful dorsoventral movement capacity of the neck), while Ceratosaurus, with its long and blade-like teeth and relatively straight neck, would have probably been incapable of doing so, instead concentrating on the deepest organs of a carcass. While probably capable of some bone consumption, Torvosaurus, with its large skull and teeth and large, powerful, and lithe body may have been specialized in opening up and dismembering exceptionally large sauropod carcasses. This would have allowed smaller theropods, like Allosaurus, better access in a possible commensalism relationship.[57]
Allosaurus was itself a potential food item to other carnivores, as illustrated by an Allosaurus pubic foot marked by the teeth of another theropod, probably Ceratosaurus or Torvosaurus. The location of the bone in the body (along the bottom margin of the torso and partially shielded by the legs) and the fact that it was among the most massive in the skeleton indicates that the Allosaurus was being scavenged.[58]

Bite marks on Allosaurus and Mymoorapelta remains were found among other bones with feeding traces in the Upper Jurassic Mygatt-Moore Quarry. Unlike the others, these have left striations that, when measured to determine denticle width, produced tooth and body size extrapolations greater than any known specimen of Allosaurus or Ceratosaurus, the two large predators known for osteological remains from the quarry. The extrapolations are instead coherent, either with an unusually large specimen of Allosaurus or a separate large taxon, like Torvosaurus or Saurophaganax, both of which are not known from the quarry. The result either increases the known diversity of the site based on ichnological evidence alone or represents powerful evidence of cannibalism in Allosaurus. Based on the position and nutrient value associated with the various skeletal elements with bite marks, it is predicted that while Mymoorapelta was either predated upon or scavenged shortly after death, Allosaurus was scavenged some time after death.[59]
References
- Oliver W. M. Rauhut; Achim H. Schwermann; Tom R. Hübner; Klaus-Peter Lanser (2020). "The oldest record of the genus Torvosaurus (Theropoda: Megalosauridae) from the Callovian Ornatenton Formation of north-western Germany" (PDF). Geologie und Paläontologie in Westfalen. 93: 1–13.
- Hendrickx, C.; Mateus, O. (2014). Evans, Alistair Robert (ed.). "Torvosaurus gurneyi n. sp., the Largest Terrestrial Predator from Europe, and a Proposed Terminology of the Maxilla Anatomy in Nonavian Theropods". PLOS ONE. 9 (3): e88905. Bibcode:2014PLoSO...988905H. doi:10.1371/journal.pone.0088905. PMC 3943790. PMID 24598585.
- Mateus, Octávio; Walen, Aart; Antunes, Miguel Telles (2006). "The large theropod fauna of the Lourinha Formation (Portugal) and its similarity to that of the Morrison Formation, with a description of a new species of Allosaurus". In Foster, John R.; Lucas, Spencer G. (eds.). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 123–129.
- Hanson, Michael; Makovicky, Peter J. "A new specimen of Torvosaurus tanneri originally collected by Elmer Riggs". Historical Biology, volume 26, issue 6 (2014). Pages 775-784. https://www.tandfonline.com/doi/abs/10.1080/08912963.2013.853056?scroll=top&needAccess=true&journalCode=ghbi20.
- Janensch, W. (1920). "Ueber Elaphrosaurus bambergi und die Megalosaurier aus den Tendaguru Schichten Deutsch-Ostafrikas". Sitzungsberichte der Gesellschaft Naturforschender Freunde zu Berlin: 225–235.
- Soto, Matías; Toriño, Pablo; Perea, Daniel (2020). "A large sized megalosaurid (Theropoda, Tetanurae) from the late Jurassic of Uruguay and Tanzania". Journal of South American Earth Sciences. 98: 102458. Bibcode:2020JSAES..9802458S. doi:10.1016/j.jsames.2019.102458. S2CID 213672502.
- "Torvosaurus: The Grizzly Bear of the Jurassic". Dinosaurs and Barbarians. August 8, 2015. Retrieved March 22, 2021.
- Soto, Matias (September 30, 2019). "Megalosaurid tooth from Uruguay". Mendeley Data. 1. doi:10.17632/6kntxfp449.1.
- Tacuarembó Formation at Fossilworks.org
- P. M. Galton and J. A. Jensen. 1979. A new large theropod dinosaur from the Upper Jurassic of Colorado. Brigham Young University Geology Studies 26(1):1–12
- Liddell, Henry George and Robert Scott (1980). A Greek-English Lexicon (Abridged ed.). United Kingdom: Oxford University Press. ISBN 0-19-910207-4.
