Non-homologous end joining
Non-homologous end joining (NHEJ) is a pathway that repairs double-strand breaks in DNA. It is called "non-homologous" because the break ends are directly ligated without the need for a homologous template, in contrast to homology directed repair (HDR), which requires a homologous sequence to guide repair. NHEJ is active in both non-dividing and proliferating cells, while HDR is not readily accessible in non-dividing cells.[1] The term "non-homologous end joining" was coined in 1996 by Moore and Haber.[2]
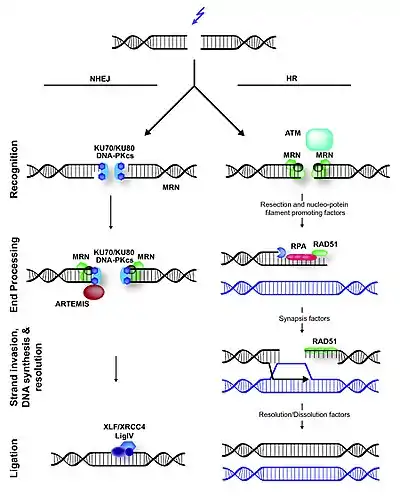
NHEJ is typically guided by short homologous DNA sequences called microhomologies. These microhomologies are often present in single-stranded overhangs on the ends of double-strand breaks. When the overhangs are perfectly compatible, NHEJ usually repairs the break accurately.[2][3][4][5] Imprecise repair leading to loss of nucleotides can also occur, but is much more common when the overhangs are not compatible. Inappropriate NHEJ can lead to translocations and telomere fusion, hallmarks of tumor cells.[6]
NHEJ implementations are understood to have been existent throughout nearly all biological systems and it is the predominant double-strand break repair pathway in mammalian cells.[7] In budding yeast (Saccharomyces cerevisiae), however, homologous recombination dominates when the organism is grown under common laboratory conditions.
When the NHEJ pathway is inactivated, double-strand breaks can be repaired by a more error-prone pathway called microhomology-mediated end joining (MMEJ). In this pathway, end resection reveals short microhomologies on either side of the break, which are then aligned to guide repair.[8] This contrasts with classical NHEJ, which typically uses microhomologies already exposed in single-stranded overhangs on the DSB ends. Repair by MMEJ therefore leads to deletion of the DNA sequence between the microhomologies.
In bacteria and archaea
Many species of bacteria, including Escherichia coli, lack an end joining pathway and thus rely completely on homologous recombination to repair double-strand breaks. NHEJ proteins have been identified in a number of bacteria, including Bacillus subtilis, Mycobacterium tuberculosis, and Mycobacterium smegmatis.[9][10] Bacteria utilize a remarkably compact version of NHEJ in which all of the required activities are contained in only two proteins: a Ku homodimer and the multifunctional ligase/polymerase/nuclease LigD.[11] In mycobacteria, NHEJ is much more error prone than in yeast, with bases often added to and deleted from the ends of double-strand breaks during repair.[10] Many of the bacteria that possess NHEJ proteins spend a significant portion of their life cycle in a stationary haploid phase, in which a template for recombination is not available.[9] NHEJ may have evolved to help these organisms survive DSBs induced during desiccation. It preferentially use rNTPs (RNA nucleotides), possibly advantageous in dormant cells.[12]
The archaeal NHEJ system in Methanocella paludicola have a homodimeric Ku, but the three functions of LigD are broken up into three single-domain proteins sharing an operon. All three genes retain substantial homology with their LigD counterparts and the polymerase retains the preference for rNTP.[13] NHEJ has been lost and acquired multiple times in bacteria and archaea, with a significant amount of horizontal gene transfer shuffling the system around taxa.[14]
Corndog and Omega, two related mycobacteriophages of Mycobacterium smegmatis, also encode Ku homologs and exploit the NHEJ pathway to recircularize their genomes during infection.[15] Unlike homologous recombination, which has been studied extensively in bacteria, NHEJ was originally discovered in eukaryotes and was only identified in prokaryotes in the past decade.
