Paleobiota of the Posidonia Shale
The Sachrang Formation or "Posidonienschiefer" Formation (Whose Vulgar name is "Posidonia Shale") is a geological formation of southwestern Germany, northern Switzerland, northwestern Austria, southeast Luxembourg and the Netherlands, that spans about 3 million years during the Early Jurassic period (early Toarcian stage). It is known for its detailed fossils, especially sea fauna, listed below.[1] Composed mostly by black shale, the formation is a Lagerstätte, where fossils show exceptional preservation (Including exquisite soft tissues), with a thickness that varies from about 1 m to about 40 m on the Rhine level, being on the main quarry at Holzmaden between 5 and 14 m.[1] Some of the preserved material has been transformed into fossil hydrocarbon Jet, specially wood remains, used for jewelry.[2] The exceptional preservation seen on the Posidonia Shale has been studied since the late 1800s, finding that a cocktail of chemical and environmental factors let to such an impressive conservation of the marine fauna.[2] The most common theory is the changes in the oxygen level, where the different anoxic events of the Toarcian left oxygen-depleted bottom waters, with the biota dying and falling to the bottom without any predator able to eat the dead bodies.[3]
.jpg.webp)
| Part of a series on |
| Paleontology |
|---|
 |
|
Paleontology Portal Category |
Biological interaction frozen in time
- Several animal behaviours have been recovered on the Posidonia Shale. The Monotis-Dactylioceras bed is one of them, as it shows an accumulation of the bivalves Meleagrinella substriata and the ammonite Dactylioceras, that were the most abundant representatives of its group on the Altdorf region, and were probably washed to near epicontinental waters by a rapid event, or as result of a large succession of events.[4] This assemblage has been compared with modern Brazilian coastal Mangroves and also linked to Tsunami events.[5]
- Related to the Ammonite fauna, on Holzmaden there have been found several empty shells of this cephalopods, with associated crustaceans inside.[6] The original specimen was reported in 1995 and consisted on a possible member of the genus Paleastacus inside a chamber of a Harpoceras.[6] Other epizoans are found related to the decayed Ammonite shells, such as Serpulid Annelids and Bivalves, creating what was denominated as "benthic islands" as reference at be isolated benthic units that attracted fauna.[6] The Decapod is related to the family Erymidae, that are considered as possible bottom-dweller carnivorous or carrion feeders.[6] The associated fossil has several spherical structures that had been interpreted as decapod coprolites, implying that the animal lived for a long period on the shell, and maybe changes on the bottom oxygen stopped the process.[6] More recent studies had recovered new data about the inquilinism of decapods inside ammonites, this time, however, recovering three Eryonoidea lobsters together within a body chamber.[7] The lobsters most likely used the ammonoid as some kind of shelter, where was excluded the possibility of transportation into the body chamber by bottom currents.[7] There are several theories about the gregarious inquilinism showed by this specimens, such as that the shell was and ideal location to molt, as has not been proved to be corpses or molts; that the shell provided protection against predators; the decomposing soft body of the ammonoid was a source of food that attracted the decapods or that was used as a long-term shelter.[7] One key aspect found was that the muddy bottom was not suitable for burrowing, implying that the decapods look for a different shelter due to being unable to make their own.[7]
- Beyond trace fossils, several vertebrate specimens show associations with crustacean exoskeletal remains such as GPIT-PV-31586 and SMNS 58389 (Pachycormus macropterus) with necrophagous interaction as well SMNS 55934 (Stenopterygius quadriscissus) or SMNS 95401 (Metopacanthus sp.).[8]
- The genus Clarkeiteuthis and its predatory behaviour, found associated with fishes of the genus Leptolepis.[9] Based on the position of prey and predator, was suggested that the coeloid cephalopods caught and killed the fishes while the schools still in well-oxygenated waters and then descended into oxygen-depleted water layers where the cephalopod suffocated and died attached to its prey.[9] The fish measured 12 centimetres (4.7 in), while the coeloid 21 and it was measured by the fossilized arms of 14 specimens of coeloids that the hunting specimens arms where contracted over the fish, probably quickly killing it by cutting its spine.[9]
- Several Geotheutis have been reported with eumelanin preserved along with its ink sacs.[10]
- A specimen of Jeletzkyteuthis found on Ohmden has appeared predating a Parabelopeltis. The association of this 2 genera shows the predatory behaviour of this group when lived on Epicontinental seas, being rather different than extant Vampyromorphs.[11]
- A Pabulite (fossilized meal when it never entered the digestive tract) was recovered on Holzmaden, being composed by an associated Passaloteuthis laevigata with its arms embracing an exuvia of a crustacean.[12] The own Belemnnite can be the remnant of a failed prey of a Hybodus, corroborating a possible tropic chain.[12]

- One of the most complex organism interactions on the Posidonia Shale where the crinoid megarafts, that group a wide variety of animals, creating large floating ecosystems, being the longest surviving communities to exist in the fossil record.[13] The largest megaraft found measured 18 metres (59 ft), and is based on an Agathoxylon trunk, where different animals were attached.[13] The first attached animals would have been the growing community of oysters, bivalves and crinoids, that would suppose and small weight to the raft about 800 kilograms (1,800 lb).[13] The presence of this megaraft was in part possible due to the absence of marine wood worms, that destroy tree logs on less than 3 years along without the presence of modern raft wood predators (that appeared on the Bathonian) those rafts can last up to 5 years, being that the main reason the crinoids attached were able to reach huge sizes.[13] Probably where also essential to distribute animals along the sea basins.[13] Seirocrinus & Pentacrinites where various of the main crinoid colonizers of the floating rafts.[14] Seirocrinus is the main representative of the pelagic crinoids, being among the tallest animals know, with a size of 26 m the largest documented specimen.[14] The ecology of the genus is widely known, where is known that the smallest stems were among the first animals to colonize the rafts, with at least 2 generations of crinoids found per raft, where the hydrodynamic changes of the log influenced the settlement of the crinoids.[14] It is believed that Seridocrinus had a seasonal reproduction, linked to the monsoonal conditions that sent new logs to the sea.[14] The large crinoids would have feed on pelagic micronutrients, and afer fall on the bottom, all the colony would have died.[14]
- Thoracic cirripedes of the genus Toarcolepas became the oldest epiplanktonic cirripede known on the fossil record, probably motivated by the appearance of the giant crinoid rafts. It has been found in situ associated with fossil wood.[15]

- The shark Hybodus includes specimens with the gastric contents, being full of belemnnite fragments.[16] That implied active predatory behaviour by the genus of several kinds of belemnnites, such as Youngibelus.[16]
- A Spienballen, a regurgitated mass composed of indigestible stomach contents had been found on the Holzmaden quarry.[17] The Speinballen measures 285 mm length with a diameter of 160 mm, and consists of 4 members of the genus Dapedium (Dapediidae) and a jaw identified as Lepidotes (Semionotidae).[17] The animals capable of it had been suggested as sharks like Hybodus, actinopterygians and several marine reptiles.[17] Hybodus, being was able to reach nearly 3 meters long and with a dentition suitable to hunt fish, although its stomach contents suggest it is a mostly invertebrate hunter.[17] Actinopterygians like Saurostomus grew up to 2 m long, and have been found with fishes, coeloids and ammonites in its stomach contents, however, not the fishes present on this Spienballen.[17] Marine reptiles included marine crocodiles, such as Platysuchus or Pelagosaurus, associated to the fishes of the Speinballen, although are proven to have eaten gastroliths to improve buoyancy and digestion.[17] Ichthyosaurs, whose diet is among the best studied of the Posidonia Shale, with Dapedium specimens in juvenile stomachs, along with coeloids. Temnodontosaurus, measuring between 8 and 13 metres (26 and 43 ft), would have been able to do such a large Speinballen.[17] Plesiosaurs were disaccredited due to the study of its teeth, that proves a diet based on soft-body prey, such as fishes of the genus Leptolepis and coeloids. Dapedium and Lepidotes, with a heavy and solid squamation can be excluded.[17]
- A specimen of Pachycormus has been found with stomach contents that include hooks similar to the ones found on genera like Clarkeiteuthis.[18]
- Another specimen of Pachycormus macropterus preserves an ammonite inside its gut, likely swallowed by accident and directly responsible for the fish’s death.[19]
- SMNS 51144 (Saurostomus esocinus) was found with Chondrites isp. burrows in the abdominal cavity, what indicates a possible opportunistic scavenger. Other Chondrites isp. includes SMNS 17500 and MHH 1981/25 (Stenopterygius uniter) that can either suggest ichthyosaurs were preserved immediately below one such bioturbation horizon or scavenger association.[8]

- One of the most emblematic finds of the formation its that of a mother Stenopterygius giving birth living young, like the modern dolphins and marine mammals, being born with the tails first. Other specimens have been found with Embryos inside, but with the bones of them scattered, partly beyond the body limits of the mother.[20] There have been various theories about this scenario: the bones of embryos had been deposited before the body of the adult went to the sea floor, covering the embryo bones and implying that the adult would not be the mother of the embryos.[20] Another option is that a pregnant ichthyosaur on its last moments sank to the bottom and may have struggled for life, given untimely birth to some of the foetuses.[20] Other option follows the presence of foetus bones outside the mother body, where a dead female sank to the bottom, with the water warm enough, helping the putrefaction gases to start to develop while the hydrostatic pressure was too high to be prevented by the body. Scavengers must have started eating from the dead body, until the chamber retaining the pressure was to thin and exploded.[20] These theories where however contested after the study, where it was criticised the absence of the presence of the bottom-current activity in the epicontinental sea covering Central Europe during the Toarcian, pointing that the mother carcass should have been translated after it sank to the bottom floor, probably exploding or expelling its embryos first, that would be transported along.[21]
- Specimen SMNS 53363 (Eurhinosaurus?) from Aichelberg was found with two encrusted large oysters (Liostrea) on the right pterygoid, considered to be part of a reef stage over bones.[8]
- SMNS 80234 (Stenopterygius quadriscissus) represents another female with embryos, yet also shows ribs broken perimortem that can be either of intraspecific aggression or a predation attempt. This specimen has several taxa associated: ammonite aptychi and two ophiuroids (Sinosura brodiei) and a articulated echinoid (Diademopsis crinifera), indicating a short-lived deadfall community.[8]
- SMNS 81841 (Stenopterygius quadriscissus) represents one of the most clear examples of deadfall communities described in the formation: the skeleton is associated with serpulids surrounded by a mass of disarticulated ophiuroid remains, indeterminate echinoid tests, an isolated crinoid ossicle, the byssate bivalve Oxytoma inaequivalvis, the pectinid Propeamussium pumilus, Eopecten strionatis, Plagiostoma sp. , Meleagrinella sp., "Cucullaea" muensteri, with the genera Parainoceramya dubia and Liostrea associated with the carcass.[22] As many of this bivalves shown overgrowth likely the community persisted for some time.[22] Fossil traces of Gastrochaenolites isp. attributed to mechanical bivalve borers are abundant implicating prolonged exposure of the skeleton on the seafloor.[22]
- SMNS 81719 (Stenopterygius uniter) includes Liostrea, Propeamussium pumilus, Plagiostoma sp. and Parainoceramya dubia, with other invertebrates found (?) not being part of the deadfall community, such as several ammonites and Parainoceramya valves stratigraphically below the specimen.[8] This specimen includes also traces of scavenging activity, possibly by crustaceans.[8]
- SMNS 80113, (Stenopterygius triscissus) was found populated by Parainoceramya, a specimen of Eopecten strionatis and an unexpected specimen of the small infaunal lucinid Mesomiltha pumila, equivocal evidence for the sulfophilic stage.[8]
- Local ichthyosaur soft tissues include skin enough well preserved to infer coloration and appearance on the living animal, as well evidence for homeothermy and crypsis.[23]
Microbial activity
Non-fenestrate Stromatolite crusts formed in Aphotic deep-water environments during intervals of very low sedimentation are recovered in places such as Teufelsgraben, Hetzles.[24] The Stromatolites of this region have evidence of live on a deeper shelf environment with a quietwater deposit which suffered repeated phases of stagnant bottom waters, where a depth water habitat developed, probably at more than 100 meters depth.[24] There is a thin, southern widespread Stromatolite crust on the Top of the Sachrang Formation, called "Wittelshofener Bank", that has made rethink the depth of the major southern basin of the formation, where with the absence of phototrophic calcareous benthic organisms (probably due to the lack of light), shows the depth character of the basin.[24] On the "Wittelshofener Bank" there is also the only occurrence of Ooids, presumably formed in the same deep-water environment.[24]
Color key
|
Notes Uncertain or tentative taxa are in small text; |
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Frutexites[24] |
|
|
Possible traces of Microbial Activity |
Probably related with Archaea activity.[24] Although Frutexites is a cryptic microfossil and an important element of many deep water stromatolites, with an Inorganic origin proposed, where are interpreted as dendritic shrubs to purely inorganic growth of Aragonitic crystals, but also resemble shrubs of the cyanobacteria Angulocellularia.[24] On the Posidonia a cryptoendopelitic mode of life is assumed, being only possible for Heterotrophic bacteria or Fungi.[24] As seen on the Stromatolites of the Posidonia, Frutexites acted mainly as a dweller or secondary binder of the deep-water stromatolites, not as their major constructor.[24] |
|
Cyanobacteria
| Genus | Species | Location | Material | Notes |
|---|---|---|---|---|
|
|
Crypt laminites |
A Cyanobacteria, member of the family Oscillatoriales. The only difference compared to the Paleozoic forms is the much thinner cellular wall of the specimens described, which otherwise seems to be always thicker, and the more irregular shape of the individual tubes. Girvanella is almost rock-forming in the Lower and Upper levels, and is very common, but can only rarely be detected in the bituminous clay marl slate due to conservation reasons.[25] The Girvanella specimens recovered on the Posidonienschiefer can come from Diatomeas rather than Cyanobacteria, but its assignation is rather controversial.[25] | |
Rhizaria
Foraminifera
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Shells |
A benthonic Foraminiferan, type member of the family Ammodiscinae inside Ammodiscina. |
||
|
Annulina[26] |
|
|
Shells |
A benthonic Foraminiferan, member of Psammosphaerinae inside the family Psammosphaeridae. |
|
|
|
Shells |
A benthonic Foraminiferan, member of Vaginulinidae inside the family Vaginulinida (Lagenina). An extant genus. Its shell resemble a mixture between an ammonite conch, due to having a lower spiral, and a mussel. |
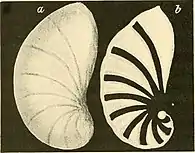 Drawing of an Astacolus shell | |
|
Citharina[26] |
|
|
Shells |
A benthonic Foraminiferan, member of Vaginulinidae inside the family Vaginulinida (Lagenina). |
|
|
Cornuspira[25] |
|
|
Shells |
A benthonic Foraminiferan, type member of Cornuspiridae inside the family Cornuspirida (Lagenina). Round-spiral shell morphology |
|
|
Cyclogyra[26] |
|
|
Shells |
A benthonic Foraminiferan, member of the family Cornuspirinae inside Cornuspiridae. |
|
|
|
Shells |
A benthonic Foraminiferan, member of Nodosariidae inside the family Nodosariacea (Lagenina). Dentalina is an extant genus, with an elongated shell, that resemble a small worm. |
||
|
Flabellinella[25] |
|
|
Shells |
A benthonic Foraminiferan, member of Vaginulinidae inside the family Vaginulinida (Lagenina). Its conch has a Myriapod-like segmented built. |
|
|
Frondicularia[26] |
|
|
Shells |
A benthonic Foraminiferan, type member of Frondiculariinae inside the family Nodosariidae (Lagenina). |
|
|
Glomospira[26] |
|
|
Shells |
A benthonic Foraminiferan, member of the family Usbekistaniinae inside Ammodiscidae. |
|
|
Ichthyolaria[25] |
|
|
Shells |
A benthonic Foraminiferan, type member of Ichthyolariidae inside the family Lagenina. Another genus with a Myriapod-like segmented built. |
|
|
Involutina[26] |
|
|
Shells |
A benthonic Foraminiferan, member of the family Involutinidae inside Involutinae. |
|
|
|
Shells |
A benthonic Foraminiferan, member of Vaginulinidae inside the family Vaginulinida (Lagenina). |
||
|
|
Shells |
A benthonic Foraminiferan, type member of Lingulininae inside the family Nodosariidae (Lagenina). |
||
|
|
Shells |
A benthonic Foraminiferan, member of Marginulininae inside the family Vaginulinida (Lagenina). |
||
|
Nodosaria[26] |
|
|
Shells |
A benthonic Foraminiferan, member of Nodosariidae inside the family Nodosariacea (Lagenina). |
|
|
|
Shells |
A benthonic Foraminiferan, member of Vaginulinidae inside the family Vaginulinida (Lagenina). |
||
|
|
Shells |
A benthonic Foraminiferan, member of Nodosariidae inside the family Nodosariacea (Lagenina). |
||
|
Reinholdella[25] |
|
|
Shells |
A benthonic Foraminiferan, member of Ceratobuliminidae inside the family Robertinida. It resembles a small ammonite. |
|
|
Saracenaria[27] |
|
|
Shells |
A benthonic Foraminiferan, member of Lenticulininae inside the family Vaginulinida (Lagenina). |
|
|
Spiroplectamina[26] |
|
|
Shells |
A benthonic Foraminiferan, member of the family Spiroplectammininae inside Spiroplectamminidae. |
|
|
Trocholina[26] |
|
|
Shells |
A benthonic Foraminiferan, member of the family Involutinidae inside Involutinae. |
|
|
|
Shells |
A benthonic Foraminiferan, type member of Vaginulinidae inside the family Vaginulinida (Lagenina). |
||
Dinoflagellata
Dinoflagellate cysts
The evolutionary burst of the Toarcian Dinoflajellates led the first appearance and rapid radiation of the Phallocystaceae (Susainium, Parvocysta, Phallocysta, Moesiodinium and related forms).[28] This occurred at the time of a widespread Lower Toarcian bituminous anoxia-derived shale of the Posidonienschiefer Formation. Is recovered on the Posidonienschiefer, Pozzale, Italy, Asturias, Spain, Bornholm, Denmark, the Lusitanian Basin of Portugal, the Jet Rock Formation in Yorkshire and to the "Schistes Carton" in northern France. Whether there is a causal connection in this co-occurrence of Phallocystaceae and bituminous facies is a problem still to be resolved. This family has its acme in diversity and quantity in the latest Toarcian and became less important in the Aalenian.[28]
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Cysts |
A Dinoflagellate cyst from the family Apodiniaceae. An Ectoparasitic dinoflagellate, whose hosts are normally Tunicates |
||
|
Argentiella[29] |
|
|
Cysts |
A Dinoflagellate cyst from the family Scriniocassiaceae. |
|
|
Balechiodinium[29] |
|
|
Cysts |
A Dinoflagellate cyst from the family Scriniocassiaceae. |
|
|
Comparodinium[30] |
|
|
Cysts |
A Dinoflagellate cyst from the family Comparodiniaceae. |
|
|
Eyachia[29] |
|
|
Cysts |
A Dinoflagellate cyst from the family Scriniocassiaceae. |
|
|
Luehndea[31] |
|
|
Cysts |
A Dinoflagellate cyst, type member of Luehndeoideae. Luehndea spinosa is common on the medium layers of the lower Sachrang Formation, while restricted to some areas on the Lias delta.[31] |
|
|
Mancodinium[31] |
|
|
Cysts |
A Dinoflagellate cyst, type member of Mancodiniaceae. Dominant genera on some layers of the Lias Delta Stage.[31] |
|
|
Moesiodinium[29] |
|
|
Cysts |
A Dinoflagellate cyst from the family Heterocapsaceae. |
|
|
Morgenrothia[29] |
|
|
Cysts |
A Dinoflagellate cyst from the family Heterocapsaceae. |
|
|
Nannoceratopsis[31] |
|
|
Cysts |
A Dinoflagellate cyst, member of Dinophyceae of the family Nannoceratopsiaceae. On the Lias Epsylon Interval (Lowermost Toarcian), most of the assemblages are dominated by Nannoceratopsis gracilis. Nannoceratopsis senex becomes highly abundant until the uppermost Tenuicostatum.[31] |
|
|
Scriniocassis[29] |
|
|
Cysts |
A Dinoflagellate cyst from the family Scriniocassiaceae. |
|
|
|
Cysts |
A Dinoflagellate cyst from the family Gonyaulacaceae. |
||
|
Susadinium[29] |
|
|
Cysts |
A Dinoflagellate cyst from the family Heterocapsaceae. |
|
|
Parvocysta[29] |
|
|
Cysts |
A Dinoflagellate cyst from the family Heterocapsaceae. |
|
|
|
Cysts |
A Dinoflagellate cyst from the family Phallocysteae. |
||
|
Valvaeodinium[31] |
|
|
Cysts |
A Dinoflagellate cyst from the family Comparodiniaceae. |
|
Algae
Includes abundant variety of algae, such as the genus of colonial Green algae Botryococcus, or the unicellular algal bodies Tasmanites, and other small examples. Algae are a good reference for changes on the oxygen conditions along the Toarcian.[34]
Algae Acritarchs
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Cymatiosphaeropsis[35] |
|
|
Cysts |
An Acritarch probably from Algal origin. Related to open shelf deposits |
|
|
Micrhystridium[31] |
|
|
Cysts |
An Acritarch probably from Algal origin. Its fossils indicate nearshore or estuarine to shallow lagoon and/or slightly brackish-water environments. It is the dominant on the nearshore sections. |
|
|
Pterosphaeridia[35] |
|
|
Cysts |
An Acritarch probably from Algal origin. Related to open shelf deposits |
|
|
Veryhachium[31] |
|
|
Cysts |
An Acritarch probably from Algal origin. Veryhachium fossils represent open marine and transgressive conditions. It has high presence on most of the samples studied from the Sachrang Formation, being nearly the 50% of the Acritarch fraction on some locations. |
|
Haptophyta
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Calcareous Skeletons |
Type member of the family Biscutaceae inside Parhabdolithaceae. |
||
|
|
Calcareous Skeletons |
A member of the family Watznaueriaceae inside Watznaueriales. |
||
|
|
Calcareous Skeletons |
Member of the family Calyculaceae inside Parhabdolithaceae. |
||
|
|
Calcareous Skeletons |