- Jensen, J.A., 1985, "Uncompahgre dinosaur fauna: A preliminary report", Great Basin Naturalist, 45: 710–720
- Britt, B., 1991, "Theropods of Dry Mesa Quarry (Morrison Formation, Late Jurassic), Colorado, with emphasis on the osteology of Torvosaurus tanneri", Brigham Young University Geology Studies 37: 1–72
- Bakker, R.T., Siegwarth, J., Kralis, D. & Filla, J., 1992, "Edmarka rex, a new, gigantic theropod dinosaur from the middle Morrison Formation, Late Jurassic of the Como Bluff outcrop region", Hunteria, 2(9): 1–24
- Carrano, M. T.; Benson, R. B. J.; Sampson, S. D. (2012). "The phylogeny of Tetanurae (Dinosauria: Theropoda)". Journal of Systematic Palaeontology 10 (2): 211–300. doi:10.1080/14772019.2011.630927
- Molina-Pérez, R.; Larramendi, A. (2019). Encyclopedia of Dinosaurs: the Theropods. London: The Natural History Museum. ISBN 9780565094973.
- Redman, P.D., 1995, Paleo Horizons, Winter Issue
- Siegwarth, J., Linbeck, R., Bakker, R. and Southwell, B. (1996). Giant carnivorous dinosaurs of the family Megalosauridae. Hunteria 3:1–77.
- Dalman, S. G. (2014). "New data on small theropod dinosaurs from the Upper Jurassic Morrison Formation of Como Bluff, Wyoming, USA" (PDF). Volumina Jurassica. 7 (2): 181–196.
- "Torvosaurus – King of the Real Jurassic World Unearthed". FossilEra. Retrieved March 21, 2021.
- "Museum of Natural History & Science (MNHS)". Cincinnati Museum Center. Retrieved March 21, 2021.
- Mateus, O., & Antunes, M. T. (2000). Torvosaurus sp.(Dinosauria: Theropoda) in the late Jurassic of Portugal. In I Congresso Ibérico de Paleontologia/XVI Jornadas de la Sociedad Española de Paleontología (pp. 115–117)
- Carrano, Matthew T.; Benson, Roger B. J.; Sampson, Scott D. (June 2012). "The phylogeny of Tetanurae (Dinosauria: Theropoda)". Journal of Systematic Palaeontology. 10 (2): 211–300. doi:10.1080/14772019.2011.630927. ISSN 1477-2019. S2CID 85354215.
- Arau´jo, Ricardo; Castanhinha, Rui; Martins, Rui M. S.; Mateus, Octa´vio; Hendrickx, Christophe; Beckmann, F.; Schell, N.; Alves, L. C. (2013). "Filling the gaps of dinosaur eggshell phylogeny: Late Jurassic Theropod clutch with embryos from Portugal" (PDF). Scientific Reports. 1924: 8. Bibcode:2013NatSR...3E1924A. doi:10.1038/srep01924. PMC 3667465. PMID 23722524. S2CID 7766400.
- Ribeiro, Vasco; Mateus, Octávio; Holwerda, Femke; Araújo, Ricardo; Castanhinha, Rui (March 4, 2014). "Two new theropod egg sites from the Late Jurassic Lourinhã Formation, Portugal". Historical Biology. 26 (2): 206–217. doi:10.1080/08912963.2013.807254. ISSN 0891-2963. S2CID 55315504.
- Rauhut, Oliver W.M.; Piñuela, Laura; Castanera, Diego; García-Ramos, José-Carlos; Sánchez Cela, Irene (July 5, 2018). "The largest European theropod dinosaurs: remains of a gigantic megalosaurid and giant theropod tracks from the Kimmeridgian of Asturias, Spain". PeerJ. 6: e4963. doi:10.7717/peerj.4963. ISSN 2167-8359. PMC 6035862. PMID 30002951.
- Barker, Chris T.; Lockwood, Jeremy A. F.; Naish, Darren; Brown, Sophie; Hart, Amy; Tulloch, Ethan; Gostling, Neil J. (June 9, 2022). "A European giant: a large spinosaurid (Dinosauria: Theropoda) from the Vectis Formation (Wealden Group, Early Cretaceous), UK". PeerJ. 10: e13543. doi:10.7717/peerj.13543. ISSN 2167-8359. PMC 9188774. PMID 35702254.