In eukaryotes
In contrast to bacteria, NHEJ in eukaryotes utilizes a number of proteins, which participate in the following steps:
End binding and tethering
In yeast, the Mre11-Rad50-Xrs2 (MRX) complex is recruited to DSBs early and is thought to promote bridging of the DNA ends.[16] The corresponding mammalian complex of Mre11-Rad50-Nbs1 (MRN) is also involved in NHEJ, but it may function at multiple steps in the pathway beyond simply holding the ends in proximity.[17] DNA-PKcs is also thought to participate in end bridging during mammalian NHEJ.[18]
Eukaryotic Ku is a heterodimer consisting of Ku70 and Ku80, and forms a complex with DNA-PKcs, which is present in mammals but absent in yeast. Ku is a basket-shaped molecule that slides onto the DNA end and translocates inward. Ku may function as a docking site for other NHEJ proteins, and is known to interact with the DNA ligase IV complex and XLF.[19][20]
End processing
End processing involves removal of damaged or mismatched nucleotides by nucleases and resynthesis by DNA polymerases. This step is not necessary if the ends are already compatible and have 3' hydroxyl and 5' phosphate termini.
Little is known about the function of nucleases in NHEJ. Artemis is required for opening the hairpins that are formed on DNA ends during V(D)J recombination, a specific type of NHEJ, and may also participate in end trimming during general NHEJ.[21] Mre11 has nuclease activity, but it seems to be involved in homologous recombination, not NHEJ.
The X family DNA polymerases Pol λ and Pol μ (Pol4 in yeast) fill gaps during NHEJ.[4][22][23] Yeast lacking Pol4 are unable to join 3' overhangs that require gap filling, but remain proficient for gap filling at 5' overhangs.[24] This is because the primer terminus used to initiate DNA synthesis is less stable at 3' overhangs, necessitating a specialized NHEJ polymerase.
Ligation
The DNA ligase IV complex, consisting of the catalytic subunit DNA ligase IV and its cofactor XRCC4 (Dnl4 and Lif1 in yeast), performs the ligation step of repair.[25] XLF, also known as Cernunnos, is homologous to yeast Nej1 and is also required for NHEJ.[26][27] While the precise role of XLF is unknown, it interacts with the XRCC4/DNA ligase IV complex and likely participates in the ligation step.[28] Recent evidence suggests that XLF promotes re-adenylation of DNA ligase IV after ligation, recharging the ligase and allowing it to catalyze a second ligation.[29]
Other
In yeast, Sir2 was originally identified as an NHEJ protein, but is now known to be required for NHEJ only because it is required for the transcription of Nej1.[30]
NHEJ and heat-labile sites
Induction of heat-labile sites (HLS) is a signature of ionizing radiation. The DNA clustered damage sites consist of different types of DNA lesions. Some of these lesions are not prompt DSBs but they convert to DSB after heating. HLS are not evolved to DSB under physiological temperature (370 C). Also, the interaction of HLS with other lesions and their role in living cells is yet elusive. The repair mechanisms of these sites are not fully revealed. The NHEJ is the dominant DNA repair pathway throughout the cell cycle. The DNA-PKcs protein is the critical element in the center of NHEJ. Using DNA-PKcs KO cell lines or inhibition of DNA-PKcs does not affect the repair capacity of HLS. Also blocking both HR and NHEJ repair pathways by dactolisib (NVP-BEZ235) inhibitor showed that repair of HLS is not dependent on HR and NHEJ. These results showed that the repair mechanism of HLS is independent of NHEJ and HR pathways[31]
Regulation
The choice between NHEJ and homologous recombination for repair of a double-strand break is regulated at the initial step in recombination, 5' end resection. In this step, the 5' strand of the break is degraded by nucleases to create long 3' single-stranded tails. DSBs that have not been resected can be rejoined by NHEJ, but resection of even a few nucleotides strongly inhibits NHEJ and effectively commits the break to repair by recombination.[23] NHEJ is active throughout the cell cycle, but is most important during G1 when no homologous template for recombination is available. This regulation is accomplished by the cyclin-dependent kinase Cdk1 (Cdc28 in yeast), which is turned off in G1 and expressed in S and G2. Cdk1 phosphorylates the nuclease Sae2, allowing resection to initiate.[32]
V(D)J recombination
NHEJ plays a critical role in V(D)J recombination, the process by which B-cell and T-cell receptor diversity is generated in the vertebrate immune system.[33] In V(D)J recombination, hairpin-capped double-strand breaks are created by the RAG1/RAG2 nuclease, which cleaves the DNA at recombination signal sequences.[34] These hairpins are then opened by the Artemis nuclease and joined by NHEJ.[21] A specialized DNA polymerase called terminal deoxynucleotidyl transferase (TdT), which is only expressed in lymph tissue, adds nontemplated nucleotides to the ends before the break is joined.[35][36] This process couples "variable" (V), "diversity" (D), and "joining" (J) regions, which when assembled together create the variable region of a B-cell or T-cell receptor gene. Unlike typical cellular NHEJ, in which accurate repair is the most favorable outcome, error-prone repair in V(D)J recombination is beneficial in that it maximizes diversity in the coding sequence of these genes. Patients with mutations in NHEJ genes are unable to produce functional B cells and T cells and suffer from severe combined immunodeficiency (SCID).
