NOD-like receptor
The nucleotide-binding oligomerization domain-like receptors, or NOD-like receptors (NLRs) (also known as nucleotide-binding leucine-rich repeat receptors),[1] are intracellular sensors of pathogen-associated molecular patterns (PAMPs) that enter the cell via phagocytosis or pores, and damage-associated molecular patterns (DAMPs) that are associated with cell stress. They are types of pattern recognition receptors (PRRs),[2] and play key roles in the regulation of innate immune response. NLRs can cooperate with toll-like receptors (TLRs) and regulate inflammatory and apoptotic response.
NLRs primarily recognize Gram-positive bacteria, whereas TLRs primarily recognize Gram-negative bacteria. They are found in lymphocytes, macrophages, dendritic cells and also in non-immune cells, for example in epithelium.[3] NLRs are highly conserved through evolution. Their homologs have been discovered in many different animal species (APAF1) [4][5] and also in the plant kingdom (disease-resistance R protein).[5]
Structure
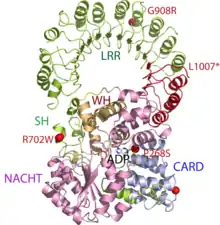
NLRs contain 3 domains – central NACHT (NOD or NBD – nucleotide-binding domain) domain, which is common to all NLRs, most of NLRs have also C-terminal leucine-rich repeat (LRR) and variable N-terminal interaction domain. NACHT domain mediates ATP-dependent self-oligomerization and LRR senses the presence of ligand. N-terminal domain is responsible for homotypic protein-protein interaction and it can consist of caspase recruitment domain (CARD), pyrin domain (PYD), acidic transactivating domain or baculovirus inhibitor repeats (BIRs).[3][6]
Nomenclature and system
Names as CATERPILLER, NOD, NALP, PAN, NACHT, PYPAF were used to describe the NLRs family. The nomenclature was unified by the HUGO Gene Nomenclature Committee in 2008. The family was characterized as NLRs to provide description of the families features – NLR means nucleotide-binding domain and leucine-rich repeat containing gene family.[7]
This system divides NLRs into 4 subfamilies based on the type of N-terminal domain:
- NLRA (A for acidic transactivating domain): CIITA
- NLRB (B for BIRs): NAIP
- NLRC (C for CARD): NOD1, NOD2, NLRC3, NLRC4, NLRC5
- NLRP (P for PYD): NLRP1, NLRP2, NLRP3, NLRP4, NLRP5, NLRP6, NLRP7, NLRP8, NLRP9, NLRP10, NLRP11, NLRP12, NLRP13, NLRP14[7]
There is also an additional subfamily NLRX which doesn't have significant homology to any N-terminal domain. A member of this subfamily is NLRX1.[8]
On the other hand, NLRs can be divided into 3 subfamilies with regard to their phylogenetic relationships:
Subfamily NODs
NODs subfamily consists of NOD1, NOD2, NOD3, NOD4 with CARD domain, CIITA containing acidic transactivator domain and NOD5 without any N-terminal domain.[9] [10]
Signalling
The well-described receptors are NOD1 and NOD2. The recognition of their ligands recruits oligomerization of NACHT domain and CARD-CARD interaction with CARD-containing serine-threonin kinase RIP2 which leads to activation of RIP2.[11] RIP2 mediates the recruitment of kinase TAK1 which phosphorylates and activates IκB kinase. The activation of IκB kinase results in the phosphorylation of inhibitor IκB which releases NF-κB and its nuclear translocation. NF-κB then activates expression of inflammatory cytokines.[12] Mutations in NOD2 are associated with Crohn's disease[13] or Blau syndrome.[14]
Ligands