A member of the family Chiastozygaceae inside Eiffellithales. |
||
|
Diductius[38] |
|
|
Calcareous Skeletons |
Member of the family Parhabdolithaceae inside Stephanolithiales. |
|
|
|
Calcareous Skeletons |
Member of the family Biscutaceae inside Parhabdolithaceae. |
||
|
|
Calcareous Skeletons |
A member of the family Watznaueriaceae inside Watznaueriales. |
||
|
|
Calcareous Skeletons |
A member of the family Parhabdolithaceae inside Stephanolithiales. Shore deposits genus. The abundance drop of M. jansae further characterise the T-OAE perturbation, where becomes the dominant Genus on most of the Saxony Basin. |
||
|
Diductius[38] |
|
|
Calcareous Skeletons |
Member of the family Parhabdolithaceae inside Stephanolithiales. |
|
|
|
Calcareous Skeletons |
Incertade Sedis |
||
|
|
Calcareous Skeletons |
Type member of the family Schizosphaerellaceae inside Parhabdolithaceae. Towards the Pliensbachian-Toarcian extincion this genus gets a decrease in abundance and size that shows the change and biotic crisis. |
||
|
|
Calcareous Skeletons |
Member of the family Biscutaceae inside Podorhabdales. |
||
|
Sollasites[37] |
|
|
Calcareous Skeletons |
Member of the family Biscutaceae inside Podorhabdales. |
|
|
|
Calcareous Skeletons |
A member of the family Chiastozygaceae inside Eiffellithales. |
||
Chlorophyta
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Cysts |
Type member of the family Botryococcaceae inside Trebouxiales. Freshwater or Deltaic Genus |
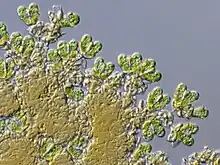 Modern Botryococcus | |
|
|
Cysts |
A member of Prasinophyceae. A genus common on green clays and other upper strata on the formation. |
||
|
|
Cysts |
A member of the family Pyramimonadales inside Prasinophyceae. Basinal deposits genus |
||
|
|
Cysts |
A member of Gonyaulacaceae inside Dinophyceae. |
||
|
Granodiscus[35] |
|
|
Cysts |
A member of the Prasinophyceae. Basinal deposits genus |
|
|
Halosphaeropsis[31] |
|
|
Cysts |
A member of the family Halosphaeraceae inside Chlorodendrales. Basinal deposits genus |
|
|
Lancettopsis[35] |
|
|
Cysts |
A member of the Prasinophyceae. Basinal deposits genus |
|
|
|
Cysts |
A member of the Prasinophyceae. Basinal deposits genus |
||
|
Nostocopsis[35] |
|
|
Cysts |
A member of the Prasinophyceae. Basinal deposits genus |
|
|
|
Cysts |
A member of Peridiniaceae inside Dinophyceae. |
||
|
|
Cysts |
A member of Prasinophyceae. It is the main genus present on silt and sand horizons, trending to be absent on shale layers. |
||
|
|
Cysts |
A member of Dinophyceae. |
||
|
|
Cysts |
A member of Prasinophyceae. A genus common on green clays and other upper strata on the formation. |
||
|
Tytthodiscus[35] |
|
|
Cysts |
A member of the Prasinophyceae. Basinal deposits genus |
|
Fungi
Fungal Spores, hypae and undeterminated remains are a rare element of the otherwise openmarine deposits of the Posidonienschiefer formation, but where recovered at Dormettingen.[43] This fungal remains are composed mostly by indeterminate spores and indicate oxygenated environments and suitable transportation by rivers.[43]
Incertae sedis
| Genus | Species | Location | Stratigraphic position | Material | Notes | Images |
|---|---|---|---|---|---|---|
|
Ostracoblabe[44] |
|
|
|
Fungal patches in ammonite shells and belemnite rostra |
A Marine parasitoid Fungus of uncertain relationships, linked with shells of marine invertebrates. Extant Ostracoblabe implexa is usually found associted with Bivalve shells as an external parasiotid. Beyond this genus, other fungal remains include indeterminate endolithic fungi linked with microbial mats |
|
Ichnofossils
The major ichnological analyses of the Posidonian Shale come from Dotternhausen/Dormettingen, where the ichnogenus Phymatoderma formed the so-called Tafelfleins and Seegrasschiefer.[45] The Tafelflein bed was deposited under anoxic bottom and pore water, where a recover of oxygen allow the Phymatoderma-producers return.[45] The two organic-rich layers (Tafelfleins and Seegrasschiefer) are characterized by the dense occurrence of trace fossils such as Chondrites and Phymatoderma, done episodically due to the fall of the oxygen levels.[45] The Coeval more nearshore Swiss deposits referred Posidonian Shale (Rietheim Member) hosted similar trace fossils asthose recovered on SW Germany.[45] Tougth this setting apparently evolved faster to more oxic-to-dysoxic bottom waters.[45] At Unken, laminated deposits of red limestone suggest well oxygenated active waters (as lack shale), where high amounts of Chondrites are found.[37]
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Burrowing and track ichnofossils |
Burrow-like ichnofossils. Interpreted as the feeding burrow of a sediment-ingesting animal.[48] A more recent study has find that Scoloplos armiger and Heteromastus filiformis, occurring in the German Wadden Sea in the lower parts of tidal flats, make burrows that are homonymous with numerous trace fossils of the ichnogenus.[49] Chondrites burrows from Holzmaden are mostly filled with granular Calcite crystals, clay minerals, and rare framboidal Pyrite. The local dysoxic seawater is reflected on the palaeoredox conditions, relating framboidal pyrite and biogenic processes. Chondrites appear to be capable of colonizing environments characterized by oxygen levels well below levels needed for survival of other animals, being considered an "extremotolerant" ichnotaxon.[50] |
 Illustration of Chondrites bollensis | |
|
|
Borings on bones |
Borings on Ichthyosaur bones. Reminiscent of Gastrochaenolites and attributed to mechanical bivalve borers |
 Example of Gastrochaenolites fossil | |
|
|
Burrowing and track ichnofossils. |
Burrow-like ichnofossils. It is controversial, since is considered a strictly a junior synonym of Palaeophycus.[52] |
 Example of Planolites fossil | |
|
Phymatoderma[53] |
|
|
Burrowing and track ichnofossils. |
Burrow-like ichnofossils. It consists commonly on a subhorizontal branching burrow system consisting of radiating tunnels filled with fecal pellets.[53] It has been interpreted as a product made by an Endobenthic deposit-feeding animal, specially a Fodinichnia, burrows produced by benthonic subsurface food-mining activity, as is proved by the tunnels and pelletal infill.[53] The study of the Fecal Pellets has revelated that the maker of this ichnogenus was an epicontinental shelf setting non-selective deposit feeder, ingesting particles on the sediment surface without selection. A mode of feeding common on aquatic Benthos, reported on modern animals such as Spionid Polychaete worms, tropical Holothurians and Spatangoid urchins.[54] |
|
|
|
Burrowing and track ichnofossils |
U-shaped tubes with sinuous, bifurcating or planispiral spreite, that can be related to Crustaceans, Annelids and Fishes, being both Domichnia and/or fodinichnia. |
 Specimens | |
|
|
Burrowing and track ichnofossils |
Burrow-like ichnofossils, that can be related to Crustaceans, Annelids and Fishes.[55] The presence of this burrows changed along the different depositional layers, interpreted as result of relative magnitudes and durations of a series oxygenation events.[55] Increased Oxygen conditions eventually led to a level that permitted both the survival of larger Chondrites and Thalassinoides producing organisms, as well the depth of the Burrow-like structures.[55] The changes on the layers are detailed enough to know that oxygenation-change events duration was sufficient to allowe the migration and establishment of trace-producing organisms, establishing an "equilibrium" with bottom-water oxygen conditions.[55] |
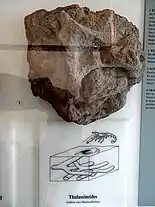 Thalassinoides found on the Sachrang Formation | |
|
|
Burrowing and track ichnofossils. |
Burrow-like ichnofossils. It has been related to Echiuran annelids, but also from moving and feeding polychaete worms.[57] |
 Example of Zoophycos fossil | |
Invertebrata
Porifera
In the non-bituminous facies located on Obereggenen im Breisgau (Shore of the Black Forest High), especially the lower semicelatum subzone, pyritized individual needles of silica sponges (Demospongiae and Hexactinellida) are found, rarely on pelagic layers to very often on the low depth marine deposits.[25] They are usually associated with radiolarian stone cores. In Dusslingen and Reutlingen, these sponge needles could be barytized in phosphorites of the Haskerense subzone and are much more common here than in any other zone of the Lower Toarcian. These needles are absent in the bituminous horizons of the entire Lower Toarcian.[25] Increased amounts of Sponge needles (dominated by Hexactinellida) are also found on the arenaceous facies of the nearshore unit that is the Unken member, being the only section if its region hosts them, probably due to be an active and well oxygenated bottom.[37] The location of this member as a possible bay on the south of the vindelician land probably allow to the development of more pre-Toarcian AOE conditions, hence the presence of biota otherwise rare on bituminous layers.[37]
Annelida
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Multiple Specimens. |
A sessile, marine annelid tube worm of the family Serpulidae. Its affinities with the genus Serpula are controversial, since the genus is known mostly since Cretaceous strata. Although there are other fossils assigned to the genus on same age deposits of France. Presumably these specimens have fallen from their growth areas.[25] |
 Head of a modern Serpula vermicularis | |
|
Toarcihalla[25] |
|
|
|
A polychaete of the family Dorvilleidae inside Eunicida. It was considered as part of the genus Halla. Eunicidan species with prionognath jaws, absent on Bituminous layers |
|
Brachiopoda
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Shells |
A pennospiriferinid rhynchonellatan.[58] |
||
|
|
Shells |
A Lingulidae rhynchonellatan. Associations of bioturbation infauna are dominated on certain sections by Palaeonucula/Lingula agrupations, developed under longer-term oxygenated conditions within the substrate and bottom waters.[58] |
||
|
Orbiculoidea[44] |
|
|
Shells |
A Discinidae rhynchonellatan. This genus was found had a planktotrophic larval stage, that adapted while growing to the local redox boundary, when this fluctuated near the sediment–water interface and oxygen availability prevailed, allowing benthic colonization. Is found on associations with Grammatodon and Pseudomytiloides.[58] |
|
|
|
Shells |
A Rhynchonellidae rhynchonellatan. Found associated with Plicatula on long-term well-oxygenated conditions within the substrate and bottom waters.[58] |
||
|
|
Shells |
A Spiriferinidae rhynchonellatan. |
||
|
Waldheimia[44] |
|
|
Shells |
||
Bivalvia
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Antiquilima[44] |
|
Dotternhausen |
Shells |
||
|
Bositra[59] |
|
All the Formation |
20.000 specimens/m2 |
A posidoniid ostreoidan. It is the type fossil of the Sachrang Formation. Originally it was named "Posidonia bronni", thought to be a new genus, and the strata was denominated the Posidonia layers after it. Years later it turned out to be a junior synonym of Bositra, and thus, it was reassigned. However, the name of the layers was retained. The habitat and mode of life of Bositra has been debated for more than a century. There have been different interpretations, such as a pseudoplanktonic organism,[60] a benthic organism related to open marine floor, where it was the main inhabitant of the basinal settings, and a hybrid mode, where it has a life cycle with holopelagic reproduction controlled by the change on Oxygen levels, and even a chemosymbiotic lifestile, related to the large crinoid rafts, being the main "Safe conduct" to evade anoxic events. All the opinions along the years led to a large study in 1998, where the size/frequency distribution, the density of growth thanks to the lines related to the shell size and the position of the redox boundary by total organic carbon diagrams has revealed that Bositra probably had a benthic mode of life.[61] |
 Thousands of specimens in one matrix |
|
Camptonectes[62] |
|
|
Shells |
A pectinoid scallop. The presence of this genus along endo- and epibenthic bivalves, which are absent farther up the section, suggest a delayed overstepping of anoxic bottom waters on the Altdorf High.[62] |
|
|
|
Shells |
A pectinoid scallop. |
 Single specimen | |
|
|
Shells |
A cucullaeid clam. |
||
|
|
Shells |
A pectinoid scallop. |
||
|
|
Shells |
|||
|
|
Shells |
A bakevelliid mud oyster. |
||
|
Goniomya[4] |
|
|
Shells |
||
|
Grammatodon[44] |
|
|
Shells |
A Grammatodontinae clam. This Genus had a lecithotrophic and planktotrophic larval development.[58] |
|
|
|
Shells |
 Various specimens | ||
|
|
Shells |
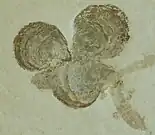 Various specimens | ||
|
|
Shells |
An oxytomid scallop. |
 Colony of specimens | |
|
|
Shells |
|||
|
Mytiloides[66] |
|
|
Shells |
An inoceramid clam. |
|
|
Nicaniella[4] |
|
|
Shells |
||
|
|
Shells |
An oxytomid scallop. |
||
|
Palaeonucula[4] |
|
|
Shells |
||
|
Parainoceramya[66] |
|
|
Shells |
An inoceramid clam. |
 Thousands of specimens on a single rock |
|
|
Shells |
|||
|
|
Shells |
 Plagiostoma giganteum, specimen multiview | ||
|
Pleuromya[4] |
|
|
Shells |
A Pleuromyid clam |
|
|
|
Shells |
A plicatulid mud scallop. |
||
|
Praearctotis[63] |
|
|
Shells |
An oxytomid scallop. Found mostly on the "Dactylioceras-Monotis-Bank", a deposit derived from large scale tectonic events on the Bohemian coastline |
|
|
|
Shells |
A propeamussiid mud scallop. |
 Various specimens on the same rock | |
|
Pseudolimea[44] |
|
|
Shells |
||
|
|
Shells |
An inoceramid clam. Being the second most common genera of Bivalve on the Formation, it had been object to several studies to find its ecological niche, like Bositra. Several opinions include a pseudoplanktonic-only organism, able to live in open sea, or a benthonic-only organism. On the 1998 evaluation with Bositra, was found that probably has a benthic early life that translated to a faculatively pseudoplanktonic mode of adult life.[61] |
 Single specimen | |
|
|
Shells |
A Pteriidaeoid wing-oyster. |
||
|
|
Shells |
A Clam, type member of the family Solemyidae inside Solemyida. |
 Single specimen | |
|
Steinmannia[44] |
|
|
Shells |
A "posidoniid" ostreoidan. Another Genera mistaken with "Posidonia bronni". |
 Various specimens in one matrix |
|
|
Shells |
A mactromyid clam. |
||
Gastropoda
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Amberleya[44] |
|
|
Shells |
A Eucyclidae sea Snail. |
|
|
|
|
A Coelodiscidae sea Snail. Is the oldest known holoplanktonic gastropod, thanks to a bilateral symmetrical shells as an adaption to active swimming. Also the most common of the sea snails of the Formation, it is also one of the most varied in size terms, with some of the biggest specimens of snail from the Lower Toarcian know.[68] It has been related to large floating driftwood as one of the primary settlers.[68] |
||
|
Eucyclus[44] |
|
|
Shells |
A Eucyclidae sea Snail. |
 Specimens |
|
|
Shells |
 Extant Specimen | ||
|
|
Shells |
A Pleurotomariidae sea Snail. |
 Specimen | |
|
|
Larval Shells |
A Pterotracheidae sea Slug. Among the oldest pelagic floating Slugs, Pterotrachea liassica had a more extended larval period than modern extant Pterotrachea coronata, because one additional whorl is present.[69] |
 Modern Specimen | |
|
Rhabdocolpus[44] |
|
|
Shells |
A Procerithiidae sea Snail. |
|
|
Rigauxia[44] |
|
|
Shells |
A Snail of uncertain placement. |
|
|
|
Shells |
A Coelodiscidae sea Snail. Possible holoplanktonic gastropod.[68] |
||
|
|
Shells |
A Zygopleuridae sea Snail. |
 Specimens | |
Cephalopoda
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Shells |
|||
|
|
Aptychi |
Ammonite internal moulds of uncertainf affinity. Represent the most diverse Ammonite remains found on all the formation and are usually not identificable to family level |
||
|
|
Phragmocones |
A Megateuthididae Belemnite. Includes some of the Biggest Know Belemnites, with an estimated maximum up to 2 m long in life, although, most specimens would have been rather smaller. |
||
|
Belotheutis[77] |
|
|
Complete Specimens with tissue, complete Arm Crown, ink Sacs and organ Traces |
A Diplobelidae Coleoidean. Some specimens belong to Clarkeiteuthis (=Phragmoteuthis) conocauda, but others are clearly different. |
|
|
Brodieia[44] |
|
|
Pyritized Fragments |
A Phymatoceratidae Ammonite. |
|
|
|
Shells |
|||
|
|
Shells |
|||
|
Catulloceras[78] |
|
|
Shells |
||
|
|
Shells |
A Nautilidae Nautilidan. Two referred specimens, identified as Nautilus spp. from Holzmaden where found encrusted with Serpulids and Bryozoans.[80] |
 Nautilidae shell from Banz, probably Cenoceras | |
|
Chitinobelus[81] |
|
|
Phragmocones |
A Belemnotheutidae Belemnite. Chitinobelus rostrum was composed of aragonite with organic material, while normal Belemnites had calcite. Has been suggested this rostrum was calcitic. |
|
|
|
Phragmocones |
A Belemnotheutidae Belemnite. |
||
|
|
|
A Diplobelidae Coleoidean. It has been found adult individuals of Clarkeiteuthis which caught small teleost fish of the species Leptolepis bronni. Further indirect evidence for the hunting behaviour comes from their body orientation in the water during life.[9] |
 Clarkeiteuthis Holzmaden specimen | |
|
|
Shells |
| ||
|
|
Shells |
Type Coeloceratidae Ammonite. |
||
|
|
Shells |
A Dactylioceratidae Ammonite. Is common on the bituminous marls (incorrectly designated as "Wilder Schiefer") of the Altdorf High. |
||
|
|
Aptychi |
Ammonite internal moulds of uncertainf affinity. Represent the most diverse Ammonite remains found on all the formation and are usually not identificable to family level |
 Cornaptychus lythensis | |
|
Cotteswoldia[78] |
|
|
Shells |
||
|
|
Shells |
 Dactyliocerascommune on Holzmaden | ||
|
|
Phragmocones |
A Megateuthididae Belemnite. |
||
|
|
Shells |
A Phymatoceratidae Ammonite. |
||
|
|
Shells |
|||
|
|
Shells |
|||
|
|
Shells |
A Hildoceratidae ammonite. The co-occurrence on Altdorf of boreal (Pseudolioceras) and Tethyan faunal elements (Frechiella) is striking, suggesting clear connection with both regions.[87] |
||
|
Furloceras[71] |
|
|
Shells |
A Phymatoceratidae Ammonite. |
|
|
|
|
A Geopeltidae Loligosepiidan (Vampyromorpha). Related to the modern Vampyroteuthis infernalis. Gladius with weakly arcuated hyperbolar zones. |
 Geopeltis specimen | |
|
Geotheutis[89] |
|
|
|
A possible primigenial Cuttlefish. Is one of the most important fossils of Cephalopods on the Sachrang Formation, due to be one of the Earliest examples of Pigments found on any species, also one of the first historically.[91] The pigments are preserved on various specimens with Eumelanin related to its ink sacs and include even phosphatized musculature.[10] |
|
|
|
Shells |
 Harpoceras specimen | ||
|
|
Shells |
A Phymatoceratidae Ammonite. |
||
|
|
Shells |
|||
|
Shells |
 Hildoceras specimen | |||
|
Hudlestonia[78] |
|
|
Shells |
||
|
|
|
A Loligosepiidae Loligosepiidan (Vampyromorpha). Related to the modern Vampyroteuthis infernalis. Gladii of Loligosepia can be distinguished from Jeletzkyteuthis by the transition lateral field/hyperbolar zone. A specimen of this genus found on Ohmden has appeared predating a Parabelopeltis.[11] The association of this 2 genera shows the predatory behaviour of this group when lived on Epicontinental seas, being rather different than extant Vampyromorphs.[11] |
||
|
Kedonoceras[86] |
|
|
Shells |
||
|
Lioteuthis[93] |
|
|
Single Specimen with tissue |
Type member of the Lioteuthididae inside Vampyromorphida. The taxonomic position of Lioteuthis is uncertain, although the wings reaching the proximal gladius section and the smooth median field suggest affinity to the Prototeuthididae[93] |
|
|
|
Shells |
|||
|
Loligosepia[94] |
|
|
|
A Loligosepiidae Loligosepiidan (Vampyromorpha).[95] The Loligosepiidae is believed to be ancestral to the Recent vampire squid, Vampyroteuthis infernalis.[88] Hooklets in the food residues in the posterior mantle indicate that Loligosepia preyed upon belemnites.[94] |
 Loligosepia Holzmaden specimen |
|
|
Shells |
A Lytoceratidae Ammonite. Lytoceras can get quite big, with nearly 50 cm in diameter. |
||
|
|
Shells |
 Mercaticeras specimen | ||
|
|
Phragmocones |
A Passaloteuthididae Belemnite. |
||
|
|
Shells |
|||
|
|
Shells |
|||
|
|
Shells |
|||
|
Odontobelus[77] |
|
|
|
A Diplobelidae Coleoidean. Has been confused with Acrocoelites tripartitus, hence the species name. |
|
|
|
Hooks |
Incertae sedis Belemnites. |
||
|
|
Shells |
|||
|
|
Shells |
|||
|
|
Shells |
|||
|
|
|
A Geopeltidae Loligosepiidan (Vampyromorpha). Related to the modern Vampyroteuthis infernalis. It is distinguished from Geoteuthis and Loligosepia by its median rib: this rib forms a narrow ridge between two narrow grooves. Probably hosted finds similar to modern Vampyroteuthis.[11] |
||
|
Paroniceras[73] |
|
|
Shells |
||
|
|
Partial Specimens with tissue |
A Plesioteuthididae Prototeuthidinan (Vampyromorpha). was originally described as "Geoteuthis" sagittata. |
||
|
|
|
A Passaloteuthididae Belemnite. |
 Passaloteuthis Holzmaden specimen | |
|
|
Shells |
|||
|
|
Shells |
A Phylloceratidae Ammonite. The largest ammonite found in the Posidonienschiefer comes from the Ohmden quarry,and belongs to a Phylloceras heterophyllum with a diameter of 87 cm.[72] |
 Phylloceras restoration | |
|
|
Shells |
|||
|
|
Shells |
A Phymatoceratidae Ammonite. |
||
|
|
Shells |
|||
|
|
Shells |
|||
|
Protogrammoceras[71] |
|
|
Shells |
||
|
Pseudogrammoceras[99] |
|
|
Shells |
||
|
|
Shells |
|||
|
|
Phragmocones |
A Salpingoteuthididae Belemnite. |
Salpingoteuthis specimen | |
|
|
Phragmocones |
A Belemnotheutidae Belemnite. |
||
|
Sueviteuthis[100] |
|
|
|
A Sueviteuthididae Coleoidean. Sueviteuthis had at least six arms with rather simple hooks, similar to the present of the genus Phragmoteuthis. |
|
|
|
|
A Teudopseinae Palaeololiginidaean (Vampyromorphida). |
 Teudopsis Ohmden specimen | |
|
|
Shells |
 Tiltoniceras specimen | ||
|
|
Shell |
|||
|
|
Phragmocones |
A Megateuthididae Belemnite. Includes really large specimens |
 Youngibelus Reconstruction | |
|
|
Shells |
|||
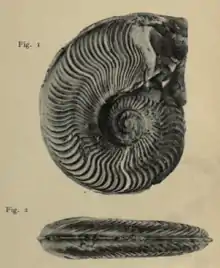
Cycloidea
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Incomplete carapace |
The First Cycloid Arthropod from the Jurassic, from the family Halicynidae inside Cycloidea.[102] Cycloids are a group of maxillopod arthropods that span between the Paleozoic until the latest Cretaceous, probably related to the crustaceans and probably detritivores.[102] |
||
Ostracoda
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Valves |
A Marine Ostracodan of the family Bairdiidae inside Bairdioidea. Abundant and diverse, is found associated with Ammonite shells. |
||
|
Bairdiacypris[103] |
|
|
Valves |
A Marine Ostracodan of the family Bairdiidae inside Bairdioidea. |
|
|
|
Valves |
A Marine Ostracodan of the family Cytherellidae inside Platycopida. |
||
|
|
Valves |
A Marine Ostracodan of the family Cytherellidae inside Platycopida. |
||
|
Eucytherura[103] |
|
|
Valves |
A Marine Ostracodan of the family Cytheruridae inside Podocopida. Is rare and the specimens found are rather incomplete. |
|
|
Hermiella[105] |
|
|
Valves |
A Marine Ostracodan of the family Healdiidae inside Podocopida. This genus is the main reported on the marine facies of the Dobbertin Clay Pit. |
|
|
Infracytheropteron[103] |
|
|