At telomeres
Telomeres are normally protected by a "cap" that prevents them from being recognized as double-strand breaks. Loss of capping proteins causes telomere shortening and inappropriate joining by NHEJ, producing dicentric chromosomes which are then pulled apart during mitosis. Paradoxically, some NHEJ proteins are involved in telomere capping. For example, Ku localizes to telomeres and its deletion leads to shortened telomeres.[37] Ku is also required for subtelomeric silencing, the process by which genes located near telomeres are turned off.
Consequences of dysfunction
Several human syndromes are associated with dysfunctional NHEJ.[38] Hypomorphic mutations in LIG4 and XLF cause LIG4 syndrome and XLF-SCID, respectively. These syndromes share many features including cellular radiosensitivity, microcephaly and severe combined immunodeficiency (SCID) due to defective V(D)J recombination. Loss-of-function mutations in Artemis also cause SCID, but these patients do not show the neurological defects associated with LIG4 or XLF mutations. The difference in severity may be explained by the roles of the mutated proteins. Artemis is a nuclease and is thought to be required only for repair of DSBs with damaged ends, whereas DNA Ligase IV and XLF are required for all NHEJ events. Mutations in genes that participate in non-homologous end joining lead to ataxia-telangiectasia (ATM gene), Fanconi anemia (multiple genes), as well as hereditary breast and ovarian cancers (BRCA1 gene).
Many NHEJ genes have been knocked out in mice. Deletion of XRCC4 or LIG4 causes embryonic lethality in mice, indicating that NHEJ is essential for viability in mammals. In contrast, mice lacking Ku or DNA-PKcs are viable, probably because low levels of end joining can still occur in the absence of these components.[39] All NHEJ mutant mice show a SCID phenotype, sensitivity to ionizing radiation, and neuronal apoptosis.
Aging
A system was developed for measuring NHEJ efficiency in the mouse.[40] NHEJ efficiency could be compared across tissues of the same mouse and in mice of different age. Efficiency was higher in the skin, lung and kidney fibroblasts, and lower in heart fibroblasts and brain astrocytes. Furthermore, NHEJ efficiency declined with age. The decline was 1.8 to 3.8-fold, depending on the tissue, in the 5-month-old compared to the 24-month-old mice. Reduced capability for NHEJ can lead to an increase in the number of unrepaired or faultily repaired DNA double-strand breaks that may then contribute to aging.[41] (Also see DNA damage theory of aging.) An analysis of the level of NHEJ protein Ku80 in human, cow, and mouse indicated that Ku80 levels vary dramatically between species, and that these levels are strongly correlated with species longevity.[42]
List of proteins involved in NHEJ in human cells
References
- Chou, Shih‐Jie; Yang, Peng; Ban, Qian; Yang, Yi‐Ping; Wang, Mong‐Lien; Chien, Chian‐Shiu; Chen, Shih‐Jen; Sun, Na; Zhu, Yazhen; Liu, Hongtao; Hui, Wenqiao; Lin, Tai‐Chi; Wang, Fang; Zhang, Ryan Yue; Nguyen, Viet Q. (May 2020). "Dual Supramolecular Nanoparticle Vectors Enable CRISPR/Cas9‐Mediated Knockin of Retinoschisin 1 Gene—A Potential Nonviral Therapeutic Solution for X‐Linked Juvenile Retinoschisis". Advanced Science. 7 (10): 1903432. doi:10.1002/advs.201903432. ISSN 2198-3844. PMC 7237855. PMID 32440478.
- Moore JK, Haber JE (May 1996). "Cell cycle and genetic requirements of two pathways of nonhomologous end-joining repair of double-strand breaks in Saccharomyces cerevisiae". Molecular and Cellular Biology. 16 (5): 2164–73. doi:10.1128/mcb.16.5.2164. PMC 231204. PMID 8628283.