- "A Giant Spinosaurid Dinosaur from the Cretaceous of the Isle of Wight". Tetrapod Zoology. June 9, 2022. Retrieved September 11, 2022.
- Malafaia, E.; Mocho, P.; Escaso, F.; Ortega, F. (March 2017). "New data on the anatomy of Torvosaurus and other remains of megalosauroid (Dinosauria, Theropoda) from the Upper Jurassic of Portugal". Journal of Iberian Geology. 43 (1): 33–59. doi:10.1007/s41513-017-0003-9. ISSN 1698-6180. S2CID 132198140.
- Holtz, Thomas R. (2012). "Dinosaurs: The Most Complete, Up-to-date Encyclopedia for Dinosaur Lovers of All Ages" (PDF).
- Siegwarth, J., Linbeck, R., Bakker, R. and Southwell, B., 1996, "Giant carnivorous dinosaurs of the family Megalosauridae", Hunteria 3: 1–77
- Paul, G. S. (2016). The Princeton Field Guide to Dinosaurs (2nd ed.). Princeton, New Jersey: Princeton University Press. p. 93. ISBN 9780691167664.
- Farlow, J. O.; Coroian, D.; Currie, P.J.; Foster, J.R.; Mallon, J.C.; Therrien, F. (2022). ""Dragons" on the landscape: Modeling the abundance of large carnivorous dinosaurs of the Upper Jurassic Morrison Formation (USA) and the Upper Cretaceous Dinosaur Park Formation (Canada)". The Anatomical Record. 306 (7): 1669–1696. doi:10.1002/ar.25024. PMID 35815600.
- Paul, Gregory S. (1988). Predatory Dinosaurs of the World. Simon & Schuster. p. 282. ISBN 0-671-61946-2.
- R. E. Molnar, S. M. Kurzanov, and Z. Dong. 1990. "Carnosauria". In: D. B. Weishampel, H. Osmólska, and P. Dodson (eds.), The Dinosauria. University of California Press, Berkeley
- O. W. M. Rauhut. 2003. The interrelationships and evolution of basal theropod dinosaurs. Special Papers in Palaeontology 69:1–213
- T.R. Holtz, 1994, "The phylogenetic position of the Tyrannosauridae: implications for theropod systematics", Journal of Paleontology 68(5): 1100–1117
- P.C. Sereno, J.A. Wilson, H.C.E. Larsson, D.B. Dutheil, and H.-D. Sues, 1994, "Early Cretaceous dinosaurs from the Sahara", Science 266(5183): 267–271
- P.C. Sereno, 1998, "A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria", Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 210(1): 41–83
- Carrano, Benson and Sampson, 2012. The phylogeny of Tetanurae (Dinosauria: Theropoda). Journal of Systematic Palaeontology. 10(2), 211–300
- Rauhut, 2000. The interrelationships and evolution of basal theropods (Dinosauria, Saurischia). Ph.D. dissertation, Univ. Bristol [U.K.], 1–440.
- Long, J. A. & McNamara, K. J. Heterochrony: The key to dinosaur evolution. in The Dinofest International (Wolberg, D.L., Stump, E. & Rosenberg, G. D., eds.) 113–123 ( Acad. Nat. Sci. Phil. , 1997).
- Balanoff, A. M., Norell, M. A., Grellet-Tinner, G. & Lewin, M. R. Digital preparation of a probable neoceratopsian preserved within an egg, with comments on microstructural anatomy of ornithischian eggshells. Naturwissenschaften 95 , 493–500 (2008).
- Varricchio, D. J. et al., 2008. Avian paternal care had dinosaur origin. Science 322 , 1826– 1828.
- Araújo, Ricardo; Castanhinha, Rui; Martins, Rui M. S.; Mateus, Octávio; Hendrickx, Christophe; Beckmann, F.; Schell, N.; Alves, L. C. (May 30, 2013). "Filling the gaps of dinosaur eggshell phylogeny: Late Jurassic Theropod clutch with embryos from Portugal". Scientific Reports. 3 (1): 1924. Bibcode:2013NatSR...3E1924A. doi:10.1038/srep01924. ISSN 2045-2322. PMC 3667465. PMID 23722524.
- "A rare find: Abandoned dinosaur nests with eggshells". NBC News. August 5, 2013.