Valves |
A Marine Ostracodan of the family Protostomia. The specimens of this genus are rather fargmentary and of uncertain nature. |
|
|
|
Valves |
A Marine Ostracodan of the family Protocytheridae inside Podocopida. A genus related with Fish fossils and anoxic bottoms. |
||
|
|
Valves |
A marine Ostracodan, member of the family Pontocyprididae inside Podocopida. |
||
|
|
Valves |
A marine Ostracodan, member of the family Macrocyprididae inside Podocopida. |
||
|
|
Valves |
A Marine Ostracodan of the family Bythocytheridae inside Cladocopina. |
||
|
|
Valves |
A marine Ostracodan, member of the family Healdiidae inside Podocopida. Rather abundant on the Toarcian profiles on Europe, this genus has a Mussel-like shape, with a very clean and round morphology. |
||
|
|
Valves |
A Marine Ostracodan of the family Healdiidae inside Podocopida. This genus is the main reported on the marine facies of the Dobbertin Clay Pit. |
||
|
|
Valves |
A Marine Ostracodan of the family Polycopidae inside Cladocopina. Scarce but well preserved specimens. |
||
|
Praeschuleridea[103] |
|
|
Valves |
A Marine Ostracodan of the family Praeschuleridea inside Podocopida. |
|
|
|
Valves |
A Marine Ostracodan of the family Healdiidae inside Podocopida. The genus is rare on the layers. |
||
|
Trachycythere[44] |
|
|
Valves |
A marine Ostracodan, incertae sedis inside Podocopida. |
|
Malacostraca
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
An Aegeridae Decapodan. |
||
|
|
Single complete specimen in late larval stage |
The specimen reported represents the oldest fossil record of an achelatan lobster larva, and the first representative of achelatan lobsters in the Posidonia Shale. Shares similarities with the late Jurassic genus Cancrinos. It is also the first example among crustaceans, which possibly could have lived as part of the plankton.[109] |
 California spiny lobsters are relatives of the Larval specimen from the Sachrang Formation | |
|
|
Partial Specimens. |
A Penaeidae Decapodan. |
 Antrimpos specimen | |
|
|
|
An Erymidae Decapodan. |
||
|
|
Complete Specimens |
Type genus of the Erymidae Decapodan family. Originally, was named Glyphea amalthei,informally used by Quenstedt and housed on the Museum Naturkunde in Württemberg. A series of posterior revisions probe it was a different genus.[113] |
 Eryma specimen | |
|
Gabaleryon[114] |
|
|
|
A Coleiidae Decapodan. Was confussed with Proeryon hartmanni specimens. Although these finds point to some potential for exceptional preservation, the Pfronten locality was never systematically sampled.[114] It was assigned to the genus Gabaleryon the basis of its ovoid carapace, a transverse groove cutting the axial carina on pleonal terga and an uropodal exopod with curved diaeresis.[114] It shares similarities with Gabaleryon garassinoi and Gabaleryon moorei.[114] A species from Gomaringen is the first know with preserved ommatidia.[115] |
|
|
Glypheopsis[116] |
|
|
Isolated Chelae |
A Decapodan of the family Glypheidae. |
|
|
|
Complete Specimens |
A Decapodan of the family Mecochiridae. |
||
|
|
Partial Specimens. |
A Penaeidae Decapodan. |
||
|
|
Partial Specimens. |
An Erymidae Decapodan. |
 Palaeastacus specimen | |
|
|
Single Specimen inside an Ammonite Shell. |
An hermit crab of the family Paguridae. This specimens where found inside an Ammonite shell, probably looking to evade anoxic conditions or predators. |
||
|
|
Partial Specimens |
A Spiny Lobster of the family Palinuridae |
||
|
|
|
A Coleiidae Decapodan. The second largest Decapodan from the formation, P. giganteus is a species reaching a larger size than most other polychelidans, with up to 15 cm. On the Sachrang Formation there is the most abundant variety of species from the genus, ranging from different sizes and morphologies, that indicate different habitat & feeding adaptations on the genus. Some like P. hartmani show less adaptations to hunt for small nectobenthic preys than other relatives, being abundant on Oyster-filled waters. There is a relatively abundance of the genus in deep-water settings from the Toarcian onward.[121] |
 Proeryon giganteus | |
|
Stenodactylina[122] |
|
|
Single Chela |
An Erymidae Decapodan. It was erroneously reported from the Late Toarcian, as was found on bituminous limestone on the Unterer Stein it is clear its provenance from the Sachrang Formation. |
|
|
|
Single Incomplete Specimen |
A Stomatopoda Malacostracan. Its affinities haven't been tested. |
 A female Odontodactylus scyllarus mantis shrimp, maybe related to the Posidonia Specimen | |
|
Tonneleryon[111] |
|
|
Associated Partial & Complete Specimens |
A gregarious Polychelidan Lobster. Specimens of Tonneleryon schweigerti where recovered generally in cluster of several individuals, a characteristic unique to this species on the whole Polychelida group.[111] The specimens in these accumulations are of similar size, lacking characters of exuviae such as a split median line or disjunction of carapace and first pleonite. Due to that and the disposition of the specimens probably represent a mass-mortality assemblage and suggest this species was gregarious.[111] |
|
| Uncina[123] |
|
|
|
An Astacidea Decapodan of the family Uncinidae. Reaching large sizes of almost half a meter (39–47 cm), Uncina Posidoniae is among the largest known Jurassic Crustaceans. Uncina posidoniae is also the largest representative of the genus Uncina.[123] This large crustacean has been found associated with Ammonite and Bivalve filled Bentos, where probably hunted different kinds of prey.[123] Its large claws would have been perfect to hunt small invertebrates and vertebrates.[123] |
 The Largest complete Uncina posidoniae specimen, with 44 cm long. |
Thoracica
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Toarcolepas[15] |
|
|
Numerous disarticulated individuals, associated with fossil wood.[15] |
A phosphatic-shelled Cirripede of the family Eolepadidae.[15] Toarcolepas is provisionally interpreted as the oldest epiplanktonic cirripede known, and is thought to have lived attached to floating driftwood.[15] |
 Modern genus Lepas is among the most related taxa to Toarcolepas |
Arachnida
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Single Incomplete Specimen. |
The type genus of the family Liassoscorpionididae, probably related to Mesophonoidea. Being the only Jurassic scorpion known, there is no evidence that L. schmidti was aquatic (which was suggested in the past) and in the absence of further, better preserved material it should be excluded from future considerations of broad patterns of scorpion evolution.[125] |
||
Insecta
Insects are a common terrestrial animals that were probably washed into the sea due to monsoon conditions present on the Sachrang Formation.[126]
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Agmatozoon[127] |
|
|
Incomplete Specimens |
Uncertain Affinity Insects, whose remains are scarce to allow an assignation to a concrete family. Also Includes insects with characters never seen in other families and genera. |
|
|
Campeulites[127] |
|
| |||
|
Tomeferusa[127] |
|
| |||
|
Trimerocephalium[127] |
|
| |||
|
Dimeretes[127] |
|
| |||
|
Elasmoscolex[127] |
|
| |||
|
Epimetrophora[127] |
|
| |||
|
Oocephalina[127] |
|
| |||
|
Platycorion[127] |
|
| |||
|
Griphoconion[127] |
|
| |||
|
Cricolia[127] |
|
| |||
|
|
|
Gryllones (Extinct clade of Basal Insects) of the family Geinitziidae. |
||
|
Roemerula[128] |
|
| |||
|
Dorniella[127] |
|
|
Isolated Wings |
An Eoblattidan of the family Blattogryllidae. |
|
|
|
Specimens |
An Odonatopteran (ancient winged insects) from the family Protomyrmeleontidae. |
||
|
Liassostenophlebia[127] |
|
|
Isolated Wings |
An Indeterminate Family dragonfly. Dragonflies non assiganted to a concrete family due to the incomplete or fragmentary nature of its remains. |
|
|
Syrrhoe[127] |
|
| |||
|
|
|
A dragonfly of the family Heterophlebiidae. Heterophlebia is a relative abundant Genus, present on most of the pits of the Sachrang Formation, even on Holzmaden. Heterothemis is the second most distributed insect on the formation. |
||
|
Plagiophlebia[127] |
|
| |||
|
Heterothemis[127] |
|
| |||
|
|
Isolated Wings |
A dragonfly of the family Liassogomphidae. |
||
|
Proinogomphus[127] |
|
| |||
|
| ||||
|
Elattogomphus[127] |
|
| |||
|
|
|
A dragonfly of the family Sphenophlebiidae. The Kerkhofen specimens of Sphenophlebia pommerana shows an almost complete dragonfly with head and abdomen as well the two pairs of wings that are partially on top of each other, consisting of fore and hind wings.[136] |
||
|
Ensphingophlebia[127] |
|
| |||
|
Mesoepiophlebia[133] |
|
| |||
|
|
Isolated Wings |
A dragonfly of the family Myopophlebiidae. |
||
|
| ||||
|
| ||||
|
Paraplagiophlebia[133] |
|
| |||
|
|
Isolated Wings |
A dragonfly of the family Campterophlebiidae. Campterophlebia is the largest Early Jurassic Insect Know, with a wings size up to 20 cm.[137] |
||
|
Gallodorsettia[138] |
|
| |||
|
Henrotayia[139] |
|
|
|
A dragonfly of the family Henrotayiidae. |
|
|
Liassogrylloides[127] |
|
|
|
grasshoppers of uncertain Placement. |
|
|
Prophilaenites[127] |
|
| |||
|
Protogryllus[127] |
|
|
|
A grasshopper of the family Protogryllidae. |
|
|
Panorpidium[127] |
|
|
|
A grasshopper of the family Elcanidae. |
|
|
|
|
A grasshopper of the family Elcanidae. |
||
|
Acridiopsis[127] |
|
|
|
A short-horned grasshopper of the family Acrididae. |
 Extant member of Acrididae, Acridiopsis was probably similar |
|
|
|
Grasshoppers of the family Locustopsidae. |
||
|
Liadolocusta[127] |
|
| |||
|
|
Isolated Wings |
A stick insect of the family Aerophasmidae. Schesslitziella is one of the described insects found more near the Bohemian Massif, where probably belong most of the terrestrial invertebrate fauna. |
||
|
Chresmodella[127] |
|
| |||
|
Liadoblattina[143] |
|
|
|
A cockroach of the family Raphidiomimidae. |
|
|
| ||||
|
Mesoblattina[143] |
|
|
Isolated Wings |
A cockroach of the family Mesoblattinidae. |
|
|
|
Isolated Wings |
A cockroach of the family Caloblattinidae. |
||
|
|
Isolated Wings |
A cockroach of the family Blattulidae. |
||
|
Elasmoscelidium[127] |
|
|
Isolated Wings |
A Planthopper of uncertain placement. |
|
|
Archijassus[127] |
|
|
Isolated Wings |
A Planthopper of the family Archijassidae. |
|
|
Fulgoridium[127] |
|
|
|
Planthoppers of the family Fulgoridiidae. |
.jpg.webp) Extant member of Fulgoridae, Fulgoridiidae genera where probably similar |
|
Fulgoridulum[127] |
|
| |||
|
Procerofulgoridium[127] |
|
| |||
|
Tetrafulgoria[127] |
|
| |||
|
Metafulgoridium[127] |
|
| |||
|
Productofulgoridium[127] |
|
| |||
|
Margaroptilon[127] |
|
| |||
|
Compactofulgoridium[127] |
|
| |||
|
Procercopis[127] |
|
|
Isolated Wings |
A Froghopper of the family Procercopidae. |
|
|
Megalocoris[127] |
|
|
|
A Shore bug (Saldidae) Of Uncertain Placement. |
|
|
Eurynotis[127] |
|
|
|
A Shore bug (Saldidae) of the family Archegocimicidae. |
 Extant member of Saldidae, Archegocimicidae genera where probably similar |
|
Somatocoris[127] |
|
| |||
|
Corynecoris[127] |
|
| |||
|
Macropterocoris[127] |
|
| |||
|
Ophthalmocoris[127] |
|
| |||
|
Deraiocoris[127] |
|
| |||
|
Eogerridium[127] |
|
| |||
|
Pronabis[127] |
|
| |||
|
Entomecoris[127] |
|
| |||
|
Ensphingocoris[127] |
|
| |||
|
Engynabis[127] |
|
| |||
|
Xulsigia[144] |
|
|
Isolated Wings |
A Sternorrhynchan of the family Pincombeomorpha. It is curious for its peculiar venation on its wings. Has been proposed its own family, Xulsigiidae. |
|
|
Indutionomarus[145] |
|
|
Isolated Wings |
A Coleorrhynchan of the family Progonocimicidae. |
|
|
Xyelula[146] |
|
|
Isolated Wings |
A Pseudo-Wasp of the family Sepulcidae. |
|
|
Thilopterus[146] |
|
|
Isolated Wings |
A Wasp of the family Ephialtitidae. |
|
|
Symphytopterus[147] |
|
| |||
|
Liadobracona[148] |
|
| |||
|
Pseudoxyelocerus[149] |
|
|
Isolated Wings |
A Wood Wasp of the family Xyelotomidae. |
|
|
|
Isolated Wings |
A Lacewing of uncertain placement. |
||
|
|
Isolated Wings |
A Lacewing of the family Prohemerobiidae. |
||
|
Parhemerobius[127] |
|
| |||
|
Paractinophlebia[150] |
|
| |||
|
| ||||
|
Stenoteleuta[150] |
|
| |||
|
Actinoptilon[150] |
|
|
Isolated Wings |
A Silky Lacewing of the family Psychopsidae. |
|
|
Panfilovia[127] |
|
|
Isolated Wings |
A Lacewing of the family Panfiloviidae. A large genus with wings around 50 mm. |
|
|
Epipanfilovia[127] |
|
| |||
|
|
Isolated Wings |
A Giant Lacewing (Kalligrammatidae) of the subfamily Liassopsychopinae and Kallihemerobiinae. It is one of the oldest known representatives of the Giant pollinator lacewings. The genus Liassopsychops was previously referred to Psychopsidae. Another specimen related, Specimen Hi 21 is regarded here as Kalligrammatidae incertae sedis. This along the occurrence of two distantly-related genera of Kalligrammatidae in the lower Toarcian is unexpected.[151] The Toarcian Kalligrammatidae lived in warm and dry conditions, as proven by its recovery from the zone of Hondelange but absence on Grimmen and other northern locations. Both genera where among the largest insects found on all the formation.[151] |
||
|
Ophtalmogramma[151] |
|
| |||
|
Mesopsychopsis[150] |
|
|
Isolated Wings |
A lance Lacewing of the family Osmylopsychopidae. |
|
|
Tetanoptilon[127] |
|
|
Isolated Wings |
Lance Lacewings of the family Osmylidae. Tetanoptilon is the largest non-Kalligrammatidae lacewing of the Jurassic, with a forewings length up to 470 mm and a wingspan estimated at 11 cm. Like the Kalligrammatids, this genus is limited to the southern zones, asbsent in northern deposits.[151] |
 Extant member of Osmylidae, Posidonia genera where probably similar |
|
Mesosmylina[150] |
|
| |||
|
Protoaristenymphes[152] |
|
|
Isolated Wings |
A lance Lacewing of the family Mesochrysopidae. |
|
|
|
Isolated Wings |
A Hairy Cicada of the family Tettigarctidae. |
 Extant member of Tettigarctidae, Posidonia genera where probably similar | |
|
Liassotettigarcta[141] |
|
| |||
|
Adelocoris[127] |
|
|
Isolated Wings |
Pentatomomorphans of the family Pachymeridiidae. Are related with the family Lygaeoidea, being possible ancestral forms of this last one. |
 Extant member of Lygaeoidea, Posidonia genera where probably similar |
|
Stiphroschema[127] |
|
| |||
|
Engerrophorus[127] |
|
| |||
|
Euraspidium[127] |
|
| |||
|
| ||||
|
Mesomphalocoris[127] |
|
| |||
|
Trachycoris[127] |
|
| |||
|
Apicasia[127] |
|
|
|
Beetles of uncertain placement inside Coleoptera. Diverse beetle specimens that due to its preservation (incomplete) or its morphological traits are not assigned to any concrete family. The abundance of beetle Elytrons indicates the proximity of terrestrial habitats. |
|
|
Apiopyrenides[127] |
|
| |||
|
Auchenophorites[127] |
|
| |||
|
Brachytrachelites[127] |
|
| |||
|
Diatrypamene[127] |
|
| |||
|
Diatrypamene[127] |
|
| |||
|
Dicyphelus[127] |
|
| |||
|
Dicyphelus[127] |
|
| |||
|
Diplothece[127] |
|
| |||
|
Entomocantharus[127] |
|
| |||
|
Episcepes[127] |
|
| |||
|
Eurynotellus[127] |
|
| |||
|
Eurysphinctus[127] |
|
| |||
|
Eusarcantarus[127] |
|
| |||
|
Gastrodelus[127] |
|
| |||
|
Laimocenos[127] |
|
| |||
|
Leptomites[127] |
|
| |||
|
Loxocamarotus[127] |
|
| |||
|
Macrotrachelites[127] |
|
| |||
|
Melanocantharis[127] |
|
| |||
|
Mesotylites[127] |
|
| |||
|
Ooidellus[127] |
|
| |||
|
Ooperiglyptus[127] |
|
| |||
|
Opiselleipon[127] |
|
| |||
|
Oxycephalites[127] |
|
| |||
|
Parnosoma[127] |
|
| |||
|
Proheuristes[127] |
|
| |||
|
Sphaericites[127] |
|
| |||
|
Tetragonides[127] |
|
| |||
|
Trichelepturgetes[127] |
|
| |||
|
Trochmalus[127] |
|
| |||
|
Pholipheron[127] |
|
| |||
|
Grasselites[127] |
|
| |||
|
Omogongylus[127] |
|
| |||
|
Sideriosemion[127] |
|
| |||
|
Metanastes[127] |
|
| |||
|
Aptilotitus[127] |
|
| |||
|
Rhomaleus[127] |
|
| |||
|
Leptosolenophorus[127] |
|
| |||
|
Gastroratus[127] |
|
| |||
|
Brachylaimon[127] |
|
| |||
|
Pleuralocista[127] |
|
| |||
|
Mesoncus[127] |
|
| |||
|
Palaeotrachys[127] |
|
| |||
|
Hydroicetes[127] |
|
| |||
|
Scalopoides[127] |
|
| |||
|
Peridosoma[127] |
|
| |||
|
| ||||
|
Sphaerocantharis[127] |
|
| |||
|
Rhysopsalis[127] |
|
| |||
|
Diplocelides[127] |
|
| |||
|
Tripsalis[127] |
|
| |||
|
Trochiscites[127] |
|
| |||
|
Zetemenos[127] |
|
| |||
|
Amphoxyne[127] |
|
| |||
|
| ||||
|
Prosynactus[127] |
|
|
|
False Ground Beetles of the family Trachypachidae. |
 Extant member of Trachypachidae, Posidonia genera where probably similar |
|
Coreoeicos[127] |
|
| |||
|
Aposphinctus[127] |
|
|
|
A Water Scavenger Beetle of the family Hydrophilidae. |
 Extant member of Hydrophilidae, Posidonia genera where probably similar |
|
Amblycephalonius[127] |
|
|
|
Beetles of the family Coptoclavidae. |
|
|
Ooperioristus[127] |
|
| |||
|
Camaricopterus[127] |
|
|
|
A Beetle of the family Phoroschizidae. |
|
|
Megachorites[127] |
|
|
Elytrons |
A Giant Beetle. It is among the largest found on all the Jurassic. |
|
|
|
Isolated Wings |
A Phantom Crane fly of the family Eoptychopterinae. |
||
|
|
Isolated Wings |
An Amphiesmenopteran of the family Necrotauliidae. The ovipositor, like terminalia of female N. parvulus, indicate that these insects laid their eggs rather in soil than in water |
||
|
Micropterygidae[155] |
|
|
Isolated Wings |
Lepidopterans probably related with the family Micropterygidae. Compared with their record on Grimmen, on Lower Saxony Lepidopterans are rather scarce and bad preserved. |
_-_(imago)%252C_Zemst%252C_Belgium.jpg.webp) Extant member of Micropterygidae, Posidonia genera where probably similar |
|
Geisfeldiella[156] |
|
|
Isolated Wings |
Mayfly of the family Protereismatidae. |
|
|
Protobittacus[127] |
|
|
Isolated Wings |
Hanginflies of the family Bittacidae. |
 Extant member of Bittacidae, Posidonia genera where probably similar |
|
Parabittacus[127] |
|
| |||
|
Neorthophlebia[127] |
|
| |||
|
Pleobittacus[127] |
|
| |||
|
Haplobittacus[127] |
|
| |||
|
Mesobittacus[127] |
|
| |||
|
Orthophlebia[127] |
|
|
Isolated Wings |
Scorpionflies of the family Orthophlebiidae. |
|
|
Reprehensa[127] |
|
| |||
|
Parorthophlebia[127] |
|
| |||
|
Mesopanorpa[127] |
|
| |||
|
|
Isolated Wings |
Scorpionflies of the family Protorthophlebiidae. |
||
|
Pseudopolycentropus[127] |
|
|
Isolated Wings |
Scorpionflies of the family Pseudopolycentropodidae. |
|
|
Homoeoptychopteris[127] |
|
|
|
Flies Of Uncertain Placement. Some of the Specimens where listed as Nymphs but are now tougth to be wingless Dipterans |
|
|
Liassonympha[127] |
|
| |||
|
Rhopaloscolex[127] |
|
| |||
|
Empidocampe[127] |
|
| |||
|
Amianta[127] |
|
| |||
|
Culiciscolex[127] |
|
| |||
|
Bodephora[127] |
|
| |||
|
Apistogrypotes[127] |
|
| |||
|
Amphipromeca[127] |
|
| |||
|
Cyrtomides[127] |
|
| |||
|
Sphallonymphites[127] |
|
| |||
|
Propexis[127] |
|
| |||
|
Amblylexis[127] |
|
| |||
|
Ellipibodus[127] |
|
| |||
|
Archipleciomima[157] |
|
| |||
|
Protoplecia[127] |
|
|
Isolated Wings |
A Fly of the family Protopleciidae. |
|
|
Mesorhyphus[157] |
|
|
Isolated Wings |
A Wood Gnat of the family Anisopodidae. |
_female_(13066528293).jpg.webp) Extant member of Anisopodidae, Posidonia genera where probably similar |
|
|
Isolated Wings |
A Snakefly of the family Priscaenigmatidae. |
||
|
Metaraphidia[148] |
|
|
Isolated Wings |
A Snakefly of the family Metaraphidiidae. |
|
|
Heterorhyphus[127] |
|
|
Isolated Wings |
A Fly of the family Heterorhyphidae. |
|
|
Protorhyphus[127] |
|
|
Isolated Wings |
A Fly of the family Protorhyphidae. |
|
|
Metatrichopteridium[158] |
|
|
Isolated Wings |
A Fly of the family Hennigmatidae. It represents the oldest know genus of this primitive family. |
|
|
Praemacrochile[127] |
|
|
Isolated Wings |
A primitive Crane fly of the family Tanyderidae. Extant members of the family are nectar feeder while extinct members cannot be determined precisely.[159] |
 Extant member of Tanyderidae, Posidonia genera where probably similar |
|
|
Isolated Wings | |||
|
Architipula[127] |
|
|
Isolated Wings |
A Crane fly of the family Limoniidae. |
 Extant member of Limoniidae, Posidonia genera where probably similar |
|
Ozotipula[127] |
|
| |||
|
Haplotipula[127] |
|
| |||
|
Leptotipuloides[127] |
|
| |||
|
Mikrotipula[127] |
|
| |||
Echinodermata
Echinoderm debris is pretty abundant on the shale-free Unken and Salzburg members, including Crinoid skeleton elements, also that of the Ophiurida; the Echinoids take their place, where really blossomed at that time. That's why Pedicellaria are observed very often.[37]
Asterozoa
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Barbaraster[161] |
|
|
Isolated Arm Plates |
An Ophiuridan of the family Ophiomusina. |
|
|
Dermacantha[161] |
|
|
Isolated Arm Plates |
An Ophiuridan of incertae sedis family on the order Ophionereididae. |
|
|
Enakomusium[162] |
|
|
|
An Ophiuridan of the family Ophiolepididae. |
 Holotype specimens from the Sachrang Formation |
|
Dermocoma[161] |
|
|
Isolated Arm Plates |
An Ophiuridan of incertae sedis family on the order Ophiodermatina. |
|
|
Inexpectacantha[161] |
|
|
Isolated Arm Plates |
An Ophiuridan of the family Euryophiurida. |
|
|
Lapidaster[161] |
|
|
Isolated Arm Plates |
An Ophiuridan of the family Ophioscolecidae. |
|
|
Ophiarachna[163] |
|
|
Isolated Arm Plates |
An Ophiuridan of the family Ophiacanthida. Very Common, related to non anoxic water sedimentation. |
|
|
Ophiogojira[161] |
|
|
Isolated Arm Plates |
An Ophiuridan of the family Ophiomusaidae. |
|
|
Ophiocopa[161] |
|
|
Isolated Arm Plates |
An Ophiuridan of the family Ophiotomidae. |
|
|
?Ophiocten[163] |
|
|
Isolated Arm Plates |
An Ophiuridan of the family Ophiuridae. Its relationships haven't been confirmed and it is based on very fragmentary remains. |
|
|
Ophiohelus[161] |
|
|
Isolated Arm Plates |
An Ophiuridan of the family Ophiohelidae. |
|
|
|
Isolated Arm Plates |
An Ophiuridan of the family Ophiomusaidae. |
||
|
Ophiomisidium[161] |
|
|
Isolated Arm Plates |
An Ophiuridan of the family Astrophiuridae. |
|
|
|
Isolated Arm Plates |
An Ophiuridan of the family Ophiactidae. Very rare on the layers. |
_(20395173566).jpg.webp) Modern Specimen | |
|
Ophiotardis[161] |
|
|
Isolated Arm Plates |
An Ophiuridan of the family Ophiopyrgidae. |
|
|
|
Isolated Arm Plates |
An Ophiuridan of the family Ophiopyrgidae. |
||
|
|
|
An Ophiuridan of the family Ophioleucidae. The Mostly know Arterozoan from the formation, including a single articulated specimen. |
_90.3.9802.jpg.webp) Fossil Specimen | |
|
Thanataster[161] |
|
|
Isolated Arm Plates |
An Ophiuridan of the family Ophiomusina. |
|
Echinoidea
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Specimens |
A sea urchin of the family Cidaridae. Common on several layers. Cidaris is genus that still alive today. A bottom dweller, is commonly found associated with Belemnnite fossils, probably due to eating its carcasses. |
 Modern Specimen | |
|
|
Specimens |
A sea urchin of the family Pedinidae. It is the most common sea urchin in the formation, present on all the levels with specimens of various sizes. Vinculated to sea bottom sediments, before Gasteropods and Bivalves, Diademospsis was the third major colonizer of the bottom, in between anoxic changes. |
 Specimens from Holzmaden | |
|
Eodiadema[44] |
|
|
Specimens |
A sea urchin of the family Diadematidae |
 Specimens from Holzmaden |
|
Hemipedina[44] |
|
|
Specimens |
A sea urchin of the family Pedinidae |
|
|
Procidaris[27] |
|
|
Specimens |
A sea urchin of the family Miocidaridae |
|
|
Pseudodiadema[44] |
|