- Boulton SJ, Jackson SP (September 1996). "Saccharomyces cerevisiae Ku70 potentiates illegitimate DNA double-strand break repair and serves as a barrier to error-prone DNA repair pathways". EMBO J. 15 (18): 5093–103. doi:10.1002/j.1460-2075.1996.tb00890.x. PMC 452249. PMID 8890183.
- Wilson TE, Lieber MR (1999). "Efficient processing of DNA ends during yeast nonhomologous end joining. Evidence for a DNA polymerase beta (Pol4)-dependent pathway". J. Biol. Chem. 274 (33): 23599–23609. doi:10.1074/jbc.274.33.23599. PMID 10438542.
- Budman J, Chu G (Feb 2005). "Processing of DNA for nonhomologous end-joining by cell-free extract". EMBO J. 24 (4): 849–60. doi:10.1038/sj.emboj.7600563. PMC 549622. PMID 15692565.
- Espejel S, Franco S, Rodríguez-Perales S, Bouffler SD, Cigudosa JC, Blasco MA (May 2002). "Mammalian Ku86 mediates chromosomal fusions and apoptosis caused by critically short telomeres". The EMBO Journal. 21 (9): 2207–19. doi:10.1093/emboj/21.9.2207. PMC 125978. PMID 11980718.
- Guirouilh-Barbat J, Huck S, Bertrand P, et al. (June 2004). "Impact of the KU80 pathway on NHEJ-induced genome rearrangements in mammalian cells". Mol. Cell. 14 (5): 611–23. doi:10.1016/j.molcel.2004.05.008. PMID 15175156.
- McVey M, Lee SE (November 2008). "MMEJ repair of double-strand breaks (director's cut): deleted sequences and alternative endings". Trends Genet. 24 (11): 529–38. doi:10.1016/j.tig.2008.08.007. PMC 5303623. PMID 18809224.
- Weller GR, Kysela B, Roy R, et al. (September 2002). "Identification of a DNA nonhomologous end-joining complex in bacteria". Science. 297 (5587): 1686–9. Bibcode:2002Sci...297.1686W. doi:10.1126/science.1074584. PMID 12215643. S2CID 20135110.
- Gong C, Bongiorno P, Martins A, et al. (April 2005). "Mechanism of nonhomologous end-joining in mycobacteria: a low-fidelity repair system driven by Ku, ligase D and ligase C". Nat. Struct. Mol. Biol. 12 (4): 304–12. doi:10.1038/nsmb915. PMID 15778718. S2CID 6879518.
- Della M, Palmbos PL, Tseng HM, et al. (October 2004). "Mycobacterial Ku and ligase proteins constitute a two-component NHEJ repair machine". Science. 306 (5696): 683–5. Bibcode:2004Sci...306..683D. doi:10.1126/science.1099824. PMID 15499016. S2CID 38823696.
- Pitcher RS, Green AJ, Brzostek A, Korycka-Machala M, Dziadek J, Doherty AJ (September 2007). "NHEJ protects mycobacteria in stationary phase against the harmful effects of desiccation" (PDF). DNA Repair (Amst.). 6 (9): 1271–6. doi:10.1016/j.dnarep.2007.02.009. PMID 17360246.
- Bartlett, EJ; Brissett, NC; Doherty, AJ (28 May 2013). "Ribonucleolytic resection is required for repair of strand displaced nonhomologous end-joining intermediates". Proceedings of the National Academy of Sciences of the United States of America. 110 (22): E1984-91. Bibcode:2013PNAS..110E1984B. doi:10.1073/pnas.1302616110. PMC 3670387. PMID 23671117.
- Sharda, Mohak; Badrinarayanan, Anjana; Seshasayee, Aswin Sai Narain (6 December 2020). "Evolutionary and Comparative Analysis of Bacterial Nonhomologous End Joining Repair". Genome Biology and Evolution. 12 (12): 2450–2466. doi:10.1093/gbe/evaa223. PMC 7719229. PMID 33078828.
- Pitcher RS, Tonkin LM, Daley JM, et al. (September 2006). "Mycobacteriophage exploit NHEJ to facilitate genome circularization". Mol. Cell. 23 (5): 743–8. doi:10.1016/j.molcel.2006.07.009. PMID 16949369.