- Jensen, J. A. and Ostrom, J. H. (1977). A second Jurassic pterosaur from North America. Journal of Paleontology 51(4):867–870
- Richmond, D.R. and Morris, T.H., 1999, Stratigraphy and cataclysmic deposition of the Dry Mesa Dinosaur Quarry, Mesa County, Colorado, in Carpenter, K., Kirkland, J., and Chure, D., eds., The Upper Jurassic Morrison Formation: An Interdisciplinary Study, Modern Geology v. 22, no. 1-4, pp. 121–143.
- Chure, Daniel J.; Litwin, Ron; Hasiotis, Stephen T.; Evanoff, Emmett; Carpenter, Kenneth (2006). "The fauna and flora of the Morrison Formation: 2006". In Foster, John R.; Lucas, Spencer G. (eds.). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 233–248.
- OCTÁVIO MATEUS LATE JURASSIC DINOSAURS FROM THE MORRISON FORMATION (USA), THE LOURINHÃ AND ALCOBAÇA FORMATIONS (PORTUGAL), AND THE TENDAGURU BEDS (TANZANIA): A COMPARISON Foster, J.R. and Lucas, S. G. R.M., eds., 2006, Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36.
- Aberhan, Martin; Bussert, Robert; Heinrich, Wolf-Dieter; Schrank, Eckhart; Schultka, Stephan; Sames, Benjamin; Kriwet, Jürgen; Kapilima, Saidi (2002). "Palaeoecology and depositional environments of the Tendaguru Beds (Late Jurassic to Early Cretaceous, Tanzania)" (PDF). Fossil Record. 5 (1): 19–44. doi:10.1002/mmng.20020050103. Retrieved April 2, 2019.
- Soto, Matías; Toriño, Pablo; Perea, Daniel (November 1, 2020). "Ceratosaurus (Theropoda, Ceratosauria) teeth from the Tacuarembó Formation (Late Jurassic, Uruguay)". Journal of South American Earth Sciences. 103: 102781. Bibcode:2020JSAES.10302781S. doi:10.1016/j.jsames.2020.102781. ISSN 0895-9811. S2CID 224842133.
- Perea, D.; Ubilla, M.; Rojas, A. (2003). "First report of theropods from the Tacuarembó Formation (Late Jurassic–Early Cretaceous), Uruguay". Alcheringa. 27 (2): 79–83. Bibcode:2003Alch...27...79P. doi:10.1080/03115510308619548. S2CID 129634955.
- Perea, D.; Ubilla, M.; Rojas, A.; Goso, C.A. (November 2001). "The West Gondwanan occurrence of the hybodontid shark Priohybodus, and the Late Jurassic–Early Cretaceous age of the Tacuarembó Formation, Uruguay". Palaeontology. 44 (6): 1227–1235. Bibcode:2001Palgy..44.1227P. doi:10.1111/1475-4983.00222.
- Bakker, Robert T.; Bir, Gary (2004). "Dinosaur crime scene investigations: theropod behavior at Como Bluff, Wyoming, and the evolution of birdness". In Currie, Philip J.; Koppelhus, Eva B.; Shugar, Martin A.; Wright, Joanna L. (eds.). Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Bloomington and Indianapolis: Indiana University Press. pp. 301–342. ISBN 978-0-253-34373-4.
- Rauhut, Ow M; Hübner, T; Lanser, K (2016). "A new megalosaurid theropod dinosaur from the late Middle Jurassic (Callovian) of north-western Germany: Implications for theropod evolution and faunal turnover in the Jurassic". Palaeontologia Electronica. 19 (2): 1–65. doi:10.26879/654. ISSN 1094-8074.
- Nash, Duane (August 10, 2015). "Antediluvian Salad: Allosaurus - More of a Vulture Than a Falcon". Antediluvian Salad. Retrieved April 24, 2021.
- Chure, Daniel J. (2000). "Prey bone utilization by predatory dinosaurs in the Late Jurassic of North America, with comments on prey bone use by dinosaurs throughout the Mesozoic" (PDF). Gaia. 15: 227–232. ISSN 0871-5424.
- Drumheller, Stephanie K.; McHugh, Julia B.; Kane, Miriam; Riedel, Anja; D'Amore, Domenic C. (May 27, 2020). "High frequencies of theropod bite marks provide evidence for feeding, scavenging, and possible cannibalism in a stressed Late Jurassic ecosystem". PLOS ONE. 15 (5): e0233115. Bibcode:2020PLoSO..1533115D. doi:10.1371/journal.pone.0233115. ISSN 1932-6203. PMC 7252595. PMID 32459808.
External links
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||

.jpg.webp)