|
Specimens |
A sea urchin of the family Pseudodiadematidae |
|
Holothuroidea
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Specimens |
A sea cucumber of the family Achistridae inside Apodida. The "fishhooks" from this genus are only rarely found in the tenuicostatum zone |
 Specimen | |
|
Mortensenites[25] |
|
|
Specimens |
A sea cucumber of the family Calclamnidae inside Dendrochirotida. A Genus whose identification was originally complex. |
|
|
|
Specimens |
A sea cucumber of the family Stichopitidae. Occurs sporadically in non-bituminous sediments of the upper bifrons zone |
||
|
|
Specimens |
A sea cucumber of the family Chiridotidae. It is the only major genus of Sea Cucumbers reported locally on the Posidonienschiefer. It was identified originally as Chirodonta mesoliasicus and Chirodonta heptalmorpha. Represents a possible cosmopolitan holothurian that occupied all three types of deep-sea ecosystems as an organic-enrichment opportunist. |
||
Crinoidea
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Type genus of Crinoidean from the family Pentacrinitidae. Like Seirocrinus, Pentacrinites formed colonies on rafting wood, getting a different role than bigger crinoid and appearing on the first stages of the decomposition of the rafting wood. Was a smaller genus, with specimens of no more than 1 meter long, usually measuring 40–70 cm. |
 Close view of one specimen from the Posidonienschiefer | |
|
Praetetracrinus[167] |
|
|
Isolated Stems |
A Crinoidean of the family Plicatocrinidae. |
|
|
Procomaster[163] |
|
|
Exceptionally well preserved individual with the arms,pinnules and cirri largely Intact |
A Crinoidean of the family Isocrinida. This Benthic Crinoid clearly represents an exotic elementement of the typical Posidonia fauna, likely moved from the coastal settings |
|
|
|
|
The largest known Crinoidean, from the family Pentacrinitidae. Among the tallest animals of its period, Seirocrinus is also one of the most famous fossils from the Sachrang Formation. It consists of fossils of colonies along large wood trunks, with specimens up to 14 m long, with the largest specimen reaching 26 m long, what makes it among the tallest know Mesozoic organisms, one of the largest invertebrates know on the fossil record and one of the tallest know animals.[169] It was an open ocean organism that lived in rafting woods, probably filtering food and serving as a refuge for other animals, such as ammonites. The crinoids had a large colonization process, based on the status of the fossil wood found.[170] The large rafts were the home for a high variety of marine organisms, such as Balanoideans, Ammonites and other. It has been estimated that without the presence of modern raft wood predators (that appeared on the Bathonian) those rafts can last up to 5 years, being that the main reason the crinoids were able to reach such huge sizes. The large rafts were also probably essential to distribute animals along the Early Jurassic Seas.[13] |
_(Posidonia_Shale%252C_Lower_Jurassic%253B_Holzmaden_area%252C_Germany)_1_(35674916591).jpg.webp) Close view of one specimen from the Posidonienschiefer | |
Vertebrata
Chondrichthyes
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Head and postcranial remains |
A member of Myriacanthidae inside Chimaeriformes. An aberrant Chimaera with an extrange elongated nose and horns over the skull. |
| |
|
|
Teeth |
Type genus of the family Acrodontidae. |
||
|
|
Upper ("palatine") toothplate |
A member of Callorhynchidae inside Chimaeriformes. Similar to Callorhinchus, among the oldest known of its type. It the first Modern lineage Chimaeras from the Toarcian. |
.gif) Callorhinchus milii can be the closest relative of Bathytheristes | |
|
|
Teeth |
A shark of the family Hybodontidae. An aberrant hybodontid with crushing dentition. |
| |
|
Crassodus[174] |
|
|
Meckelian Cartilages, Jaws, teeth, Palatoquadrates, placoid scales and dearticualted parts of the labial, hyoid and branchial skeleton. |
A shark of the family Hybodontidae. The Type specimen belongs to a large hybodontid, with an estimated total length of up to 3 m.[174] It has a Meckelian Cartilage more robust than Hybodus hauffianus and an acrodontine dentition, as seen on Bdellodus.[174] Probably is related with "Hybodus" delabechei, very likely a junior synonym of Crassodus.[174] |
|
|
|
|
Type genus of the family Hybodontidae. It is the most abundant shark on the layers of the Sachrang Formation, with some of the best preserved specimens of the genus known. It was probably an open ocean hunter, with small horns over the eyes. With a size around 2 m, it was also one of the largest representatives of the Chondrichthyes on the formation.[177] |
 Hybodus Holzmaden specimen, among the best preserved of the genus, with Belemmnites inside. | |
|
|
|
A member of Myriacanthidae inside Chimaeriformes. An aberrant Chimaera with a second jaw-like structure on its head. |
||
|
|
Anterior part of body with basicranium, palatoquadrates, Meckel's cartilage, ceratohyals, epihyals, teeth, traces of the branchial arches and the anterior finspine |
Type member of the family Palaeospinacidae. |
||
|
Palidiplospinax[181] |
|
|
Articulated vertebral column, girdles, both fin spines and clasper organ |
A member of the family Palaeospinacidae. |
|
| Pseudonotidanus[182] |
|
|
Partial, articulated specimen |
A shark of the family Hexanchiformes. It was identified originally as a member of the genus Palaeospinax. |
|
|
Gen et sp. nov |
|
SMNS 52666, Incomplete Specimen |
A possible member of Batoidea. It was originally identified as a member of Galeiformes. This Genus was found to be sister taxa to the extant genus Raja, yet maybe represents an stem Rajiform. It is probably the oldest described pelagic Rajiform. It wears an enlogated rostrum. |
 SMNS 52666, the unnamed rajiform holotype | |
|
Recurvacanthus[184] |
|
|
Isolated Fin Spine |
A member of Myriacanthidae inside Chimaeriformes. |
|

Actinopterygii
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Various Complete and nearly complete Specimens |
Type genus of the family Caturidae inside Amiiformes |
| |
|
|
|
A deep-bodied neopterygian, the type genus of the family Dapediidae. Unpublished material indicates the presence of one or even two more still undescribed species of Dapedium in the Lower Toarcian.[187] A specimen of Lytoceras has a Dapedium sp. in the body chamber probably trapped when feeding on the carcass of the cephalopod, and may therefore indicate an opportunistic carrion feeder.[188] |
| |
|
|
|
A member of the family Pachycormidae. |
| |
| Germanostomus[190] |
|
|
|
A pachycormid. |
|
|
Haasichthys[191] |
|
|
|
A pachycormid. |
|
|
|
|
A member of the family Furidae inside Ionoscopiformes |
||
|
Holzmadenfuro[192] |
|
|
Complete Specimen |
First ganoin-scaled Ophiopsiformes (Halecomorphi) from the Posidonienschiefer. The type specimen measures 51 cm, and has elongated and serrated body scales before the dorsal fin and tiny ganoid scales after it.[192] |
|
|
|
|
A common member of the Lepisosteiformes. |
| |
|
|
|
A member of the family Leptolepididae. The most common member of its family, Leptolepis is commonly associated with crustaceans and small marine invertebrates, probably the main constituents of its diet. One on the most predated vertebrates on the formation, with abundance of larger fishes and reptiles with specimens associated. |
| |
|
|
MB. f.7612, nearly complete specimen. |
A member of the family Leptolepididae. Was identified as Paraleptolepis, but this name is currently occupied by a Japanese fish genus of Early Cretaceous age.[198] It differs from Leptolepis coryphaenoides in the presence of a few autapomorphies and also in the retention of several primitive features not present on the last one.[197] Small genus, of about 14 cm length.[197] |
 Longileptolepis | |
|
|
|
A possible representative of the family Saurichthyidae. Is based on rather fragmentary specimens. |
||
|
Ohmdenfuro[192] |
|
|
Nearly complete specimen with broken skull |
First ganoin-scaled Ophiopsiformes (Halecomorphi) from the Posidonienschiefer. Elongated morphology, with a length of ~39 cm, covered by smooth, massive ganoin scales.[192] |
|
| Ohmdenia[199] |
|
|
|
A large member of the family Pachycormidae, with a length of up to 2.5–3 m and an estimated weight over 200 kg.[199] |
|
|
|
|
Type member of the family Pachycormidae. Large representative of its family, with a size up to 1.5 m. One specimen preserved the alimentary canal, with the stomach filled by numerous hooklets that can be referred to the coleoid cephalopod Phragmoteuthis, implying a diet of cephalopods for this genus.[18] |
| |
|
|
|
A member of the family Pholidophoridae. Is among the most abundant fishes on the late liassic of Europe, present on the sub-Mediterranean boreal, with specimens of several sizes. |
| |
|
|
|
Type genus of the family Ptycholepididae inside Ptycholepiformes. It is one of the youngest representatives of its family. |
| |
|
|
|
A large member of the family Pachycormidae. |
| |
|
|
|
The youngest representative of the family Saurichthyidae, known for its large jaws, similar to modern Belonidae. |
| |
|
|
|
A member of the family Pachycormidae. Large representative of the family, reaching sizes up to 2.3 m. |
| |
| Strongylosteus[206] |
|
|
|
A large member of the Chondrosteidae and the largest non-reptilian marine vertebrate of the Sachrang Formation, with a size between 3 and 4.5 m, and an estimated weight over 800 kg to 1 tonne.[206] |
|
|
|
|
A deep-bodied neopterygian of the family Dapediidae. |
| |














Sarcopterygii
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
A large coelacanth of the family Mawsoniidae, related to the genera Axelrodichthys, Chinlea, Diplurus and the type, Mawsonia.[209] The largest specimen known from the Sachrang Formation is GPIT.OS.770 (Holotype), with a length over 1.6 m.[208] The specimen presents an ossified lung inside the abdominal cavity, and most of the body, being also one of the most complete coelacanths of the Jurassic found.[208][209] Trachymetopon precedes the presence of the family Mawsoniidae in Europe by about 120 Ma and the northernmost occurrence of a member of the group, implying an extensive geographical range during the Early Jurassic.[209] Due to the specimens being found on pelagic deposits suggest that probably was an open ocean swimmer.[208][209] |
 Trachymetopon GPIT.OS.770 | |
Ichthyosauria
Inderminate specimens are known.[27][193][194][195]
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
A large ichthyosaur of the family Leptonectidae with convergent evolution with modern swordfish. Like these fishes, Eurhinosaurus is believed to be a fast swimming predator, able to hunt fish schools on same way. Large specimens of up to 6 m are known. |
 Complete specimen from the Sachrang Formation | |
|
|
Complete Specimens |
Small sized ichthyosaur, probably a member of Parvipelvia, sister group to Stenopterygius + Ophthalmosauridae. A small- to mid-sized ichthyosaur, 2–3 m in length, with a relatively short and slender antorbital rostrum.[213] |
 Hauffiopteryx typicus fossil | |
|
|
|
A possible member of the family Leptonectidae. Mostly of the specimens of this genus have been referred to Leptonectes or Temnodontosaurus, although some remains on the Posidonienschiefer are too complex to being clearly referred. |
||
|
|
|
An Ichthyosaur of the family Stenopterygiidae. Magnipterygius may not have grown to a total length of much more than 120 cm. It is therefore potentially only the second post-Triassic ichthyosaur known with such a small body size |
||
|
|
|
Type genus of the family Stenopterygiidae. A common Toarcian ichthyosaur, present in multiple layers. The rather exquisite level of preservation has led to know even the coloration, that exposes a clear countershading, with an upper part being more obscure than the lower, similar to modern killer whales, the Heaviside's dolphin or the Dall's porpoise. There is also evidence of changes in color with ontogenic changes, going from dark juveniles to countershaded adults. The skin was flexible & scaleless, as in dolphins.[217] The study of several specimens has revelated that Stenopterygius quadriscissus underwent a size-related trophic niche shift through ontogeny, shifting from a piscivorous diet to a teuthophagous diet, known thanks to exquisitely preserved stomach contents.[218] |
.jpg.webp) Fossil | |
|
|
|
Type genus of the family Suevoleviathanidae. Includes specimens up to 4 m long. |
 Suevoleviathan integer fossil | |
|
|
|
Type genus of the family Temnodontosauridae. A large macroraptorial ichthyosaur, apex predator of its environment. It ranges between 9 and the 12 m, being one of the largest known ichthyosaurs, characterized by skulls and jaws over 1 m in length, with the largest being over 1.9 m long. It has been found with fragments of young icthyosaur in his stomach.[17] |
 Temnodontosaurus trigonodon specimen. | |
Plesiosauria
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Complete specimen. |
A basal member of Pliosauridae. A moderately sized (3.4 m) Pliosaurid, ecologically adapted to fish hunt, as has been observed due to comparing the long snouted skull with that of Peloneustes, Gharial Crocodiles or Dolphins.[224] |
| |
|
|
|
A junior synonym of M. brachypterygius.[226] |
| |
|
|
|
A Rhomaleosauridae Plesiosaur. Its detailed fossils have helped to study plesiosaur movement. |
| |
|
|
|
Type member of the Plesiosaur family Microcleididae. Small Plesiosaur, with a length of less than three metres. It is characterised by a really elongated neck, and was probably an ichthyophagous form that occurred rarely in the Posidonienschiefer fauna. |
| |
|
|
|
An indeterminate plesiosauroid plesiosaur. An impressively preserved immature specimen, different from Hydrorion brachypterygius and Seeleyosaurus guilelmiimperatoris, the most abundant plesiosaurs found locally, and its anatomical characters suggest it represents a new genus.[226] It fossilized buff-coloured material, identified as mainly composed of calcium phosphate and interpreted as phosphatised muscle tissues.[226] It also contains eumelanin and hence possibly corresponds to areas dark-coloured in life.[226] The stomach contains quartz grains that were most likely ingested during the animal's life, maybe used for food trituration. This gastroliths have importance for its exotic provenance compared with the surrounding lithology of the deposits (mostly shale), and as sandy turbidites have never been reported from the SW German Basin, the individual may have acquired the sand-sized grains many kilometres away from the burial site.[226] Strata containing fine sand ('Glaukonit und viel Feinsand') at Obereggenen im Breisgau (western side of the Black Forest between Freiburg and Basel), 200 km from Holzmaden suggest a nearshore deposit was allocated here, and that the black forest emerged at this time. Probably this young specimen reached that location in search for gastroliths.[226] |
||
|
|
Complete Specimen. |
A basal Plesiosaur that has been linked with Cryptoclididae. It is one of the smallest from the Posidonia, with a complete skeleton measuring less than 2.5 m. It is considered a possible junior synonym of Seeleyosaurus. |
| |
|
|
Isolated caudal & cervical vertebrae |
A Plesiosaur assigned to the genus Plesiosaurus, yet shows more affinities with Anningasaura |
||
|
|
|
A Plesiosaur of the family Microcleididae. It was named originally "Plesiosaurus guilelmiimperatoris". It was a moderate-sized plesiosauroid, measuring up to 3,5 m in length with a skull length of 170 mm. |
| |






Sphenodontia
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
An aquatic sphenodont of the family Pleurosauridae. Palaeopleurosaurus evidences that there was a slightly skeletal specialization for an aquatic lifestyle, achieved through the Jurassic gradually on pleurosaurs.[232] Recent studies suggest a shorter lifespan than modern Tuatara, based on irregular spacing of growth marks.[233] |
| |

Testudinata
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
A marine turtle of the superfamily Eurysternidae inside Thalassochelydia. Is the main formally identified fossil as Turtle from the Sachrang Formation, representing a rather basal genus. The pleurals resemble those of the genus Plesiochelys.[234] Being found in the zone of Franconia that on the Toarcian was at -80 km from the shore can suggest that early marine turtles lived in the epicontinental waters of the European shallow seas before reach richer ecosystem diversity on the Late Jurassic.[234] This would explain the serious lack of Turtle fossils on the formation, as mostly of the deposits are located far from the coast.[234] |
||
|
|
|
Possible unclassified testudine remains. Münster (1834) cited: "there were also rare things at the quarries of Altdorf, among other remains there were ones of a turtle on lias limestone"". The remains are not catalogued and some specimens are in Private Collections.[237] |
||
|
|
|
A marine turtle of the clade Thalassochelydia inside Angolachelonia. The only testudine identified from the northern realm of the Sachrang Formation.[238] |
||
Crocodylomorpha
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
A longirostrine thalattosuchian of the family Machimosauridae. Was considered synonymous with Steneosaurus until in 2020 this last was recovered as invalid. It reached large sizes, with specimens exceeding 5 m, being a generalist predator.[239] |
 Macrospondylus bollensis fossil with Skin Impressions | |
|
|
|
A mesorostrine thalattosuchian of the family Teleosauridae. A marine crocodylomorph with a diet probably based on fish. Was considered synonymous with Steneosaurus until recently.[242] Due to this unusual placement of the external nares, Mystriosaurus was more terrestrial, or spent a greater amount of time on land, than other teleosauroids. This would explain its greater presence in zones of the formation more proximal to the emerged landmasses. Its morphology suggest it was a mesorostrine generalist.[242] |
| |
|
|
|
A thalattosuchian with a complex assignation, probably the basalmost metriorhynchoid. Pelagosaurus typus was a small-bodied thalattosuchian (~1 m in length) considered to be an adept aquatic pursuit predator, with a long streamlined snout ideal for snapping at fast moving prey (one specimen was found with Leptolepis fishes inside) and large, anterolaterally placed orbits for increased visual acuity.[243] |
| |
|
|
|
A Longirostrine Thalattosuchian, the most basal know. Was considered synonymous with Steneosaurus. Longirostrine specialist, probably active fish hunter. |
||
|
|
|
A longirostrine thalattosuchian of the family Teleosauridae. Platysuchus was slightly more robust than its contemporaneous relatives, being probably adapted to hunt more voluminous fish. A heavily armoured, semi-terrestrial longirostrine generalist form, indicated by the extensive and tightly packed rows of dorsal osteoderms.[239] |
| |
-05.JPG.webp)


Pterosauria
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
A Novialoidean Pterosaur, type genus of the family Campylognathoidea. Mark Witton suggests the construction of Campylognathoides' extremely robust forelimbs, with proportionally long wing fingers, could be a specialization for a fast aerial lifestyle comparable to those of Falcons and mastiff bats, being more probably an insect & vertebrate hunter and living on nearshore environments. |
 Nearly complete Campylognathoides | |
|
|
A Rhamphorhynchinae Pterosaur. It is one of the best known Early Jurassic Pterosaurs.[246] Unlike Campylognthoides, Dorygnathus was an oceanic hunter, with teeth disposed to catch marine prey, such as Belemnittes and several species of fishes. Dorygnathus mistelgauensis is considered a junior synonym until more data can be recovered from the specimen, held on a private collection.[246] |
 Nearly complete Dorygnathus | ||
|
|
Pelvis and several vertebrae. |
A Novialoidean Pterosaur, probably a member of the family Campylognathoidea. Has been assigned to the genus Campylognathoides, although it is clearly different than any other pterosaur from the Sachrang Formation.[90] The name "Schandelopterus" is invalid and lacks any study, assigned without species to refer to the specimen on private German Fossil Groups. The pelvis indicates a laterally, slightly upwardly directed orientation of the acetabula which does not support a bird-like bipedal locomotion of this pterosaur as has been suggested on the past.[90] |
||
|
|
Skull |
A Rhamphorhynchinae Pterosaur. Has been assigned to the genus Dorygnathus. It has a really complete skull that can help to explain the status of the genus Parapsicephalus.[251] |
||
Dinosauria
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Tibia and astragalus |
A Gravisaurian Sauropod. One of the few formally described from the Toarcian. Was confused as a Plesiosaur bone.[252] |
| |
|
|
3 cm long, tooth-studded fragment of a lower jaw |
An indeterminate possible Sauropodomorph dinosaur, possibly a member of Sauropodiformes inside Anchisauria (Resemble Yunnanosaurus teeth).[253] The Fossil was reported from the Lias Epsilon level, that on Oedhof is occupied by the Sachrang Formation.[253] It was found with abundant Plant debris and Belemnite remains.[253] Hasn't been revised since 1956 |
 The Oedhof jaw can belong to a creature similar to Yunnanosaurus | |

Plantae

The macroflora of the Posidonia slate can be described as extremely poor in species.[254] Apart from the remains of Horsetails, it is without exception the remains of coarse branches and fronds from gymnosperms, in which one has a certain can assume transport resistance. Remains of Ferns are completely missing, except for tall arboreal ferns (Peltaspermales).[255] Mostly of the flora was reported from the area of Braunschweig.[254] The major explanation for the flora could be that the plants in question are mono-or oligotypic stands on the edge of the waters that flow into the Posidonienschiefer sea, probably tear away in the course of flood events, easily fragmented during transport and wave waves, possibly especially in the occasional storm events postulated.[256] In terms of taphonomy, this would result in a comparison with today's reed Phragmites, which can form extensive stocks on the edge of shallower and slowly flowing waters ("Reed belts").[254] The Wood remnants clearly indicate one higher diversity of Coniferous flora in the delivery area than the remains of leafy branches.[254] This fact is likely to be proportionate, similar to that frequent occurrence of charcoalized or gagged trunks, mostly of them are believed to be "driftwoods" that only take a long time drifting also suggests a frequent settlement with mussels and full-grown Sea Lilies.[254][256] The deposition settings are at large distance from the nearest coastline (for southern Germany about 100 kilometers), making only plants strong to transportation able to resist enough to get deposited.[257][140] At Irlbach and Kheleim, NE of Regensburg, where the Posidonienschiefer has its near mainland deposit with abundant sand, a rich deposit filled with plant remains of different kind (Seds, Reproductive organs, Leafs, Stems, Cuticles and wood) with traces of coal was recovered, however, it was never studied in depth.[79] Of all the plant material expected only a few Bennetites leafs and two conifer branches with leaves where cited and none studied.[79] At the Austrian realm The sachrang Member was developed in the basinal area, while the Unken Member, sandwiched between red, often condensed limestones, represents the marginal facies.[37] Due to be more marginal and connected with the southern Vindelician land, the most diverse palynological assemblages of the formation are found, transported from zonas with moldanuvian granites as proven by the feldspar accumulations.[37]