- Chen L, Trujillo K, Ramos W, Sung P, Tomkinson AE (2001). "Promotion of Dnl4-catalyzed DNA end-joining by the Rad50/Mre11/Xrs2 and Hdf1/Hdf2 complexes". Mol Cell. 8 (5): 1105–1115. doi:10.1016/s1097-2765(01)00388-4. PMID 11741545.
- Zha S, Boboila C, Alt FW (August 2009). "Mre11: roles in DNA repair beyond homologous recombination". Nat. Struct. Mol. Biol. 16 (8): 798–800. doi:10.1038/nsmb0809-798. PMID 19654615. S2CID 205522532.
- DeFazio LG, Stansel RM, Griffith JD, Chu G (June 2002). "Synapsis of DNA ends by DNA-dependent protein kinase". The EMBO Journal. 21 (12): 3192–200. doi:10.1093/emboj/cdf299. PMC 126055. PMID 12065431.
- Palmbos PL, Wu D, Daley JM, Wilson TE (December 2008). "Recruitment of Saccharomyces cerevisiae Dnl4-Lif1 complex to a double-strand break requires interactions with Yku80 and the Xrs2 FHA domain". Genetics. 180 (4): 1809–19. doi:10.1534/genetics.108.095539. PMC 2600923. PMID 18832348.
- Yano K, Morotomi-Yano K, Wang SY, et al. (January 2008). "Ku recruits XLF to DNA double-strand breaks". EMBO Rep. 9 (1): 91–6. doi:10.1038/sj.embor.7401137. PMC 2246615. PMID 18064046.
- Ma Y, Pannicke U, Schwarz K, Lieber MR (2002). "Hairpin opening and overhang processing by an Artemis/DNA-dependent protein kinase complex in nonhomologous end joining and V(D)J recombination". Cell. 108 (6): 781–794. doi:10.1016/s0092-8674(02)00671-2. PMID 11955432.
- Nick McElhinny SA, Ramsden DA (August 2004). "Sibling rivalry: competition between Pol X family members in V(D)J recombination and general double strand break repair". Immunol. Rev. 200: 156–64. doi:10.1111/j.0105-2896.2004.00160.x. PMID 15242403. S2CID 36516952.
- Daley JM, Laan RL, Suresh A, Wilson TE (August 2005). "DNA joint dependence of pol X family polymerase action in nonhomologous end joining". J. Biol. Chem. 280 (32): 29030–7. doi:10.1074/jbc.M505277200. PMID 15964833.
- Daley JM, Laan RL, Suresh A, Wilson TE (August 2005). "DNA joint dependence of pol X family polymerase action in nonhomologous end joining". J. Biol. Chem. 280 (32): 29030–7. doi:10.1074/jbc.M505277200. PMID 15964833.
- Wilson T. E.; Grawunder U.; Lieber M. R. (1997). "Yeast DNA ligase IV mediates non-homologous DNA end joining". Nature. 388 (6641): 495–498. Bibcode:1997Natur.388..495W. doi:10.1038/41365. PMID 9242411.
- Ahnesorg P, Smith P, Jackson SP (Jan 2006). "XLF interacts with the XRCC4-DNA ligase IV complex to promote DNA nonhomologous end-joining". Cell. 124 (2): 301–13. doi:10.1016/j.cell.2005.12.031. PMID 16439205.
- Buck D, Malivert L, de Chasseval R, Barraud A, Fondaneche MC, Sanal O, Plebani A, Stephan JL, Hufnagel M, et al. (Jan 2006). "Cernunnos, a novel nonhomologous end-joining factor, is mutated in human immunodeficiency with microcephaly". Cell. 124 (2): 287–99. doi:10.1016/j.cell.2005.12.030. PMID 16439204.
- Callebaut I, Malivert L, Fischer A, Mornon JP, Revy P, de Villartay JP (2006). "Cernunnos Interacts with the XRCC4•DNA-ligase IV Complex and Is Homologous to the Yeast Nonhomologous End-joining Factor Nej1". J Biol Chem. 281 (20): 13857–60. doi:10.1074/jbc.C500473200. PMID 16571728.
- Riballo E, Woodbine L, Stiff T, Walker SA, Goodarzi AA, Jeggo PA (February 2009). "XLF-Cernunnos promotes DNA ligase IV-XRCC4 re-adenylation following ligation". Nucleic Acids Res. 37 (2): 482–92. doi:10.1093/nar/gkn957. PMC 2632933. PMID 19056826.