NOD1 and NOD2 recognize peptidoglycan motifs from bacterial cell which consists of N-acetylglucosamine and N-acetylmuramic acid. These sugar chains are cross-linked by peptide chains that can be sensed by NODs. NOD1 recognizes a molecule called meso-diaminopimelic acid (meso-DAP) mostly found in Gram-negative bacteria (for example Helicobacter pylori, Pseudomonas aeruginosa). NOD2 proteins can sense intracellular muramyl dipeptide (MDP), typical for bacteria such as Streptococcus pneumoniae or Mycobacterium tuberculosis.[3][10]
Subfamilies NLRPs and IPAF
NLRPs subfamily contains NLRP1-NLRP14 that are characterized by the presence of PYD domain. IPAF subfamily has two members – IPAF with CARD domain and NAIP with BIR domain.[9][10]
Signalization
NLRPs and IPAF subfamilies are involved in the formation of the inflammasome. The best characterized inflammasome is NLRP3, the activation through PAMPs or DAMPs leads to the oligomerization.[9] The pyrin domain of NLRs binds to an adaptor protein ASC (PYCARD) via PYD-PYD interaction. ASC contains PYD and CARD domain and links the NLRs to inactive form of caspase 1 through the CARD domain.[15] All these protein-protein interaction form a complex called the inflammasome. The aggregation of the pro-caspase-1 causes the autocleavage and formation of an active enzyme. Caspase-1 is important for the proteolytic processing of the pro-inflammatory cytokines IL-1β and IL-18.[9][10] NLRP3 mutations are responsible for the autoinflammatory disease familial cold autoinflammatory syndrome or Muckle–Wells syndrome.[16][17]
Ligands
There are three well-characterized inflammasomes – NLRP1, NLRP3 and IPAF. The formation of NLRP3 inflammasome can be activated by PAMPs such as microbial toxins (for example alpha-toxin of Staphylococcus aureus) or whole pathogens, for instance Candida albicans, Saccharomyces cerevisiae, Sendai virus, Influenza. NLRP3 recognize also DAMPs which indicate stress in the cell. The danger molecule can be extracellular ATP, extracellular glucose, monosodium urate (MSU) crystals, calcium pyrophosphate dihydrate (CPPD), alum, cholesterol or environmental irritants – silica, asbestos, UV irradiation and skin irritants. The presence of these molecules causes a production of ROS and K+ efflux. NLRP1 recognizes lethal toxin from Bacillus anthracis and muramyl dipeptide. IPAF senses flagellin from Salmonella typhimurium, Pseudomonas aeruginosa, Listeria monocytogenes.[3][9][10]
References
- Mahla, Ranjeet (2013). "Sweeten PAMPs: Role of Sugar Complexed PAMPs in Innate Immunity and Vaccine Biology". Front Immunol. 4: 248. doi:10.3389/fimmu.2013.00248. PMC 3759294. PMID 24032031.
- Mahla RS, Reddy MC, Prasad DV, Kumar H (September 2013). "Sweeten PAMPs: Role of Sugar Complexed PAMPs in Innate Immunity and Vaccine Biology". Frontiers in Immunology. 4: 248. doi:10.3389/fimmu.2013.00248. PMC 3759294. PMID 24032031.
- Franchi L, Warner N, Viani K, Nuñez G (2009). "Function of Nod-like receptors in microbial recognition and host defense". Immunol Rev. 227 (1): 106–28. doi:10.1111/j.1600-065X.2008.00734.x. PMC 2679989. PMID 19120480.
- Ogura Y, Inohara N, Benito A, Chen FF, Yamaoka S, Nunez G (2001). "Nod2, a Nod1/Apaf-1 family member that is restricted to monocytes and activates NF-kappaB". J Biol Chem. 276 (7): 4812–8. doi:10.1074/jbc.M008072200. PMID 11087742.
- Inohara N, Ogura Y, Chen FF, Muto A, Nuñez G (2001). "Human Nod1 confers responsiveness to bacterial lipopolysaccharides". J Biol Chem. 276 (4): 2551–4. doi:10.1074/jbc.M009728200. PMID 11058605.