Phytoclasts
Phytoclasts have been recovered from several sections on the formation, but only studied in depth from the Dotternhausen and specially Dormettingen.[43] Here two kinds of Phytoclasts where recovered, opaque phytoclasts (charcoal, indicator of wildfire activity on nearby landmasses, indicator of seasonal alterations of the water column) and translucent phytoclasts (indicator of proximal landmasses with high availability of wood and other plant material, as well transport conditions).[43] On the lowermost part of the section opaque phytoclasts are low (15% of the total organic matter) while translucent are incredibly abundant (40%), lowering its abundance to a 20-10% on the next section.[43] The Exaratum Subzone is the only one with an inverse trend and more abundance of opaque phytoclasts. On the Bifrons level, both types reach between a 15% and a 30%, showing a rapid increase, to decrease on the end of the section to values of less than 10%.[43] Opaque Phytoclasts, for a supposed marine deposit are relatively abundant on some sections, whose decreasing on others suggest (along with increasing levels of Kaolinite) an increased delivery of land plant material by rivers, from areas with wetter climate and less frequent fires, while its rise suggest the opposite, nearby continental setting with dry climate and continuous wildfire activity.[43]
Palynology
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Pollen |
Affinities with the families Peltaspermaceae, Corystospermaceae or Umkomasiaceae inside Peltaspermales. Pollen of Uncertain provenance, that can be derived from any of the members of the Peltaspermales. The lack o distinctive characters and bad conservation are among the main factors to make this Palynological residues difficult to classify. Arboreal to arbustive seed ferns. |
||
|
Baculatisporites[259] |
|
|
Spores |
Affinities with the family Osmundaceae in the Polypodiopsida. Near fluvial current ferns, related to the modern Osmunda regalis. |
|
|
|
Pollen |
| ||
|
|
Pollen |
Affinities with both Sciadopityaceae and Miroviaceae inside Pinopsida. This Pollen resemblance with extant Sciadopitys suggest that Miroviaceae can be an extinct lineage of sciadopityaceaous-like plants.[261] |
 Extant Sciadopitys. Cerebropollenites likely come from a related plant | |
|
|
Pollen |
Affinities with Cycadaceae and probably Cycadales. Alternatively can be Pollen from Bennettitales. It is the most abundant non conifer Pollen recovered on the formation, recovered on all the major sampled areas. Probably derived from arbustive cycads, this genus is related with dry settings, even from desertic regions. |
 Closer Look of Macrozamia cones, a common Cycad. Chasmatosporites maybe come from a related plant | |
|
Circulina[37] |
|
|
Pollen |
Affinities with the Cheirolepidiaceae inside Pinales. Pollen from Arboreal to Arbustive Plants. It is rare on the Samples measured. |
|
|
|
Pollen |
Affinities with Cheirolepidiaceae inside Coniferae. Pollen of medium to large arboreal plants, specially coniferales. |
||
|
|
Pollen |
Affinities with Cheirolepidiaceae inside Coniferae. Abundant on the Lower Jurassic of North and Southern Europe, represents pollen of medium to large arboreal plants, specially coniferales. The abundance of pollen of Classopollis and other thermophile plants was observed in this region in the lower Toarcian from the end of the antiquum (= tenuicostatum) zone to the middle of commune zone.[263] Classopollis is correlated with evaporites and are therefore associated with desert basins, but the shrubs may have also lived in xeric upland areas with seasonal fires. Evidence of fires is absent on the marine Posidonienschiefer, but has been recovered on the coeval nearshore calcareous sandstones.[263] It increases with the appearance of charcoal phytoclasts, as derived from dry settings with increased wilfires.[43] |
||
|
|
Pollen |
Affinities with Gnetopsida and probably Gnetophyta. Has Been considered Pollen of Chloranthaceae. However, it is to old for belonging to advanced Angiosperms. It probably comes from cones related to the Genera Piroconites kuesperti from the Lowermost Jurassic of Germany, resembling pollen of extant Ephedra and Welwitschia. |
 Closer Look of Ephedra cones, a common Gnetal. Clavatipollenites maybe come from a related plant | |
|
|
Spores |
Affinities with Gleicheniaceae inside Gleicheniales. Suggest relative increase of humidity on the rivers flowing towards the Austrian realm. Most abundant Fern spore in this region. |
.jpg.webp) Dicranopteris, Concavisporites come probably from similar genera | |
|
|
Pollen |
Affinities with Cheirolepidiaceae and Araucariaceae inside Pinaceae. Non concreted affinities |
||
|
Cyathidites[43] |
|
|
Spores |
Affinities with the family Cyatheaceae inside Cyatheales. Arboreal Fern Spores |
 Example of extant Cyathea, Cyathidites come probably from similar genera |
|
Cycadopites[37] |
|
|
Pollen |
Affinities with the Cycadopsida inside Cycadales. Pollen related with modern Cycas, arbustive to lower floor plants, relatively abundant, present on various of the measured samples. The mos common found on the Austrian realm, indicator of dry settings. |
 Encephalartos, Cycadopites come probably from similar genera |
|
|
Spores |
Affinities with Dicksoniaceae inside Pteridopsida. Tree Fern Spores |
 Modern Dicksonia, Deltoidosporites come probably from similar genera | |
|
Densosporites[259] |
|
|
Spores |
Affinities with the Selaginellaceae in the Lycopsida. Herbaceous lycophyte flora, similar to ferns, found in humid settings |
 Extant Selaginella, typical example of Selaginellaceae |
|
|
Pollen |
Affinities with Podocarpaceae inside Pinopsida. Pollen From arbustive to arboreal plants |
.jpg.webp) Extant Afrocarpus Cone, example of the Podocarpaceae. Disaccites is similar to the pollen found on this genus | |
|
Exesipollenites[258] |
|
|
Pollen |
Affinities with the family Cupressaceae inside Pinopsida. Pollen that resembles extant genera such as the Genus Actinostrobus and Austrocedrus, probably derived from Dry environments. |
 Extant Austrocedrus. Exesipollenites maybe come from a related plant |
|
|
Spores |
Affinities with Selaginellaceae and probably Lycopsida. A rare element on the palynological records of the German Basin, although more abundant than any other Spore recovered locally. |
||
|
|
Pollen |
Affinities with the Pinidae inside Coniferae. Abundant on the Lower Jurassic of NW Europe. Its identification on the Posidonienschiefer is rather complex due to the bad preservation of the Pollen Grains. |
 Extant Pinus cembra Cone, example of the Pinidae. Inaperturopollenites is similar to the pollen found on this genus | |
|
Ischyosporites[37] |
|
|
Spores |
Affinities with Pteridopsida. Spores from several types of ferns, relatively rare, present only on 2 samples. |
|
|
Leptolepidites[259] |
|
|
Spores |
Affinities with the family Dennstaedtiaceae in the Polypodiales. Forest fern spores. |
.jpg.webp) Extant Dennstaedtia specimens; Leptolepidites probably comes from similar genera |
|
Lycopodiacidites[37] |
|
|
Spores |
Affinities with the Ophioglossaceae inside Filicopsida. Spores related with modern floor Ferns, that appear on abundant water locations. The Unken Member is considered a more basinal deposit, where Wood and Sporomorph remains are more common. |
.jpg.webp) Ophioglossum, Lycopodiacidites come probably from similar genera |
|
|
Spores |
Affinities with the family Osmundaceae inside Polypodiopsida. Near Fluvial currents ferns, reted to the modern Osmunda regalis |
_-_Cape_St._Mary's_Ecological_Reserve%252C_Newfoundland_2019-08-10.jpg.webp) Example of extant Osmunda specimens, Osmundacidites come probably from similar genera or maybe a species from the genus | |
|
Podocarpidites[259] |
|
|
Pollen |
Affinities with the Podocarpaceae inside Pinopsida. Conifer pollen from medium to large arboreal plants |
.jpg.webp) Extant Podocarpus. Podocarpidites maybe come from a related plant |
|
Polycingulatisporites[259] |
|
|
Spores |
Affinities with the family Notothyladaceae inside Anthocerotopsida. Hornwort spores. |
).png.webp) Example of extant Notothylas specimens |
|
Quadraeculina[259] |
|
|
Pollen |
Affinities with Podocarpaceae and Pinaceae inside Coniferophyta. |
|
|
Retitriletes[259] |
|
|
Spores |
Affinities with Lycopodiaceae inside Lycopsida. |
|
|
|
Pollen |
Affinities with Cheirolepidiaceae inside Pinaceae. Abundant on the Lower Jurassic of NW Europe. Spheripollenites co-occurs on the coeval Sorthat Formation with cuticles of Dactyletrophyllum ramonensis, and after a test of relationships it was found a highly significant correlation that may suggest that the species S. psilatus was produced by the conifer genus Dactyletrophyllum.[265] |
||
|
Stereisporites[259] |
|
|
Spores |
Affinities with Sphagnaceae inside Sphagnopsida. |
|
|
Striatella[259] |
|
|
Spores |
Affinities with Polypodiaceae inside Filicopsida. |
|
|
Todisporites[259] |
|
|
Spores |
Affinities with Osmundaceae inside Filicopsida. |
|

Equisetaceae
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Stems |
Affinities with Equisetaceae inside Equisetopsida. Number of mostly very fragmented and not particularly well preserved, but clear horsetail remains described. So far recognizable, leaf sheaths where developed in most cases, but the state of preservation does not allow a more precise determination.[266][267] |
||
|
|
Stems and incomplete axes |
Affinities with Equisetaceae inside Equisetopsida. Neocalamites is the most common more distributed of all the Posidonia Shale, being even found on Luxembourg Posidonia Strata.[268] Mostly of the Stems reported come from Aeolian-Dunar related deposits, or from nearshore-basinal deposition. Probably was related to the seashore.[268] Some stems are big, resembling the rates of growth seen on modern Bamboo specimens, suggesting +6–7 m tall Equisetopsids.[268] |
 Neocalamites merianii specimen | |
Pteridospermatophyta
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Affinities with Umkomasiaceae inside Corystospermaceae. Is based on bipinnate leaves, rachis longitudinally striated, with a long petiole and secondary rachises. It belongs to large tree ferns. Sachrang Formation Specimen is characterized for its large size and probably where attached to trunks similar in built to the Cretaceous genus Tempskya.[269] |
 The only Pachypteris specimen know from the Sachrang Formation | |
Bennettitales
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
A member of Williamsoniaceae inside Bennettitales. Identified originally as Zamites oblongifolius |
||
|
|
|
Affinities with Cycadeoidaceae inside Bennettitales. It is the most abundant medium-sized plant on the environment. Found specially on seashore depositional settings, but also on deltaic and lagoonar environments. It was a low arbustive-arboreal Bennetite related to arid environments, with a leave similar of that of the modern genus Encephalartos, specially Encephalartos munchii, but also Dioon mejiae. Otozamites has been considered synonym with Otopteris, but since the 1990s everybody used the name Otozamites, and Otopteris was forgotten.[272] |
 Otozamites gracilis specimen from the Sachrang Formation | |
|
|
|
Affinities with Cycadeoidaceae inside Bennettitales. This Bennettitalean is related to Shrub built. Some specimens were assigned to Dioonites acutifolium (Junior synonym). Consists on leaves with pinnate, leaflets perpendicular or oblique to the rhachis, on the top of the leaf axis. It comes from fragments of fairly large fronds. In its external form it closely follows the modern genus Dioon, only the leaflets are of our kind wider and shorter, they are further apart, but the tendency to rhachis is the same. |
 Pterophyllum fossil | |
|
|
|
Affinities with Williamsoniaceae inside Bennettitales. Arboreal Cycadaceans, some with the presence of flower-like structures. It resembles the leaf of the modern Microcycas calocoma, and probably had a similar arboreal built, being the leave of tall Bennetite trees such as Bucklandia, found on the middle jurassic of England. |
 Ptilophyllum sp. specimen from the Sachrang Formation | |
|
|
|
A member of Williamsoniaceae inside Bennettitales. It has been interpreted as a cycad in the family Cycadaceae or a Bennettitalean plant, and also a late surviving member of Noeggerathiales. Leaflets somewhat removed, oval-oblong, a little narrower near the base, rounded at the tip, nerves partially diverging from the base towards the edge. It was assigned to Pterophyllum oblongifolium and on the genus Glossozamites. This genus was the leaf of arboreal Bennetites, similar in appearance to the modern Encephalartos woodii. |
 Zamites mandelslohi specimen | |
Ginkgoales
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Affinities with Ginkgoaceae inside Ginkgoales. Various leaves are known from Ohmden, coming from marine deposits and where identified as Ginko digitata, identified originally as the only Ginko specimen reported on the Sachrang Formation. In the southern and northern Germany, there are regular remains of coal, which are initially reminiscent of small Ginkgo leaves. The leaves are hard to identify, more or less regularly concentric structures, as they sometimes appear like the coarse fruiting bodies of wood-dwelling fungi, such as the genus Trametes. |
 Ginkgoites digitata specimens | |
Pinophyta
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Affinities with Araucariaceae or Cheirolepidiaceae inside Pinales. Specimens whose spiral foliage of the branch, in which the individual leaves open about 2/3 of their length (without the tip) are fused with the branch they hold in their free part fits tightly, suggests belonging to the genus Brachyphyllum. |
||
|
Conites[195] |
|
|
|
Affinities with Araucariaceae or Cheirolepidiaceae inside Pinales. Represent various kinds of cones from diverse conifer origin. |
|
|
|
Ovuliferous dwarf-shoots |
Affinities with Cheirolepidiaceae. Is the type genus of this family, and is related with Arid Settings. Coming from The land-plant-taphocoenose from posidonia slate 3, is ready for processing. |
||
|
|
|
Affinities with Araucariaceae or Cheirolepidiaceae inside Pinales. Pagiophyllum araucarinum predominates among the two types of leafy coniferous branches that have become known from the Posidonia. However, there is no indication whether this fact reflects their respective share in the vegetation of the delivery area. Cheirolepidiaceae Pollen is the most abundant and diverse found on the formation, what is correlated with the abundance of this genus. Other factor that puts local Pagiophyllum on Cheirolepidiaceae is the dominance of an arid climate, the preferred for this type of conifers. |
 Pagiophyllum kurri specimen from Banz | |
|
|
|
A possible ancestral member of the Callitroideae inside Cupressaceae, or a member of Cheirolepidiaceae. Named also "Cupressites" liasinus, represents probably Arbustive to arboreal-derived axis. |
 Widdringtonites liasinus specimen from the Sachrang Formation | |
Fossil wood
Fossil Wood increases on the marginal Unken Member, with great amounts of logs and fragments of more than 1 m. Surface studies suggest relationships with the wood genera identified on the coeval Úrkút Manganese Ore Formation.[274]
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Fossil wood |
Affinities with Araucariaceae inside Pinales. Has araucarioid structures, but lacks proper characters of extant genera such as Araucaria & Agathis The largest known rafting wood on the fossil record is assigned to this genus, with a length of 18 m. The rafts were populated with Crinoid colonies, and a wide variety of organisms.[14] |
 Araucarioxylon (=Agathoxylon) reconstruction | |
|
Araucariopytis[44] |
|
|
Fossil wood |
Affinities with Abietinae inside Pinales. This genus was found to be related with Protocedroxylon found on the coeval Whitby Mudstone, crucial to the interpretation of the fossil record of Early Abietinae. |
 Abies specimen, whose role was probably covered by the makers of the Araucariopytis woods. |
|
|
Fossil wood |
Affinities with Podocarpaceae inside Pinales. Acmopyle pancheri is the only living conifer that has real resemblance with the wood assigned to Circoporoxylon. Xenoxylon phyllocladoides and C. grandiporosum are commonly found associated, for example in Braunschweig area. |
||
|
|
Fossil wood |
Affinities with Cupressaceae inside Pinales. Wood with a morphology similar to the modern genus Metasequoia. Probably related to Widdringtonites liasinus. |
 Metasequoia specimen, whose role was probably covered by the makers of the Cupressinoxylon woods. | |
|
|
Fossil Wood |
Affinities with Podocarpaceae inside Pinales. resembling modern genera such as Dacrycarpus, with other specimens resembling Juniperus. Includes wood more related to nearshore arbustive Conifers (columnar or low-spreading Shrubs with long, trailing branches), being the most abundant, but also medium to large arboreal conifers from nearshore forests. |
 Podocarpoxylon specimen | |
|
|
Fossil wood |
Affinities with Cheirolepidiaceae inside Pinales. Includes large sized trunks up to 1.7 m tall and 115 cm wide. Wood of other species of this genus, such as Protocupressinoxylon purbeckensis grown on flood-influenced settings of vegetated soil with brackish-water, like modern Swamp Cypress. |
 Protocupressinoxylon catenatum specimen | |
|
|
Fossil wood |
Affinities with Podocarpaceae and Cupressaceae inside Pinales. The assignation of this genus is rather complex. |
 Bivalves added to a Protophyllocladoxylon driftwood | |
|
|
Fossil wood |
Wood of the Protopinaceae, a possible "morpho-group" of the family Cheirolepidiaceae. Belongs to an invalid group of mostly Paleozoic Woods, and all the woods assigned to the family should be treated as Incertae Sedis.[279] |
 Woodworthia, example of Protopinaceae | |
|
|
Fossil wood |
Affinities with Podocarpaceae inside Pinales. Phyllocladus is the extant genus of conifer whose wood is more similar. |
 Phyllocladus specimen, whose role was probably covered by the makers of the Phyllocladoxylon woods. | |
|
|
Fossil wood |
Affinities with the Cupressaceae inside Pinales. Probably related to the coastal settings. It maybe covered a similar role that modern Swamp Cypress, Taxodium distichum. |
.jpg.webp) Taxodium distichum specimen, whose role was probably covered by the makers of the Taxodioxylon woods. | |
|
|
Fossil wood |
Affinities with Coniferales, concretely is closer to the Podocarpaceae, Cupressaceae and in a lesser extend to the Cheirolepidiaceae. Finally can be a member of the extinct family Miroviaceae. It is the more abundant genus of wood present on the Bohemian Realm of the Sachrang Formation. |
||
References
- Hess, H. (1999). "Lower Jurassic Posidonia Shale of Southern Germany" (PDF). Fossil Crinoids. 3 (1): 183–196. doi:10.1017/CBO9780511626159.025. ISBN 9780521450249. Retrieved 3 March 2022.
- Martill, D. M. (1993). "Soupy substrates: a medium for the exceptional preservation of ichthyosaurs of the Posidonia Shale (Lower Jurassic) of Germany" (PDF). Kaupia. 2 (1): 77–97. Retrieved 3 March 2022.
- Schmid–Röhl, A.; Röhl, H. J. (2003). "Overgrowth on ammonite conchs: environmental implications for the Lower Toarcian Posidonia Shale". Palaeontology. 46 (2): 339–352. Bibcode:2003Palgy..46..339S. doi:10.1111/1475-4983.00302. S2CID 128413601. Retrieved 3 March 2022.
- Arp, G.; Gropengießer, S. (2016). "The Monotis–Dactylioceras Bed in the Posidonienschiefer Formation (Toarcian, southern Germany): condensed section, tempestite, or tsunami-generated deposit?". PalZ. 90 (2): 271–286. doi:10.1007/s12542-015-0271-7. S2CID 128091360. Retrieved 2 March 2022.
- Birzer, F.; Joos, O. (1936). "Die Monotisbank in den Posidonienschiefern, besonders Frankens: Zur Geologie der Ehrenbürg (Walberla) bei Forchheim. Bayer" (PDF). Oberbergamt. 35 (2): 1–46. Retrieved 2 April 2022.
- Fraaye, R.; Jäger, M. (1995). "Decapods in ammonite shells: examples of inquilinism from the Jurassic of England and Germany". Palaeontology. 38 (1): 63–76. Retrieved 2 March 2022.
- Klompmaker, A.A.; Fraaije, R.H.B. (2012). "Animal Behavior frozen in time: gregarious behavior of early jurassic lobsters within an ammonoid body chamber". PLOS ONE. 7 (3): e31893. Bibcode:2012PLoSO...731893K. doi:10.1371/journal.pone.0031893. PMC 3296704. PMID 22412846.
- Maxwell, Erin E.; Cooper, Samuel L. A.; Mujal, Eudald; Miedema, Feiko; Serafini, Giovanni; Schweigert, Günter (2022). "Evaluating the Existence of Vertebrate Deadfall Communities from the Early Jurassic Posidonienschiefer Formation". Geosciences. 12 (4): 158–176. Bibcode:2022Geosc..12..158M. doi:10.3390/geosciences12040158.
- Jenny, D.; Fuchs, D.; Arkhipkin, A.I. (2019). "Predatory behaviour and taphonomy of a Jurassic belemnoid coleoid (Diplobelida, Cephalopoda)". Sci Rep. 9 (7944): 65–77. Bibcode:2019NatSR...9.7944J. doi:10.1038/s41598-019-44260-w. PMC 6538661. PMID 31138838.
- Glass, K.; Ito, S.; Wilby, P. R.; Sota, T.; Nakamura, A.; Bowers, C. R; Wakamatsu, K. (2013). "Impact of diagenesis and maturation on the survival of eumelanin in the fossil record". Organic Geochemistry. 64 (2): 29–37. Bibcode:2013OrGeo..64...29G. doi:10.1016/j.orggeochem.2013.09.002. Retrieved 2 March 2022.
- Klug, C.; Schweigert, G.; Fuchs, D. (2021). "Distraction sinking and fossilized coleoid predatory behaviour from the German Early Jurassic". Swiss J Palaeontol. 140 (7): 17–31. doi:10.1186/s13358-021-00218-y. PMC 7965854. PMID 33815267.
- Klug, C.; Schweigert, G.; Hoffmann, R. (2021). "Fossilized leftover falls as sources of palaeoecological data: a "pabulite" comprising a crustacean, a belemnite and a vertebrate from the Early Jurassic Posidonia Shale". Swiss J Palaeontol. 140 (10): 18–31. doi:10.1186/s13358-021-00225-z. PMC 8549986. PMID 34721282. S2CID 233450993.
- Hunter, A. W.; Mitchell, E. G.; Casenove, D.; Mayers, C. (2019). "Reconstructing the ecology of a Jurassic pseudoplanktonic megaraft colony". Royal Society Open Science. 7 (7): 12–31. doi:10.1098/rsos.200142. PMC 7428219. PMID 32874621.
- Matzke, A. T.; Maisch, M. W. (2019). "Palaeoecology and taphonomy of a Seirocrinus (Echinodermata: Crinoidea) colony from the Early Jurassic Posidonienschiefer Formation (Early Toarcian) of Dotternhausen (SW Germany)". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 291 (1): 89–107. doi:10.1127/njgpa/2019/0791. S2CID 134199163. Retrieved 3 March 2022.
- Gale, A.; Schweigert, G. (2016). "A new phosphatic-shelled cirripede (Crustacea, Thoracica) from the Lower Jurassic (Toarcian) of Germany–the oldest epiplanktonic barnacle". Palaeontology. 59 (1): 59–70. Bibcode:2016Palgy..59...59G. doi:10.1111/pala.12207. S2CID 128383968. Retrieved 2 March 2022.
- Schmidt, M. (1921). "Hybodus hauffianus und die Belemnitenschlachtfelder". Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg. 77 (1): 103–107.