- Lee SE, Pâques F, Sylvan J, Haber JE (July 1999). "Role of yeast SIR genes and mating type in directing DNA double-strand breaks to homologous and non-homologous repair paths". Curr. Biol. 9 (14): 767–70. doi:10.1016/s0960-9822(99)80339-x. PMID 10421582.
- Abramenkovs, A., & Stenerlöw, B. (2018). Removal of heat-sensitive clustered damaged DNA sites is independent of double-strand break repair. Plos one, 13(12), e0209594.
- Mimitou EP, Symington LS (September 2009). "DNA end resection: Many nucleases make light work". DNA Repair (Amst.). 8 (9): 983–95. doi:10.1016/j.dnarep.2009.04.017. PMC 2760233. PMID 19473888.
- Jung D, Alt FW (Jan 2004). "Unraveling V(D)J recombination; insights into gene regulation". Cell. 116 (2): 299–311. doi:10.1016/S0092-8674(04)00039-X. PMID 14744439.
- Schatz DG, Baltimore D (April 1988). "Stable expression of immunoglobulin gene V(D)J recombinase activity by gene transfer into 3T3 fibroblasts". Cell. 53 (1): 107–15. doi:10.1016/0092-8674(88)90492-8. PMID 3349523. S2CID 42040516.
- Gilfillan S, Dierich A, Lemeur M, Benoist C, Mathis D (August 1993). "Mice lacking TdT: mature animals with an immature lymphocyte repertoire". Science. 261 (5125): 1175–8. Bibcode:1993Sci...261.1175G. doi:10.1126/science.8356452. PMID 8356452. S2CID 36801225.
- Komori T, Okada A, Stewart V, Alt FW (August 1993). "Lack of N regions in antigen receptor variable region genes of TdT-deficient lymphocytes". Science. 261 (5125): 1171–5. Bibcode:1993Sci...261.1171K. doi:10.1126/science.8356451. PMID 8356451.
- Boulton SJ, Jackson SP (1998). "Components of the Ku-dependent non-homologous endjoining pathway are involved in telomeric length maintenance and telomeric silencing". EMBO J. 17 (6): 1819–28. doi:10.1093/emboj/17.6.1819. PMC 1170529. PMID 9501103.
- Kerzendorfer C, O'Driscoll M (September 2009). "Human DNA damage response and repair deficiency syndromes: Linking genomic instability and cell cycle checkpoint proficiency". DNA Repair (Amst.). 8 (9): 1139–52. doi:10.1016/j.dnarep.2009.04.018. PMID 19473885.
- Li H, Vogel H, Holcomb VB, Gu Y, Hasty P (December 2007). "Deletion of Ku70, Ku80, or both causes early aging without substantially increased cancer". Mol. Cell. Biol. 27 (23): 8205–14. doi:10.1128/MCB.00785-07. PMC 2169178. PMID 17875923.
- Vaidya A, Mao Z, Tian X, Spencer B, Seluanov A, Gorbunova V (2014). "Knock-in reporter mice demonstrate that DNA repair by non-homologous end joining declines with age". PLOS Genet. 10 (7): e1004511. doi:10.1371/journal.pgen.1004511. PMC 4102425. PMID 25033455.
- Bernstein H, Payne CM, Bernstein C, Garewal H, Dvorak K (2008). Cancer and aging as consequences of un-repaired DNA damage. In: New Research on DNA Damages (Editors: Honoka Kimura and Aoi Suzuki) Nova Science Publishers, Inc., New York, Chapter 1, pp. 1-47. open access, but read only https://www.novapublishers.com/catalog/product_info.php?products_id=43247 Archived 2014-10-25 at the Wayback Machine ISBN 978-1604565812
- Lorenzini A, Johnson FB, Oliver A, Tresini M, Smith JS, Hdeib M, Sell C, Cristofalo VJ, Stamato TD (2009). "Significant correlation of species longevity with DNA double strand break recognition but not with telomere length". Mech. Ageing Dev. 130 (11–12): 784–92. doi:10.1016/j.mad.2009.10.004. PMC 2799038. PMID 19896964.
| Excision repair | |
|---|---|
| Other forms of repair | |
| Other/ungrouped proteins | |
| Regulation | |
| Other/ungrouped | |
| |