- Shaw MH, Reimer T, Kim YG, Nuñez G (2008). "NOD-like receptors (NLRs): bona fide intracellular microbial sensors". Curr Opin Immunol. 20 (4): 377–82. doi:10.1016/j.coi.2008.06.001. PMC 2572576. PMID 18585455.
- Ting JP, Lovering RC, Alnemri ES, Bertin J, Boss JM, Davis BK, et al. (2008). "The NLR gene family: a standard nomenclature". Immunity. 28 (3): 285–7. doi:10.1016/j.immuni.2008.02.005. PMC 2630772. PMID 18341998.
- Tattoli I, Carneiro LA, Jéhanno M, Magalhaes JG, Shu Y, Philpott DJ, et al. (2008). "NLRX1 is a mitochondrial NOD-like receptor that amplifies NF-kappaB and JNK pathways by inducing reactive oxygen species production". EMBO Rep. 9 (3): 293–300. doi:10.1038/sj.embor.7401161. PMC 2267388. PMID 18219313.
- Schroder K, Tschopp J (2010). "The inflammasomes". Cell. 140 (6): 821–32. doi:10.1016/j.cell.2010.01.040. PMID 20303873.
- Chen G, Shaw MH, Kim YG, Nuñez G (2009). "NOD-like receptors: role in innate immunity and inflammatory disease". Annu Rev Pathol. 4: 365–98. doi:10.1146/annurev.pathol.4.110807.092239. PMID 18928408.
- Park JH, Kim YG, McDonald C, Kanneganti TD, Hasegawa M, Body-Malapel M, et al. (2007). "RICK/RIP2 mediates innate immune responses induced through Nod1 and Nod2 but not TLRs". J Immunol. 178 (4): 2380–6. doi:10.4049/jimmunol.178.4.2380. PMID 17277144.
- Hasegawa M, Fujimoto Y, Lucas PC, Nakano H, Fukase K, Núñez G, et al. (2008). "A critical role of RICK/RIP2 polyubiquitination in Nod-induced NF-kappaB activation". EMBO J. 27 (2): 373–83. doi:10.1038/sj.emboj.7601962. PMC 2234345. PMID 18079694.
- Cantó E, Ricart E, Busquets D, Monfort D, García-Planella E, González D, et al. (2007). "Influence of a nucleotide oligomerization domain 1 (NOD1) polymorphism and NOD2 mutant alleles on Crohn's disease phenotype". World J Gastroenterol. 13 (41): 5446–53. doi:10.3748/wjg.v13.i41.5446. PMC 4171278. PMID 17907287.
- Okafuji I, Nishikomori R, Kanazawa N, Kambe N, Fujisawa A, Yamazaki S, et al. (2009). "Role of the NOD2 genotype in the clinical phenotype of Blau syndrome and early-onset sarcoidosis". Arthritis Rheum. 60 (1): 242–50. doi:10.1002/art.24134. hdl:2433/124253. PMID 19116920.
- Srinivasula SM, Poyet JL, Razmara M, Datta P, Zhang Z, Alnemri ES (2002). "The PYRIN-CARD protein ASC is an activating adaptor for caspase-1". J Biol Chem. 277 (24): 21119–22. doi:10.1074/jbc.C200179200. PMID 11967258.
- Hoffman HM, Mueller JL, Broide DH, Wanderer AA, Kolodner RD (2001). "Mutation of a new gene encoding a putative pyrin-like protein causes familial cold autoinflammatory syndrome and Muckle-Wells syndrome". Nat Genet. 29 (3): 301–5. doi:10.1038/ng756. PMC 4322000. PMID 11687797.
- Kubota T, Koike R (2007). "[Biological and clinical aspects of Muckle-Wells syndrome]". Nihon Rinsho Meneki Gakkai Kaishi. 30 (2): 114–22. doi:10.2177/jsci.30.114. PMID 17473514.