- Thies, D.; Hauff, R. B. (2013). "A Speiballen from the lower jurassic posidonia shale of South Germany" (PDF). Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 267 (1): 117–124. doi:10.1127/0077-7749/2012/0301. Retrieved 10 February 2022.
- Přikryl, T.; Košták, M.; Mazuch, M.; Mikuláš, R. (2012). "Evidence for fish predation on a coleoid cephalopod from the Lower Jurassic Posidonia Shale of Germany". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 263 (1): 25–33. doi:10.1127/0077-7749/2012/0206. Retrieved 19 February 2022.
- Cooper, S. L.; Maxwell, E. E. (2023). "Death by ammonite: fatal ingestion of an ammonoid shell by an Early Jurassic bony fish". Geological Magazine. 160 (7): 1254–1261. Retrieved 2 October 2023.
- van Loon, A. J. (2013). "Ichthyosaur embryos outside the mother body: not due to carcass explosion but to carcass implosion". Palaeobiodiversity and Palaeoenvironments. 93 (1): 103–109. doi:10.1007/s12549-012-0112-6. S2CID 199406163. Retrieved 2 April 2022.
- Reisdorf, A. G.; Anderson, G. S.; Bell, L. S.; Klug, C.; Schmid-Röhl, A.; Röhl, H. J.; Wyler, D. (2014). "Reply to "Ichthyosaur embryos outside the mother body: not due to carcass explosion but to carcass implosion" by van Loon (2013)". Palaeobiodiversity and Palaeoenvironments. 94 (3): 487–494. doi:10.1007/s12549-014-0162-z. S2CID 129387302. Retrieved 2 April 2022.
- Dick, D.G. (2015). "An Ichthyosaur Carcass-Fall Community from the Posidonia Shale (Toarcian) of Germany". PALAIOS. 30 (2): 353–361. Bibcode:2015Palai..30..353D. doi:10.2110/palo.2014.095. S2CID 129077450. Retrieved 2 April 2022.
- Eriksson, M. E.; De La Garza, R.; Horn, E.; Lindgren, J. (2022). "A review of ichthyosaur (Reptilia, Ichthyopterygia) soft tissues with implications for life reconstructions". Earth-Science Reviews. 226 (1): 103–124. Bibcode:2022ESRv..22603965E. doi:10.1016/j.earscirev.2022.103965. S2CID 246846785. Retrieved 2 April 2022.
- Böhm, F.; Brachert, T. C. (1993). "Deep-water stromatolites and Frutexites Maslov from the early and Middle Jurassic of S-Germany and Austria". Facies. 28 (1): 145–168. doi:10.1007/bf02539734. S2CID 129365360. Retrieved 2 March 2022.
- Riegraf, W (1985). "Mikrofauna, Biostratigraphie, und Fazies im Unteren Toarcium Südwestdeutschlands und Vergleiche mit benachbarten Gebieten". Tübinger Mikropaläontologische Mitteilungen. 3 (1): 1–232.
- Ebli, O. (1989). "Foraminiferen und Coccolithen aus den Lias-Epsilon-Schiefern der Unkener Mulde (Tirolikum, Nördliche Kalkalpen)" (PDF). Mitt. Bayer. Staatsslg. Paläont. Hist. Geol. 29 (1): 61–83. Retrieved 3 March 2022.
- Riegraf, W. (1985). "Biostratigraphie, Fauna und Mikropaläontologie des Untertoarcium-Profiles von Unterstürmig (Oberfranken, Süddeutschland)". Geologische Blätter für Nordost-Bayern. 34 (1): 241–272. Retrieved 20 February 2022.
- Feist-Burkhardt, S.; Wille, W. (1992). "Jurassic palynology in southwest Germany–state of the art". Cahiers de Micropaléontologie (2): 13–16. Retrieved 3 March 2022.
- Wille, W. (1982). "Evolution and ecology of Upper Liassic dinoflagellates from southwest Germany". Neues Jahrbuch für Geologie und Paläontologie. 164 (1): 74–82. doi:10.1127/njgpa/164/1982/74.
- Wille, W. (1979). "Dinoflagellates from the Lias of SW Germany [Dinoflagellaten aus dem Lias Sмdwestdeutschlands]". Neues Jahrbuch für Geologie und Paläontologie. 158 (2): 221–258.
- Prauss, M.; Ligouis, B.; Luterbacher, H. (1991). "Organic matter and palynomorphs in the 'Posidonienschiefer'(Toarcian, Lower Jurassic) of southern Germany". Geological Society, London, Special Publications. 58 (1): 335–351. Bibcode:1991GSLSP..58..335P. doi:10.1144/GSL.SP.1991.058.01.21. S2CID 130285775. Retrieved 3 March 2022.
- Lund, J.J. (1996). "Jurassic and Cretaceous microfloras used to determine the stratigraphical succession of steeply dipping strata along the Prahl Fault, Dobenwohr Hafgraben, northeast Bavaria". Neues Jahrbuch für Geologie und Paläontologie. 200 (1): 133–147. doi:10.1127/njgpa/200/1996/133. Retrieved 3 March 2022.
- Gotch, H. (1964). "Planktonische Kleinformen aus dem Lias-Dogger Grenzbereich Nord- und Süddeutschlands". Neues Jahrb. Geol. Paläontol. 119 (3): 113–133. Retrieved 3 March 2022.
- Schouten, S.; van Kaam-Peters, H. M.; Rijpstra, W. I. C.; Schoell, M.; Damste, J. S. S. (2000). "Effects of an oceanic anoxic event on the stable carbon isotopic composition of early Toarcian carbon". American Journal of Science. 300 (1): 1–22. Bibcode:2000AmJS..300....1S. doi:10.2475/ajs.300.1.1. hdl:1874/4290. Retrieved 3 March 2022.
- Madler, K.A. (1963). "Organic microstructures of the Posidonia Shale. [ III. Die figurierten organischen Bestandteile der Posidonoenschiefer]". Beihefte zum Geologischen Jahrbuch. 58 (1): 287–406. Retrieved 13 October 2021.
- Visentin, S.; Erba, E.; Mutterlose, J. (2019). "Biostratigraphic constraints of the Early Toarcian Oceanic Anoxic Event: new data from calcareous nannofossil investigations of Boreal and Tethyan sections". PeerJ Preprints. 299 (23): 11. Retrieved 3 March 2022.
- Ebli, O.; Draxler, I.; Klein, P.; Kodina, L. A.; Lobitzler, H. (1991). "Fazies, Paläontologie und organische Geochemie der Sachranger Schiefer (Untertoarcium) im Mittelabschnitt der Nördlichen Kalkalpen zwischen Isar und Saalach" (PDF). Jahrbuch der Geologischen Bundesanstalt. 134 (1): 5–14. Retrieved 3 March 2022.
- Visentin, S. (2020). "Calcareous nannofossil biostratigraphy and taxonomy across the Early Toarcian Oceanic Anoxic Event: a comparison between Tethyan and Boreal sections". Tesi di dottorato. IRIS Institutional Research Information System: 1–202. Retrieved 2 October 2023.
- Grün, W.; Grün, P.; Prins, F. (1974). "Zweili Coccolithophoriden aus dem Lias epsilon von Holzmaden (Deutschland)". Neues Jahrb. Geol. Paläontol. Abh. 147 (2): 294–328.
- Visentin, S.; Faucher, G.; Mattioli, E.; Erba, E. (2021). "Taxonomic revision of the genus Carinolithus (Early-Middle Jurassic) based on morphometric analyses and diagenesis observations: Implications for biostratigraphy and evolutionary trends". Marine Micropaleontology. 162 (1): 412–432. Bibcode:2021MarMP.162j1950V. doi:10.1016/j.marmicro.2020.101950. hdl:2434/796274. S2CID 229440280. Retrieved 3 March 2022.
- Madler, K.A. (1969). "Tasmanites and related planktonic organisms from the Posidonian Shales. [ Tasmanites und verwandte planktonformen aus dem Posidonienschiefer-Meer. ]". International Conference on Planktonic Microfossils. 1 (23): 375–377. Retrieved 3 March 2022.
- Wetzel, O. (1958). "New microfossils from the Lias, especially from the Posidonian Shale. [ Neue mikrofossilien aus dem Lias, insbesondere aus dem Posidonienschiefer. ]". Paläontologische Zeitschrift. 32 (1): 15. Retrieved 3 March 2022.
- Galasso, F.; Schmid-Röhl, A.; Feist-Burkhardt, S.; Bernasconi, S. M.; Schneebeli-Hermann, E. (2021). "Changes in organic matter composition during the Toarcian Oceanic Anoxic Event (T-OAE) in the Posidonia Shale Formation from Dormettingen (SW-Germany)". Palaeogeography, Palaeoclimatology, Palaeoecology. 569 (1): 675–689. Bibcode:2021PPP...569k0327G. doi:10.1016/j.palaeo.2021.110327. S2CID 233550926. Retrieved 3 March 2022.
- Riegraf, W.; Werner, G.; Lörcher, F. (1984). Der Posidonienschiefer: Biostratigraphie, Fauna und Fazies des südwestdeutschen Untertoarciums (Lias e) (PDF). Berlin: F. Enke. ISBN 343294361X. Retrieved 20 February 2022.
- Rodríguez-Tovar, F. J. (2021). "Ichnology of the Toarcian Oceanic Anoxic Event: An understimated tool to assess palaeoenvironmental interpretations". Earth-Science Reviews. 216 (1): 122–146. Bibcode:2021ESRv..21603579R. doi:10.1016/j.earscirev.2021.103579. S2CID 233849558. Retrieved 3 March 2022.
- Kuhn, O.; Etter, W. (1994). "Der Posidonienschiefer der Nordschweiz: Lithostratigraphie, Biostratigraphie und Fazies". Eclogae Geologicae Helvetiae. 87 (1): 113–138. Retrieved 3 March 2022.
- Simpson, S. (1956). "On the trace-fossil Chondrites". Quarterly Journal of the Geological Society. 112 (3): 475–499. doi:10.1144/GSL.JGS.1956.112.01-04.23. S2CID 129365180. Retrieved 3 March 2022.
- Osgood, R. G. (1975). The history of invertebrate ichnology. In The study of trace fossils (pp. 3–12). Springer, Berlin, Heidelberg.
- Hertweck, G.; Wehrmann, A.; Liebezeit, G. (2007). "Bioturbation structures of polychaetes in modern shallow marine environments and their analogues to Chondrites group traces". Palaeogeography, Palaeoclimatology, Palaeoecology. 245 (4): 382–389. Bibcode:2007PPP...245..382H. doi:10.1016/j.palaeo.2006.09.001. Retrieved 3 March 2022.
- Baucon, A.; Bednarz, M.; Dufour, S.; Felletti, F.; Malgesini, G.; De Carvalho, C. N.; McIlroy, D. (2019). "Ethology of the trace fossil Chondrites: form, function and environment". Earth-Science Reviews. 202 (1): 223–238. doi:10.1016/j.earscirev.2019.102989. S2CID 210269603. Retrieved 3 March 2022.
- Van Acken, D.; Tütken, T.; Daly, J. S; Schmid-Röhl, A.; Orr, P. J. (2019). "Rhenium‑osmium geochronology of the Toarcian Posidonia Shale, SW Germany". Palaeogeography, Palaeoclimatology, Palaeoecology. 534 (2): 416–428. Bibcode:2019PPP...534j9294V. doi:10.1016/j.palaeo.2019.109294. S2CID 201318850. Retrieved 3 March 2022.
- Keighley, D. G.; Pickerill, R. K. (1995). "The ichnotaxa Palaeophycus and Planolites_ historical perspectives and recommendations". Ichnos. 33 (4): 566–572. doi:10.1080/10420949509386400. Retrieved 3 March 2022.
- Izumi, K. (2012). "Formation process of the trace fossil Phymatoderma granulata in the Lower Jurassic black shale (Posidonia Shale, southern Germany) and its paleoecological implications". Palaeogeography, Palaeoclimatology, Palaeoecology. 353 (2): 116–122. Bibcode:2012PPP...353..116I. doi:10.1016/j.palaeo.2012.07.021. Retrieved 3 March 2022.
- Izumi, K. (2013). "Geochemical composition of faecal pellets as an indicator of deposit-feeding strategies in the trace fossil Phymatoderma". Lethaia. 46 (4): 496–507. doi:10.1111/let.12028. Retrieved 3 March 2022.
- Savrda, C. E.; Bottjer, D. J. (1989). "Anatomy and Implications of Bioturbated Beds in "Black Shale" Sequences: Examples from the Jurassic Posidonienschiefer (Southern Germany)". PALAIOS. 4 (4): 330–341. Bibcode:1989Palai...4..330S. doi:10.2307/3514557. JSTOR 3514557. OSTI 7154351. Retrieved 3 March 2022.
- Krainer, K.; Mostler, Helfried; Haditsch, J. G. (1994). "Jurassische Bekkenbildung in den Nördlichen Kalkalpen bei Lofer (Salzburg) unter besonderer Berücksichtigung der Manganerz-Genese [Annotated record of the detailed examination of Mn deposits from the Northern Kalkalpen near Lofer (Salzburg, Austria)]" (PDF). Abhandlungen der Geologischen Bundesanstalt. 50 (1): 257–293. Retrieved 3 March 2022.
- Kotake, N. (1992). "Deep-sea echiurans: possible producers of Zoophycos". Lethaia. 25 (3): 311–316. doi:10.1111/j.1502-3931.1992.tb01400.x. Retrieved 2 April 2022.
- Röhl, H. J.; Schmid-Röhl, A.; Oschmann, W.; Frimmel, A.; Schwark, L. (2001). "The Posidonia Shale (Lower Toarcian) of SW-Germany: an oxygen-depleted ecosystem controlled by sea level and palaeoclimate". Palaeogeography, Palaeoclimatology, Palaeoecology. 165 (2): 27–52. Bibcode:2001PPP...165...27R. doi:10.1016/S0031-0182(00)00152-8. Retrieved 3 March 2022.
- Jefferies, R. P. S.; Minton, P. (1965). "The mode of life of two Jurassic species of Posidonia (Bivalvia)". Palaeontology. 8 (1): 156–185. Retrieved 3 March 2022.
- Hauff, B. (1921). "Untersuchung der Fossilfundstätten von Holzmaden im Posidonienschiefer des Oberen Lias Württembergs". Palaeontographica. 64 (1): 1–42. Retrieved 3 March 2022.
- Röhl, H. J. (1998). "Hochauflösende palökologische und sedimentologische Untersuchungen im Posidonienschiefer (Lias e)[epsilon)] von SW-Deutschland". Tübinger Geowissenschaftliche Arbeiten, Reihe A. 48 (1): 1–189. Retrieved 3 March 2022.
- Arp, G.; Gropengießer, S.; Schulbert, C.; Jung, D.; Reimer, A. (2021). "Biostratigraphy and sequence stratigraphy of the Toarcian Ludwigskanal section (Franconian Alb, Southern Germany)". Zitteliana. 95 (1): 95–57. doi:10.3897/zitteliana.95.56222. S2CID 237384737. Retrieved 3 March 2022.
- Lutikov, O.A.; Arp, G. (2020). "Revision Monotis substriata (Münster, 1831) and new species of bivalve molluscs in the Lower Toarcian in northern Russia and southern Germany (family Oxytomidae Ichikawa, 1958)". Jurassic System of Russia: Problems of Stratigraphy and Paleogeography. 2 (1): 125–131. Retrieved 3 March 2022.
- Lutikov, O. A.; Arp, G. (2022). "Taxonomy and Biostratigraphic Significance of the Toarcian Bivalves of the Genus Meleagrinella Whitfield, 1885". Stratigraphy and Geological Correlation. 30 (supplement issue 1): S47–S77. Bibcode:2022SGC....30S..47L. doi:10.1134/S0869593823010045. S2CID 257605092.
- Caswell, B. A.; Coe, A. L.; Cohen, A. S. (2009). "New range data for marine invertebrate species across the early Toarcian (Early Jurassic) mass extinction". Journal of the Geological Society. 166 (5): 859–872. Bibcode:2009JGSoc.166..859C. doi:10.1144/0016-76492008-0831. S2CID 140675044. Retrieved 3 March 2022.
- Ros-Franch, S.; Damborenea, S. E.; Márquez-Aliaga, A.; Manceñido, M. O. (2015). "Parainoceramya n. gen. for Parainoceramus Cox, 1954 (ex Voronetz, 1936) partim (Bivalvia, Jurassic)". Journal of Paleontology. 89 (1): 20–27. Bibcode:2015JPal...89...20R. doi:10.1017/jpa.2014.3. S2CID 86571525. Retrieved 3 March 2022.
- Hille, P. J. (2002). "De fossielen uit de Posidonienschiefer van Holzmaden en omgeving" (PDF). GEA. 35 (2): 8–17. Retrieved 3 March 2022.
- Teichert, S.; Nützel, A. (2015). "Early Jurassic anoxia triggered the evolution of the oldest holoplanktonic gastropod Coelodiscus minutus by means of heterochrony". Acta Palaeontologica Polonica. 60 (2): 269–276. Retrieved 3 March 2022.
- Bandel, K.; Hemleben, C. (1987). "Jurassic heteropods and their modern counterparts (planktonic Gastropoda, Mollusca)". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 174 (2): 1–22. Retrieved 3 March 2022.
- Brosamlen, R. (1909). "Beitrag zur Kenntnis der Gastropoden des schwäibischen Jura". Palaeontographica. 56 (1): 177–321.
- Maisch, Michael W. (2021). "Neubewertung der Ammonitenfauna der Posidonienschiefer-Formation (Unterjura, Toarcium) von Baden-Württemberg, Südwestdeutschland". Jahreshefte der Gesellschaft für Naturkunde in Württemberg. 177 (1): 265–347. doi:10.26251/jhgfn.177.2021.265-347. Retrieved 2 March 2022.
- Hille, P. J. (2002). "De fossielen uit de Posidonienschiefer van Holzmaden en omgeving" (PDF). GEA. 35 (2): 8–17. Retrieved 2 March 2022.
- Jacobshagen, V. (1965). "Die Allgäu-Schichten (Jura-Fleckenmergel) zwischen Wettersteingebirge und Rhein" (PDF). Geol. Bundesanstalt. 108 (1): 1–114. Retrieved 2 March 2022.
- Delsate, D.; Weis, R. (2010). "La Couche à Crassum (Toarcien moyen) au Luxembourg: stratigraphie et faunes de la coupe de Dudelange-Zoufftgen". Ferrantia. 62 (2): 35–62. Retrieved 2 March 2022.
- Riegraf, W. (1980). "Revision der Belemniten des Schwäbischen Jura". Palaeontographica. 169 (2): 128–206. Retrieved 2 March 2022.
- Schlegelmilch, R. (1998). "Formenkundlicher Teil. In Die Belemniten des süddeutschen Jura". Spektrum Akademischer Verlag, Heidelberg. 87 (1): 39–89. doi:10.1007/978-3-8274-3083-0. ISBN 978-3-8274-3082-3. Retrieved 2 March 2022.
- Tripp, K. (1938). "Der Stammbaum der Belemniten des Lias Schwabens". Paläontologische Zeitschrift. 19 (4): 180–198. doi:10.1007/BF03042240. S2CID 129424006. Retrieved 2 March 2022.
- Schulbert, C. (2001). Die Ammonitenfauna und Stratigraphie der Tongrube Mistelgau bei Bayreuth (Oberfranken) (4 ed.). Berlin: Digital Druck AG. Retrieved 2 March 2022.
- Pompeckj, J. (1901). "Der Jura zwischen Regensburg und Regenstauf: Geognostische Jahreshefte" (PDF). Bayerischen Staatsministeriums des Innern. 14 (3): 1–290. Retrieved 2 March 2022.
- Landman, N. H.; Saunders, W. B.; Winston, J. E; Harries, P. J. (2010). "Incidence and Kinds of Epizoans on the Shells of Live Nautilus". Nautilus. 2 (1): 163–177. doi:10.1007/978-90-481-3299-7_10. Retrieved 2 March 2022.
- Fischer, K. C.; KC, F. (1981). "Chitinobelus acifer ngn sp., ein ungewöhnlicher Belemnit aus dem Lias epsilon von Holzmaden". Neues Jahrbuch für Geologie und Paläontologie - Monatshefte. 78 (1): 141–148. doi:10.1127/njgpm/1981/1981/141. Retrieved 2 March 2022.
- Bode, A. (1933). "Chondroteuthis wunnenbergi ngn sp., eine neue Belemnoideenform, in gunstiger Erhaltung". Sonderabdruck aus dem 25. Jahresbericht des Niedersachsischen geologischen Vereins zu Hannover (Geologische Abteilung der Naturhistorischen Gesellschaft zu Hannover). 25 (1): 33–66.
- Fuchs, D.; Donovan, D. T.; Keupp, H. (2013). "Taxonomic revision of "Onychoteuthis" conocauda Quenstedt, 1849 (Cephalopoda: Coleoidea)". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 270 (3): 245–255. doi:10.1127/0077-7749/2013/0368. Retrieved 2 March 2022.
- Neumeister, S.; Gratzer, R.; Algeo, T. J.; Bechtel, A.; Gawlick, H. J.; Newton, R. J.; Sachsenhofer, R. F. (2015). "Oceanic response to Pliensbachian and Toarcian magmatic events: Implications from an organic-rich basinal succession in the NW Tethys". Global and Planetary Change. 126 (3): 62–83. Bibcode:2015GPC...126...62N. doi:10.1016/j.gloplacha.2015.01.007. Retrieved 2 March 2022.
- Mutterlose, J.; Klopschar, M.; Visentin, S. (2022). "Ecological Adaptation of Marine Floras and Faunas Across the Early Jurassic Toarcian Oceanic Anoxic Event–a Case Study from Northern Germany". SSRN. 34 (1): 1–43. SSRN 4039594. Retrieved 2 March 2022.
- Von Michael, W. Maisch (2018). "Neue Dactylioceratiden (Cephalopoda, Ammonitina) aus dem untersten Toarcium (Tenuicostatum-Zone) von Baden-Württemberg, Südwestdeutschland Jh". Ges. Naturkde. Württemberg174Stuttgart. 174 (1): 143–172. Retrieved 2 March 2022.
- Weiß, K.P.; Freytag, D. (1991). "Frechiella subcarinata (Young & Bird, 1822), ein bemerkenswerter Ammonit aus dem höheren Untertoarcium (bifrons-Zone) der Fränkischen Alb". Geologische Blätter für Nordost-Bayern. 41 (2): 125–132.
- Fuchs, D.; Weis, R. (2008). "Taxonomy, morphology and phylogeny of Lower Jurassic loligosepiid coleoids (Cephalopoda)". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 249 (1): 93–112. doi:10.1127/0077-7749/2008/0249-0093. Retrieved 2 March 2022.
- Riegraf, W. (1997). "On the proposed conservation of the names Geopeltis (Regteren Altena, 1949), Geoteuthis (Muenster, 1843), Jeletzkyteuthis (Doyle, 1990), Loligosepia (Quenstedt, 1839), Parabelopeltis (Naef, 1921), Paraplesioteuthis (Naef, 1921) and Belemnotheutis montefiorei (Buckman, 1880) (Mollusca, Coleoidea)". The Bulletin of Zoological Nomenclature. 54 (2): 184. doi:10.5962/bhl.part.105. Retrieved 2 March 2022.
- Wellnhofer, P.; Vahldiek, B.W. (1986). "Ein Flugsaurier-Rest aus dem Posidonienschiefer (Unter-Toarcium) von Schandelah bei Braunschweig". Paläontologische Zeitschrift. 60 (1): 329–340. doi:10.1007/BF02985677. S2CID 129838740. Retrieved 10 February 2022.
- Beyermann, K.; Hasenmaier, D. (1977). "Identification of 180 million years old, probably unchanged melanine". NASA Technical Reports Server. 21 (1): 67–72. Retrieved 2 March 2022.
- Keupp, H.; Kohring, R. (1993). "Ein Magensteinfund aus dem Lias Epsilon von Altdorf (Mittelfranken)". Geologische Blätter für Nordost-Bayern und angrenzende Gebiete. 43 (3): 95–104.
- Riegraf, W. (1987). "On Lower and Upper Jurassic dibranchiate cephalopods from Germany and England" (PDF). Paläontologische Zeitschrift. 61 (1): 261–272. doi:10.1007/BF02985908. S2CID 129847245. Retrieved 2 March 2022.
- Fuchs, D.; Keupp, H.; Schweigert, G. (2013). "First record of a complete arm crown of the Early Jurassic coleoid Loligosepia (Cephalopoda)". Paläontologische Zeitschrift. 87 (3): 431–435. doi:10.1007/s12542-013-0182-4. S2CID 129112888. Retrieved 2 March 2022.
- Doguzhaeva, L. A; Mutvei, H. (2003). "Gladius composition and ultrastructure in extinct squid-like coleoids: Loligosepia, Trachyteuthis and Teudopsis". Revue de Paléobiologie. 22 (2): 877–894.
- Maisch, M. W.; Hoffmann, R. (2017). "Lytoceratids (Cephalopoda, Ammonoidea) from the Lower Posidonienschiefer Formation (Tenuicostatum Zone, Early Jurassic) of Baden-Württemberg (south-western Germany)". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 283 (3): 275–289. doi:10.1127/njgpa/2017/0640. Retrieved 2 March 2022.
- Münster, G.; Graf, Z. (1843). "Die schalenlosen Cephalopoden im unteren Jura, den Lias-Schiefern von Franken und Schwaben". Beiträge zur Petrefaktenkunde. 6 (2): 57–77.
- Riegraf, W.; Reitner, J. (1979). "Die" Weichteilbelemniten" des Posidonienschiefers (Untertoarcium) von Holzmaden (Baden-Württemberg) sind Fälschungen". Neues Jahrbuch für Geologie und Paläontologie. 56 (5): 291–304. Retrieved 2 March 2022.
- Schulbert, C. (2001). "Die Ammonitenfauna und Stratigraphie der Tongrube Mistelgau bei Bayreuth (Oberfranken)" (PDF). Beihefte zu den Berichten der Naturwissenschaftlichen Gesellschaft Bayreuth. 4 (1): 1–183. Retrieved 2 March 2022.
- Reitner, J.; Engeser, T. (1982). "Zwei neue Coleoidea-Arten aus dem Posidonienschiefer (Untertoarcium) aus der Gegend von Holzmaden (Baden-Württemberg)". Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie). 84 (19): 231–242. Retrieved 2 March 2022.
- Fuchs, D.; Weis, R. (2010). "Taxonomy, morphology and phylogeny of Lower Jurassic teudopseid coleoids (Cephalopoda)". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 257 (3): 351–366. doi:10.1127/0077-7749/2010/0083. Retrieved 2 March 2022.
- Schweigert, G. (2007). "Juracyclus posidoniae n. gen. and sp., the first cycloid arthropod from the Jurassic". Journal of Paleontology. 81 (1): 213–215. doi:10.1666/0022-3360(2007)81[213:JPNGAS]2.0.CO;2. S2CID 131620349. Retrieved 2 March 2022.
- Knitter, H. (1983). "Biostratigraphische Untersuchungen mit Ostracoden im Toarcien Süddentschlands Facies". Erlangen. 8 (2): 213–262.
- Knitter, H.; Riegraf, W. (1984). "Biostratigraphie (Cephalopoden, Ostracoden ) des Oberen Toarcium von Blumberg Achdorf, Wutachs und Weilheim/Teck (Baden-Württemberg)". Jahrbuch der Geologischen Landesamt Baden-Württemberg. 26 (2): 57–97. Retrieved 2 March 2022.
- Malz, H. (1975). "Eine Entwicklungsreihe "vallater" Ogmoconchen (Ostracoda) im S-deutschen Lias". Senckenbergiana Lethaea. 55 (6): 485–505.
- Malz Zur Kenntnis, H. (1966). "Ostracoden-Arten der Gattungen Kinkelinella und Praeschuleridea". Senckenbergiana Lethaea. 47 (4): 385–404.
- Fischer, W. (1961). "Neue Arten Ostracoden-Gattung Polycope SARS (1865) aus dem Oberen Lias (Württemberg)". Neues Jahrbuch für Geologie und Paläontologie. 8 (2): 497–501.
- Förster, R. (1967). "Zur Kenntnis natanter Jura-Dekapoden" (PDF). Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie. 7 (3): 157–174. Retrieved 2 March 2022.
- Haug, J. T.; Haug, C.; Schweigert, G. (2019). "The oldest "intermetamorphic" larva of an achelatan lobster from the Lower Jurassic Posidonia Shale, South Germany" (PDF). Acta Palaeontologica Polonica. 64 (4): 685–692. doi:10.4202/app.00627.2019. S2CID 207820936. Retrieved 2 March 2022.
- Ilger, J.M. (2014). "Insekten und andere Gliederfüßer". _niedersachsens Versunkene Urwelt. 1 (1): 10–18.
- Audo, D. (2016). "Tonneleryon, a new gregarious polychelidan lobster from the early Toarcian Posidonia Shale of Holzmaden (Germany)". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 280 (3): 285–298. doi:10.1127/njgpa/2016/0580. Retrieved 2 March 2022.
- Beurlen, K. (1930). "Nachträge zur Decapodenfauna des schwäbischen Jura. I. Neue Decapodenfunde aus dem Posidonienschiefer von Holzmaden" (PDF). Neues Jahrbuch für Mineralogie, Geologie und Paläontologie. 64 (3): 219–234. Retrieved 2 March 2022.
- Devillez, J.; Charbonnier, S. (2019). "Review of the Early and Middle Jurassic erymid lobsters (Crustacea: Decapoda) Révision des Érymides (Crustacea: Decapoda) du Jurassique inférieur et moyen". Bulletin de la Société Géologique de France. 190 (1): 6–21. doi:10.1051/BSGF/2019005. S2CID 196643332. Retrieved 2 March 2022.
- Audo, D.; Williams, M.; Charbonnier, S.; Schweigert, G. (2017). "Gabaleryon, a new genus of widespread early Toarcian polychelidan lobsters". Journal of Systematic Palaeontology. 15 (3): 205–222. doi:10.1080/14772019.2016.1167786. S2CID 87613086. Retrieved 2 March 2022.
- Aud, D.; Haug, J. T.; Haug, C.; Charbonnier, S.; Schweigert, G.; Müller, C. H.; Harzsch, S. (2016). "On the sighted ancestry of blindness–exceptionally preserved eyes of Mesozoic polychelidan lobsters". Zoological Letters. 2 (1): 13. doi:10.1186/s40851-016-0049-0. PMC 4947519. PMID 27429789.
- Schweigert, G. (2018). "Neufund von Glypheopsis grandichela im Posidonienschiefer". Fossilien. 35 (6): 56–57.
- Frentzen, K. (1937). "Paläontologische Skizzen aus den Badischen Landessamlungen für Naturkunde, Karlsruhe i. Br. II. Mecochirus eckerti nov. spec. aus dem Lias Epsilon (Posidonienschiefer) von Langenbrücken". Carolinea. 2 (2): 103–105.
- Schweigert, G.; Röper, M. (2001). "Neue Krebse der Gattung Palaeastacus (Crustacea: Decapoda: Erymidae) aus ober jurassischen Plattenkalken Süddeutschlands" (PDF). Staatliches Museum für Naturkunde. 313 (5): 1–12. Retrieved 2 March 2022.
- Seilacher, A.; Reif, W. E.; Westphal, F. (1985). "Sedimentological, ecological and temporal patterns of fossil Lagerstätten". Philosophical Transactions of the Royal Society of London. 311 (11): 5–24. JSTOR 2396966.
- Beurlen, K. (1944). "Neue Reste von Proeryon (Crustacea Decapoda, Eryonidea)". Neues Jahrb Miner Geolog Paläont. 88 (1): 374–384.
- Audo, D.; Schweigert, G.; Charbonnier, S. (2019). "Proeryon, a geographically and stratigraphically widespread genus of polychelidan lobsters". Annales de Paléontologie. 106 (2): 102–118. Bibcode:2020AnPal.10602376A. doi:10.1016/j.annpal.2019.102376. S2CID 213469487. Retrieved 2 March 2022.
- Beurlen, K. (1928). "Die Decapoden des Schwäbischen Jura mit Ausnahme der aus den oberjurassischen Plattenkalken stammenden". Palaeontographica. 70 (3): 115–278. Retrieved 2 March 2022.
- Schweigert, G. (2003). "The lobster genus Uncina Quenstedt, 1851 (Crustacea: Decapoda: Astacidea: Uncinidae) form the Lower Jurassic". Stuttgarter Beiträge zur Naturkunde. 332 (4): 1–43.
- Bode, A. (1951). "Ein liassischer Scorpionide". Paläontologische Zeitschrift. 24 (2): 58–65. doi:10.1007/BF03044552. S2CID 140650635. Retrieved 2 March 2022.
- Dunlop, J. A.; Kamenz, C.; Scholtz, G. (2007). "Reinterpreting the morphology of the Jurassic scorpion Liassoscorpionides". Arthropod Structure & Development. 36 (2): 245–252. doi:10.1016/j.asd.2006.09.003. PMID 18089103. Retrieved 2 March 2022.
- Ansorge, J. (2003). "Insects from the Lower Toarcian of Middle Europe and England". Proceedings of the Second Palaeoentomological Congress, Acta Zoologica Cracoviensia. 46 (1): 291–310. Retrieved 30 July 2021.
- Bode, A. (1953). "Die Insektenfauna des Ostniedersachsischen Oberen Lias" (PDF). Palaeontographica Abteilung A. 103 (1): 1–375. Retrieved 2 March 2022.
- Bode, A. (1905). "Orthoptera and Neuroptera from the Upper Lias of Braunschweig". Yearbook of the Royal Prussian State Geological Institute and Mining Academy in Berlin. 25 (1): 218–245.
- Henrotay, M.; Nel, A.; Jarzembowski, E. A. (1997). "New Protomyrmeleontid Damselflies from the Triassic of Australia and the Liassic of Luxembourg, with the description of Tillyardomyrmeleon petermilleri gen. nov. & spec. nov. (Archizygoptera: Protomyrmeleontidae)". Odonatologica. 26 (4): 395–404. Retrieved 2 March 2022.
- Nel, A.; Henrotay, M. (1992). "Les Protomyrmeleontidae (Odonatoptera, Odonata, Archizygoptera stat. rest.): état actuel des connaissances". Annales de Paléontologie. 78 (2): 1–47.
- Ansorge, J. (1999). "Heterophlebia buckmani (Brodie 1845) (Odonata: "Anisozygoptera") - das erste Insekt aus dem untertoarcischen Posidonienschiefer von Holzmaden (Württemberg, SW Deutschland)". Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie). 275 (1): 1–9. Retrieved 2 March 2022.
- Etter, W.; Kuhn, O. (2000). "An articulated dragonfly (Insecta, Odonata) from the Upper Liassic Posidonia Shale of Northern Switzerland". Palaeontology. 43 (3): 967–977. Bibcode:2000Palgy..43..967E. doi:10.1111/1475-4983.00157. S2CID 140165815. Retrieved 2 March 2022.
- Nel, A.; Martínez-Delclòs, X.; Paicheler, J. C.; Henrotay, M. (1993). "Les "Anisozygoptera" fossiles Phylogenie et classification (Odonata)". Martinia. 3 (4): 1–311. Retrieved 2 March 2022.
- Bechly, G. (2018). "First record and a new species of the fossil dragonfly genus Proinogomphus (Odonata: Liassogomphidae) from the Early Jurassic of Bascharage in the Grand Duchy of Luxembourg". Zootaxa. 4450 (2): 108–114. doi:10.11646/zootaxa.4450.1.7. PMID 30313860. S2CID 52977117. Retrieved 2 March 2022.
- Handlirsch, A. (1939). "Neue Untersuchungen über die fossilen Insekten mit Ergänzungen und Nachträgen sowie Ausblicken auf phylogenetische, palaeogeographische und allgemein biologische Probleme". Annalen des Naturhistorischen Museums in Wien. 2 (3): 1–240. JSTOR 41768379.
- Ansorge, J.; Reich, M. (2018). "Komplette Libelle Sphenophlebia pommerana" (PDF). Fossilien. 35 (1): 60–61. Retrieved 2 March 2022.
- Nel, A.; Bechly, G.; Delclòs, X.; Huang, D. (2009). "New and poorly known Mesozoic damsel-dragonflies (Odonata: Isophlebioidea: Campterophlebiidae, Isophlebiidae)". Palaeodiversity. 2 (4): 209–232. Retrieved 2 March 2022.
- Nel, A.; Weis, R. (2017). "A new Early Jurassic damselfly from the Grand Duchy of Luxembourg (Odonata: Campterophlebiidae)". Alcheringa. 41 (5): 378–382. doi:10.1080/03115518.2017.1289417. S2CID 132602020. Retrieved 2 March 2022.
- Fleck, G.; Bechly, G.; Martinez-Delclos, X.; Jarzembowski, E.; Coram, R.; Nel, A. (2003). "Phylogeny and classification of the Stenophlebioptera (Odonata: Epiproctophora)". Annales de la Société Entomologique de France. 39 (1): 55–93. doi:10.1080/00379271.2003.10697363. S2CID 85417279. Retrieved 2 March 2022.
- Brachert, T.H. (1987). "Makrofossilführung der "Siemensi-Geoden" (Mittlerer Lias Epsilon, Unteres Toarcium) von Kerkhofen/Oberpfalz (Bayern): Neue Insekten- und Pflanzenfunde". Geologische Blätter NO-Bayern. 37 (4): 217–240.
- Berger, G. (1989). "Über Insektenfunde beim Kanalbau". Fossilien. 6 (1): 44–47.
- Ansorge, J. (1996). "Zur systematischen Position von Schesslitziella haupti Kuhn 1952 (Insecta: Phasmatodea) aus dem Oberen Lias von Nordfranken (Deutschland)". Paläontologische Zeitschrift. 70 (4): 475–479. doi:10.1007/BF02988086. Retrieved 2 March 2022.
- Vršanský, P.; Ansorge, J. (2007). "Lower Jurassic cockroaches (Insecta: Blattaria ) from Germany and England". African Invertebrates. 48 (1): 103–126. Retrieved 23 October 2021.
- Szwedo, J.; R., Weis; Nel, A. (2017). "A bizarre sternorrhynchan wing from the Lower Jurassic of Luxembourg (Hemiptera: Sternorrhyncha: Pincombeomorpha?)". Historical Biology. 31 (3): 806–812. doi:10.1080/08912963.2017.1395423. S2CID 90106633. Retrieved 2 March 2022.
- Szwedo, J. (2011). "The Coleorrhyncha (Insecta: Hemiptera) of the European Jurassic, with a description of a new genus from the Toarcian of Luxembourg". Volumina Jurassica. 9 (2): 3–20. Retrieved 2 March 2022.
- Rasnitsyn, A. P.; Ansorge, J.; Zessin, W. (2003). "New hymenopterous insects (Insecta: Hymenoptera) from the lower Toarcian (Lower Jurassic) of Germany". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 227 (1–3): 321–342. doi:10.1127/njgpa/227/2003/321.
- Zessin, W. (1985). "Neue oberliassische Apocrita und die Phylogenie der Hymenoptera". Deutsche Entomologische Zeitschrift. 32 (3): 129–142. doi:10.1002/mmnd.19850320118. Retrieved 2 March 2022.
- Willmann, R. (1994). "Raphidiodea aus dem Lias und die Phylogenie der Kamelhalsfliegen (Insecta: Holometabola)". Paläontologische Zeitschrift. 68 (2): 167–197. doi:10.1007/BF02989439. S2CID 128926273. Retrieved 2 March 2022.
- Nel, A.; Petrulevicius, J. F.; Henrotay., M. (2004). "New Early Jurassic sawflies from Luxembourg: the oldest record of Tenthredinoidea (Hymenoptera: "Symphyta")". Acta Palaeontologica Polonica. 49 (2): 283–288. Retrieved 2 March 2022.
- Ponomarenko, A.G. (1995). "Upper Liassic neuropterans (Insecta) from Lower Saxony, Germany". Russian Entomological Journal. 4 (4): 73–89.
- Ansorge, J.; Makarkin, V. N. (2020). "The oldest giant lacewings (Neuroptera: Kalligrammatidae) from the Lower Jurassic of Germany". Palaeoworld. 30 (2): 296–310. doi:10.1016/j.palwor.2020.07.001. S2CID 225633800. Retrieved 2 March 2022.
- Nel, A.; Henrotay, M. (1994). "Les Chrysopidae Mésozoïques. État actuel des connaissances. Description d'un nouveau genre et nouvelle espèce dans le Jurassique inférieur (Lias) (Insecta: Neuroptera)". Annales de la Société Entomologique de France. 30 (2): 295–318. Retrieved 2 March 2022.
- Nel, A. (1996). "Un Tettigarctidae fossile du Lias européen (Cicadomorpha, Cicadoidea, Tettigarctidae)". École Pratique des Hautes Études, Biologie et Évolution des Insectes. 9 (6): 83–94.
- Lukashevich, E.; Ansorge, J.; Krzemiński, W.; Krzemińska, E. (1998). "Revision of Eoptychopterinae (Diptera: Eoptychopteridae)". Polskie Pismo Entomologiczne. 67 (1): 311–343. Retrieved 23 October 2021.
- Ansorge, J. (2003). "Upper Liassic Amphiesmenopterans (Trichoptera+ Lepidoptera) from Germany–a review". Acta Zoologica Cracoviensia. 46 (3): 285–290. S2CID 55218822. Retrieved 2 March 2022.
- Kuhn O. (1951): Ein vermutlicher Schmetterling, Geisfeldiella benkerti n.g.n.sp. aus dem Lias I Nordfrankens. Neues Jahrbuch Geologie Paläontologie Mh. 1 951: 58–61
- Kopeć, K.; Soszyńska-Maj, A.; Gehler, A.; Ansorge, J.; Krzemiński, W. (2018). "Mecoptera and Diptera from the early Toarcian (Early Jurassic) deposits of Wolfsburg–Große Kley (Lower Saxony, Germany)". Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 107 (3): 163–171. doi:10.1017/S1755691017000226. S2CID 134568600. Retrieved 2 March 2022.
- Ansorge, J. (2001). "Lower Jurassic Hennigmatidae (Diptera) from Germany". Studia dipterologica. 8 (1): 97–102. Retrieved 23 October 2021.
- Khramov, A. V.; Bashkuev, A. S.; Lukashevich, E. D. (2020). "The Fossil Record of Long-Proboscid Nectarivorous Insects". Entomological Review. 100 (7): 881–968. doi:10.1134/S0013873820070015. S2CID 234675037. Retrieved 2 March 2022.
- Ansorge, J. (1994). "Tanyderidae and Psychodidae (Insecta: Diptera) from the Lower Jurassic of northeastern Germany". Paläontologische Zeitschrift. 68 (1): 199–210. doi:10.1007/BF02989440. S2CID 128958459. Retrieved 23 October 2021.
- Thuy, B.; Numberger-Thuy, L. D. (2021). "Brittlestar diversity at the dawn of the Jenkyns Event (early Toarcian Oceanic Anoxic Event): new microfossils from the Dudelange drill core, Luxembourg". Geological Society, London, Special Publications. 514 (1): 34–58. Bibcode:2021GSLSP.514...83T. doi:10.1144/SP514-2021-3. S2CID 234842742. Retrieved 20 February 2022.
- Kutscher, M. (1992). "Ophiomusium geisingense n.sp. eine neue Ophiurenart aus dem Lias Epsilon (Unteres Toarcium) von Bachhausen/Bayern". Archaeopteryx. 10 (1): 25–30. Retrieved 20 February 2022.
- Simms, M. J. (1988). "An intact comatulid crinoid from the Toarcian of Southern Germany" (PDF). Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie). 140 (1): 1–7. Retrieved 20 February 2022.
- Hess, H. (1991). "Neue Schlangensterne aus dem Toarcium und Aalenium des Schwäbischen Jura (Baden-Württemberg)". Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie). 180 (2): 1–11. OCLC 28384653. Retrieved 20 February 2022.
- Gall, J. C. (1983). Ancient Sedimentary Environments and the Habitats of Living Organisms: The Holzmaden Bituminous Shale Sea. Berlin: Springer. pp. 158–166. doi:10.1007/978-3-642-68909-3. ISBN 978-3-642-68911-6. Retrieved 20 February 2022.
- Hauff, R. B. (1984). "Pentacrinites quenstedti (Oppel) aus dem oberen Untertoarcium (Lias Epsilon) von Ohmden bei Holzmaden (SW-Deutschland)". Paläontologische Zeitschrift. 58 (1): 255–263. doi:10.1007/BF02986064. S2CID 128699904. Retrieved 20 February 2022.
- Jäger, M. (1995). "Echinodermata aus dem Ober-Toarcium und Aalenium Deutschlands I. Crinoidea: Cyrtocrinina und Millericrinina" (PDF). Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie). 226 (1): 1–51. Retrieved 20 February 2022.
- Seilacher, A.; Drozdzewski, G.; Haude, R. (1968). "Form and function of the stem in a pseudoplanktonic crinoid (Seirocrinus)" (PDF). Palaeontology. 11 (2): 275–282. Retrieved 20 February 2022.
- Simms (1989). "Contrasting lifestyles in lower Jurassic crinoids: A comparison of benthic and pseudopelagic Isocrinida". Palaeontology. 29 (2): 475–493. Retrieved 20 February 2022.
- Haude, R.; Jangoux, M. (1980). "Constructional morphology of the stems of Pentacrinitidae, and mode of life of Seirocrinus". Proceedings of the European Colloquium on Echinoderms, Brussels. AA Balkema. 1 (2): 17–23. doi:10.1201/9781003078913-3. S2CID 225515238. Retrieved 20 February 2022.
- Fraas, E. (1910). "Chimäridenreste aus dem oberen Lias von Holzmaden" (PDF). Jahreshefte des Vereins für Vaterländische Naturkunde in Württemberg. 66 (2): 55–63. Retrieved 19 February 2022.
- Duffin, C. J. (1995). "Holocephalans in the Staatliches Museum für Naturkunde in Stuttgart 3. First chimaeroid from the Lias of Baden-Württemberg (Early Toarcian of Ohmden)" (PDF). Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie). 231 (4): 1–12. Retrieved 19 February 2022.
- Quenstedt, F. A. (1882). "Bdellodus bollensis aus dem Posidonienschiefer von Boll". Jahreshefte des Vereins für Vaterländische Naturkunde in Württemberg. 38 (2): 132–142.
- Maisch, M. W.; Matzke, A. T. (2016). "A new hybodontid shark (Chondrichthyes, Hybodontiformes) from the Lower Jurassic Posidonienschiefer Formation of Dotternhausen, SW Germany". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 280 (1): 241–257. doi:10.1127/njgpa/2016/0577. Retrieved 19 February 2022.
- Fraas, E. (1895). "Ein Fund von Skeletresten von Hybodus (Hybodus hauffianus)". Jahresberichte und Mitteilungen des Oberrheinischen Geologischen Vereins. 28 (3): 24–26.
- Fraas, E. (1896). "Neue Selachier-Reste aus dem oberen Lias von Holzmaden in Württemberg". Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg. 52 (1): 1–25. Retrieved 19 February 2022.
- Duffin, C. J. (1997). "The dentition of Hybodus hauffianus Faas, 1895 (Toarcian, Early Jurassic)". Stuttgarter Beiträge zur Naturkunde. 256 (2): 1–20.
- Duffin, C. J. (1983). "Holocephalans in the Staatliches Museum für Naturkunde in Stuttgart. 1. Myriacanthoids and squalorajoids" (PDF). Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie). 97 (2): 1–41. Retrieved 19 February 2022.
- Reif, W. E. (1974). "Metopacanthus sp.(Holocephali) und Palaeospinax egertoni S. Woodward (Selachii) aus dem unteren Toarcium von Holzmaden. Staatl. Museum für Naturkunde" (PDF). Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie). 10: 1–9. Retrieved 19 February 2022.
- Maisey (1977). "The fossil selachian fishes Palaeospinax Egerton, 1872 and Nemacanthus Agassiz, 1837". Zoological Journal of the Linnean Society. 60 (3): 259–273. doi:10.1111/j.1096-3642.1977.tb01029.x. Retrieved 19 February 2022.
- Klug, S.; Kriwet, J. (2008). "A new basal galeomorph shark (Synechodontiformes, Neoselachii) from the Early Jurassic of Europe". Naturwissenschaften. 95 (5): 443–448. Bibcode:2008NW.....95..443K. doi:10.1007/s00114-007-0341-0. PMID 18196213. S2CID 8460659. Retrieved 19 February 2022.
- Thies, D. (1992). "A new species of Palaeospinax (Chondrichthyes, Neoselachii) from the Lower Jurassic Posidonia Shale of southern Germany". Paläontologische Zeitschrift. 66 (1): 137–146. doi:10.1007/BF02989484. S2CID 128557361. Retrieved 19 February 2022.
- Maisey, J. G.; Ehret, D. J.; Denton, J. S. (2020). "A new genus of Late Cretaceous angel shark (Elasmobranchii; Squatinidae), with comments on squatinid phylogeny". American Museum Novitates (2020): 1–29. doi:10.1206/3954.1. hdl:2246/7230. S2CID 219700378. Retrieved 19 February 2022.
- Duffin, C. J.; Joiko, L. (2020). "A fin spine of Recurvacanthus (Myriacanthidae, Holocephali) from the Posidonienschiefer (Early Jurassic) of SW Germany". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 296 (3): 317–326. doi:10.1127/njgpa/2020/0908. S2CID 225844997. Retrieved 19 February 2022.
- White, E. I. (1925). "LXIX.—Additions to the upper Liassic fish-fauna of Holzmaden". Annals and Magazine of Natural History. 15 (9): 601–611. doi:10.1080/00222932508633256. Retrieved 19 February 2022.
- Woodward, A. S. (1893). "On the cranial osteology of the Mesozoic ganoid fishes, Lepidotus and Dapedius". Proceedings of the Zoological Society of London. 38 (3): 559–565.
- Thies, D.; Waschkewitz, J. (2016). "Redescription of Dapedium pholidotum (Agassiz, 1832)(Actinopterygii, Neopterygii) from the Lower Jurassic Posidonia Shale, with comments on the phylogenetic position of Dapedium Leach, 1822". Journal of Systematic Palaeontology. 14 (4): 339–364. doi:10.1080/14772019.2015.1043361. S2CID 130282395. Retrieved 19 February 2022.
- Thies, D.; Hauff, R. B. (2011). "A new species of Dapedium Leach, 1822 (Actinopterygii, Neopterygii, Semionotiformes) from the Early Jurassic of South Germany" (PDF). Palaeodiversity. 4 (3): 185–221. Retrieved 19 February 2022.
- Bürgin, T. (2000). "Euthynotus cf. incognitus (Actinopterygii, Pachycormidae) als Mageninhalt eines Fischsauriers aus dem Posidonienschiefer Süddeutchlands (Unterer Jura, Lias epsilon)". Eclogae Geologicae Helvetiae. 93 (1): 491–496.
- Cooper, Samuel L. A.; Giles, Sam; Young, Holly; Maxwell, Erin E. (December 2022). "A New Large †Pachycormiform (Teleosteomorpha: †Pachycormiformes) from the Lower Jurassic of Germany, with Affinities to the Suspension-Feeding Clade, and Comments on the Gastrointestinal Anatomy of Pachycormid Fishes". Diversity. 14 (12): 1026. doi:10.3390/d14121026. ISSN 1424-2818.
- Delsate, D. (1999). "Haasichthys michelsi, nov. gen., nov. sp., un nouveau Pachycormiforme (Osteichthyes, Actinopterygii) du Toarcian inférieur (Jurassique) luxembourgeois". Trav. Sci. Mus. Nat. Hist. Nat. Lux. 32: 87–140.
- Ebert, M.; Thies, D.; Hauff, R. B. (2020). "First evidence of ganoin-scaled Halecomorphi (Neopterygii) in the Lower Jurassic of Holzmaden and Ohmden, Germany". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 295 (3): 307–326. doi:10.1127/njgpa/2020/0889. S2CID 216427872. Retrieved 19 February 2022.
- Jäger, Manfred (2005). Das Fossilienmuseum im Werkforum. Dotternhausen: Führer durch die Ausstellung von Jura-Fosslilien. Retrieved 3 March 2022.
- Henrotay, M.; Marques, D.; Paicheler, J.C.; Gall, J.C.; Nel, A. (1998). "Le Toarcien inférieur des régions de Bascharage et de Bettembourg (grand-duché du Luxembourg): évidences paléontologiques et sédimentologiques d'environnements restreints proches de l'émersion" (PDF). Geodiversitas. 20 (2): 263–284. Retrieved 3 March 2022.
- Wunnenberg, C. (1950). "Zur Ausbildung des Posidonienschiefers in der Umgebung von Braunschweig mit besonderer Berucksichtigung der Fossilisation: Neües Jahrbuch fur Geologie und Palaontologie Monatshefte". Paläontologische Zeitschrift. 46 (1): 161–182.
- Woodward, A. S. (1895). Catalogue of the fossil fishes in the British Museum (Natural History). London: British Museum. p. 112. Retrieved 19 February 2022.
- Arratia, G.; Thies, D. (2001). "A new teleost (Osteichthyes, Actinopterygii) from the Early Jurassic Posidonia shale of northern Germany". Fossil Record. 4 (1): 167–187. doi:10.5194/fr-4-167-2001. Retrieved 19 February 2022.
- Arratia (2003). "Leptolepis, Paraleptolepis (Teleostei) and a new fish name". Mitteilungen aus dem Museum für Naturkunde in Berlin. 6 (1): 157–159. Retrieved 19 February 2022.
- Hauff, B. (1953). "Ohmdenia multidentata nov. gen. et nov. sp. Ein neuer grober Fischfund aus den Posidonienschiefern des Lias e von Ohmden/Holzmaden in Württemburg". Neues Jahrb. Geol. P.-A. 97 (1): 39–50.
- Wretman, L.; Blom, H.; Kear, B. P. (2016). "Resolution of the Early Jurassic actinopterygian fish Pachycormus and a dispersal hypothesis for Pachycormiformes". Journal of Vertebrate Paleontology. 36 (5): 16–34. Bibcode:2016JVPal..36E6022W. doi:10.1080/02724634.2016.1206022. S2CID 89338085. Retrieved 19 February 2022.
- Hennig, E. (1918). "Uber Ptycholepis bollensis". Jahreshefte Verein vaterländischer Naturkunde. 74 (1): 173.
- Wenz, S. (1959). "Étude de Ptycholepis bollensis, poisson du Lias supérieur de l'Yonne et du Wurtemberg". Bulletin de la Société Géologique de France. 7 (9): 916–928. doi:10.2113/gssgfbull.S7-I.9.916. Retrieved 19 February 2022.
- Agassiz, L. (1843). Recherches sur les poissons fossiles.. (Vol. 2). Paris: Petitpierre.
- Maxwell, E. E.; Stumpf, S. (2017). "Revision of Saurorhynchus (Actinopterygii: Saurichthyidae) from the Early Jurassic of England and Germany". European Journal of Taxonomy (321): 1–29. doi:10.5852/ejt.2017.321. Retrieved 24 October 2021.
- Woodward, A. S. (1916). "I.—On a New Specimen of the Liassic Pachycormid Fish Saurostomus esocinus, Agassiz". Geological Magazine. 3 (2): 49–51. Bibcode:1916GeoM....3...49W. doi:10.1017/S0016756800191113. S2CID 128701357. Retrieved 19 February 2022.
- Hennig, E. (1925). "Chondrosteus Hindenburgi Pomp.---Ein "Stör" des württembergischen Ölschiefers (Lias epsilon)". Palaeontographica. 89 (2): 115–134.
- Thies, D. (1989). "Der Hirnschädel und das Gehirn von Tetragonolepis semicincta Bronn 1830 (Actinopterygii, Semionotiformes)". Palaeontographica Abteilung A. 45 (2): 1–32.
- Hennig, E. (1951). "Trachymetopon liassicum, Ald., ein Reisen-Crossopterygier aus Schwäbischem Ober-Lias". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 94 (1): 67–79.
- Dutel, Hugo; Herbin, Marc; Clément, Gaël (2015). "First occurrence of a mawsoniid coelacanth in the Early Jurassic of Europe". Journal of Vertebrate Paleontology. 35 (3): e929581. Bibcode:2015JVPal..35E9581D. doi:10.1080/02724634.2014.929581. S2CID 140557693. Retrieved 10 February 2022.
- McGowan, C. (1990). "Computed tomography confirms that Eurhinosaurus (Reptilia: Ichthyosauria) does have a tailbend". Canadian Journal of Earth Sciences. 27 (11): 1541–1545. Bibcode:1990CaJES..27.1541M. doi:10.1139/e90-164. Retrieved 10 February 2022.
- Maisch, M. W. (2022). "Ein neuer Eurhinosaurus (Reptilia: Ichthyosauria) aus der Posidonienschiefer-Formation (Unteres Toarcium) von Südwest-Deutschland mit Bemerkungen zur Nomenklatur und Taxonomie der Gattung". Jahreshefte der Gesellschaft für Naturkunde in Württemberg. 178: 117–148. Retrieved 2 May 2023.
- Maisch, Michael W. (2008). "Revision der Gattung Stenopterygius Jaekel, 1904 emend. von Huene, 1922 (Reptilia: Ichthyosauria) aus dem unteren Jura Westeuropas". Palaeodiversity. 1 (1): 227–271. Retrieved 10 February 2022.
- Maxwell, Erin E.; Cortés, Dirley (2020). "A revision of the Early Jurassic ichthyosaur Hauffiopteryx (Reptilia: Ichthyosauria), and description of a new species from southwestern Germany". Palaeontologia Electronica. 23 (2): 1–43. JSTOR 937. Retrieved 10 February 2022.
- Büttcher, R. (1989). "Uber die Nahrung eines Leptopterygius (Ichthyosauria, Reptilia) aus dem süddeutschen Posidonienschiefer (Unterer Jura) mit Bemerkungen uber den Magen der Ichthyosaurier". Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie). 155 (1): 1–19. Retrieved 10 February 2022.
- W. Maisch, Michael; T. Matzke, Andreas (2022). "Magnipterygius huenei n. gen. n. sp., a new small stenopterygiid (Reptilia: Ichthyosauria) from the Posidonienschiefer Formation of SW Germany". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 303 (2): 169–201. doi:10.1127/njgpa/2022/1042. S2CID 246797718. Retrieved 10 February 2022.
- Maxwell, E. E. (2012). "New Metrics To Differentiate Species of Stenopterygius (Reptilia: Ichthyosauria) from the Lower Jurassic of Southwestern Germany". Journal of Paleontology. 86 (1): 105–115. doi:10.1666/11-038.1. JSTOR 41409134. S2CID 130298530.
- Lindgren, J; Sjövall, P; Thiel, V; Zheng, W; Ito, S; Wakamatsu, K; Hauff, R; Kear, BP; Engdahl, A; Alwmark, C; Eriksson, ME; Jarenmark, M; Sachs, S; Ahlberg, PE; Marone, F; Kuriyama, T; Gustafsson, O; Malmberg, P; Thomen, A; Rodriguez-Meizoso, I; Uvdal, P; Ojika, M; Schweitzer, MH (2018). "Soft-tissue evidence for homeothermy and crypsis in a Jurassic ichthyosaur". Nature. 564 (1): 359–365. Bibcode:2018Natur.564..359L. doi:10.1038/s41586-018-0775-x. PMID 30518862. S2CID 54458324. Retrieved 10 February 2022.
- Dick, D. G.; Schweigert, G.; Maxwell, E. E. (2016). "Trophic niche ontogeny and palaeoecology of early Toarcian Stenopterygius(Reptilia: Ichthyosauria)". Palaeontology. 59 (3): 423–431. Bibcode:2016Palgy..59..423D. doi:10.1111/pala.12232. S2CID 87906152. Retrieved 10 February 2022.
- Maisch, M. W. (1998). "A new ichthyosaur genus from the Posidonia Shale (Lower Toarcian, Jurassic) of Holzmaden, SW-Germany with comments on the phylogeny of post-Triassic ichthyosaurs". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 209 (2): 47–48. doi:10.1127/njgpa/209/1998/47. Retrieved 10 February 2022.
- Maxwell, E.E. (2018). "Redescription of the "lost" holotype of Suevoleviathan integer (Bronn, 1844) (Reptilia: Ichthyosauria)". Journal of Vertebrate Paleontology. 38 (2): 23–36. Bibcode:2018JVPal..38E9833M. doi:10.1080/02724634.2018.1439833. S2CID 91013635. Retrieved 10 February 2022.
- Theodori, C. V. (1843). "Über einen kolossalen Ichthyosaurus trigonodon". Gelehrte Anzeigen der Königlich Bayerischen Akademie der Wissenschaften. 16 (2): 906–911.
- Laboury, Antoine; Bennion, Rebecca F; Thuy, Ben; Weis, Robert; Fischer, Valentin (2022). "Anatomy and phylogenetic relationships of Temnodontosaurus zetlandicus (Reptilia: Ichthyosauria)". Zoological Journal of the Linnean Society. 118 (1): 26–38. doi:10.1093/zoolinnean/zlab118. Retrieved 2 March 2022.
- Großmann (2006). "Taxonomy, Phylogeny and Palaeoecology of the Plesiosauroids (sauropterygia, Reptilia) from the Posidonia Shale (Toarcian, Lower Jurassic) of Holzmaden, South West Germany: Dissertation Zur Erlangung Des Grades Eines Doktors Der Naturwissenschaften". Geowissenschaftlichen Fakultät der Eberhard-Karls-Universität. 1 (1): 1–135. Retrieved 10 February 2022.
- Vincent, P. (2011). "A re-examination of Hauffiosaurus zanoni, a pliosauroid from the Toarcian (Early Jurassic) of Germany". Journal of Vertebrate Paleontology. 31 (1): 340–351. Bibcode:2011JVPal..31..340V. doi:10.1080/02724634.2011.550352. S2CID 84743241. Retrieved 10 February 2022.
- Maisch, M. W.; RŘcklin, M. (2000). "Cranial osteology of the sauropterygian Plesiosaurus brachypterygius from the Lower Toarcian of Germany". Palaeontology. 43 (1): 29–40. Bibcode:2000Palgy..43...29M. doi:10.1111/1475-4983.00117. S2CID 84916676. Retrieved 10 February 2022.
- Vincent, P.; Allemand, R.; Taylor, P. D.; Suan, G.; Maxwell, E. E. (2017). "New insights on the systematics, palaeoecology and palaeobiology of a plesiosaurian with soft tissue preservation from the Toarcian of Holzmaden, Germany". The Science of Nature. 104 (6): 51. Bibcode:2017SciNa.104...51V. doi:10.1007/s00114-017-1472-6. hdl:10141/622228. PMID 28578532. S2CID 24957821. Retrieved 10 February 2022.
- Adam S. Smith; Peggy Vincent (2010). "A new genus of pliosaur (Reptilia: Sauropterygia) from the Lower Jurassic of Holzmaden, Germany". Palaeontology. 53 (5): 1049–1063. Bibcode:2010Palgy..53.1049S. doi:10.1111/j.1475-4983.2010.00975.x.
- Huene, F. v. (1923). "Ein neuer Plesiosaurier aus dem oberen Lias Württembergs" (PDF). Jahreshefte des Vereins für Vaterländische Naturkunde in Württemberg. 79 (1): 3–23. Retrieved 10 February 2022.
- Vincent, P.; Weis, R.; Kronz, G.; Delsate, D. (2019). "Microcleidus melusinae, a new plesiosaurian (Reptilia, Plesiosauria) from the Toarcian of Luxembourg". Geological Magazine. 156 (1): 99–116. Bibcode:2019GeoM..156...99V. doi:10.1017/S0016756817000814. S2CID 135111068. Retrieved 10 February 2022.
- O'keefe, F. R. (2004). "Preliminary description and phylogenetic position of a new plesiosaur (Reptilia: Sauropterygia) from the Toarcian of Holzmaden, Germany" (PDF). Journal of Paleontology. 78 (5): 973–988. doi:10.1666/0022-3360(2004)078<0973:PDAPPO>2.0.CO;2. JSTOR 4094922. S2CID 53590349.
- Sachs, Sven; Abel, Pascal; Madzia, Daniel (2023). "Unusual plesiosaur vertebrae from the Lower Jurassic Posidonia Shale of Germany". Historical Biology. Retrieved 17 August 2023.
- Carroll, R. L. (1985). "A pleurosaur from the Lower Jurassic and the taxonomic position of the Sphenodontida". Palaeontographica Abteilung A. 189 (1): 1–28. Retrieved 10 February 2022.
- Klein, N.; Scheyer, T.M. (2017). "Microanatomy and life history in Palaeopleurosaurus (Rhynchocephalia: Pleurosauridae) from the Early Jurassic of Germany". The Science of Nature. 104 (4): 4. Bibcode:2017SciNa.104....4K. doi:10.1007/s00114-016-1427-3. PMID 28005148. S2CID 27133670. Retrieved 10 February 2022.
- Kuhn (1961). "Eine Schildkröte aus dem Lias epsilon Von Süddeutschland". Jh. Ver. Vaterländ. Naturk. Württemb. 116 (1): 285–287.
- Münster, G.G. (1834). "Mittheilung, an Professor Bronn gerichtet". Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde. 16 (1): 42–43.
- Theodori, L. (1831). "Über die Knochen vom Genus Pterodactylus aus der Lias-Formation in der Gegend von Banz". Isis von Oken. 21 (1): 276–281.
- Joyce, W. G. (2017). "A review of the fossil record of basal Mesozoic turtles". Bulletin of the Peabody Museum of Natural History. 58 (1): 65–113. doi:10.3374/014.058.0105. S2CID 54982901. Retrieved 10 February 2022.
- Joger, U.; Kosma, R.; Zellmer, H.; Röhling, H. G. (2018). "Saurier im Braunschweiger Land. Die Fund-und Grabungsstellen von Hondelage und Schandelah (Unterjura, Posidonienschiefer) sowie des Langenberg bei Goslar/Oker (Oberjura, Malm)(Exkursion N am 7. April 2018)". Jahresberichte und Mitteilungen des Oberrheinischen Geologischen Vereins. 4 (1): 447–479. doi:10.1127/jmogv/100/0013. Retrieved 10 February 2022.
- Johnson, Michela M.; Young, Mark T.; Brusatte, Stephen L. (2020). "The phylogenetics of Teleosauroidea (Crocodylomorpha, Thalattosuchia) and implications for their ecology and evolution". PeerJ. 8: e9808. doi:10.7717/peerj.9808. PMC 7548081. PMID 33083104.
- Michela, M Johnson; Young, Mark T; Brusatte, Stephen L (2020). "Emptying the wastebasket: a historical and taxonomic revision of the Jurassic crocodylomorph Steneosaurus" (PDF). Zoological Journal of the Linnean Society. 189 (428–448). Retrieved 20 February 2022.
- Heller, F. (1953). "Ein Mystriosaurus-Fund im Lias epsilon von Mistelgau". Geologische Blätter für Nordost-Bayern und angrenzende Gebiete. 6 (2): 146–148.
- Sachs, S; Johnson, M.M.; Young, M.T.; Abel, P. (2019). "The mystery of Mystriosaurus: Redescribing the poorly known Early Jurassic teleosauroid thalattosuchians Mystriosaurus laurillardi and Steneosaurus brevior". Acta Palaeontologica Polonica. 64 (3): 565–579. doi:10.4202/app.00557.2018. S2CID 202892925. Retrieved 10 February 2022.
- Pierce, S. E.; Williams, M.; Benson, R. B. (2017). "Virtual reconstruction of the endocranial anatomy of the early Jurassic marine crocodylomorph Pelagosaurus typus (Thalattosuchia)". PeerJ. 3225 (1): 1–32. Retrieved 10 February 2022.
- Wellnhofer, Peter (1974). Campylognathoides Liasicus (Quenstedt), an Upper Liassic Pterosaur from Holzmaden. USA: Carnegie Museum of Natural History.
- Plieninger, F. (1894). "Campylognathus Zittelli. Ein neuer Flugsaurier aus dem Oberen Lias Schwabens". Palaeontographica. 41 (1): 193–222.
- Padian, K. (2008). "The Early Jurassic pterosaur Dorygnathus banthensis (Theodori, 1830)". Special Papers in Palaeontology. 92: 69–07. Retrieved 10 February 2022.
- Hübner, M.; Gischler, E.; Kosma, R. (2020). "Rare pterosaur remains tentatively referred to Dorygnathus banthensis (Theodori, 1830) from the Lower Jurassic (Posidonia Shale) of Schandelah (Lower Saxony, Germany)" (PDF). Braunschweiger Naturkundliche Schriften. 16 (1): 59–82. Retrieved 10 February 2022.
- Wild, R. (1971). "Dorygnathus mistelgauensis n. sp., ein neuer Flugsaurier aus dem Lias Epsilon von Mistelgau (Fränkischer Jura)". Geologische Blätter für Nordost-Bayern und angrenzende Gebiete. 21 (4): 178–195.
- Frey, E.; Tischlinger, H.; Buchy, MC; Martill, DM (2003). "New specimens of Pterosauria (Reptilia) with soft parts with implications for pterosaurian anatomy and locomotion". Geological Society, London, Special Publications. 217 (1): 233–266. Bibcode:2003GSLSP.217..233F. doi:10.1144/GSL.SP.2003.217.01.14. S2CID 130462931. Retrieved 10 February 2022.
- Broili, F. (1939). "Ein Dorygnathus mit Hautresten" (PDF). Sitzungsberichte - Bayerische Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Klasse (4): 129–132. Retrieved 10 February 2022.
- O'Sullivan, M.; Martill, D.M. (2017). "The taxonomy and systematics of Parapsicephalus purdoni (Reptilia: Pterosauria) from the Lower Jurassic Whitby Mudstone Formation, Whitby, U.K". Historical Biology. 29 (8): 1009–1018. doi:10.1080/08912963.2017.1281919. S2CID 132532024. Retrieved 10 February 2022.
- Wild, R. (1978). "Ein Sauropoden-Rest (Reptilia, Saurischia) aus dem Posidonienschiefer (Lias, Toarcium) von Holzmaden". Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie). 41 (2): 1–15.
- Zürlick, F. (1956). Der Teufelsgraben bei Ödhof. Berlin: Aufschluss.
- Wilde, V. (2001). "Die Landpflanzen-Taphozönose aus dem Posidonienschiefer des Unteren Jura (Schwarzer Jura [Epsilon], Unter-Toarcium) in Deutschland und ihre Deutung". Staatliches Museum für Naturkunde. 304 (2): 1–12. Retrieved 3 March 2022.
- Vogellehner, D. (1982). "Zur Anatomie und Systematik von "Treibhölzern" aus dem Posidonienschiefer von Holzmaden (Schwäb. Alb)". Cour. Forsch.-Inst. Senckenberg. 56 (1): 15–22.
- Seilacher, A. (1990). "Die Holzmadener Posidonienschiefer. Entstehung der Fossillagerstätte und eines Erdölmuttergesteines". Klassische Fundstellen der Paläontologie. 2 (1): 107–131. Retrieved 3 March 2022.
- Ammon, V.L. (1875). Die Jura Ablagerungen zwischen Regensburg und Passau. München: Theodor Ackermann. Retrieved 3 March 2022.
- Heunisch, C. (2014). "Das schwarze Jurameer und seine unsichtbaren Bewohner". Jurameer. 2 (1): 7–10. Retrieved 3 March 2022.
- Galasso, F.; Feist-Burkhardt, S.; Schneebeli-Hermann, E. (2022). "The palynology of the Toarcian Oceanic Anoxic Event at Dormettingen, southwest Germany, with emphasis on changes in vegetational dynamics". Review of Palaeobotany and Palynology. 304 (1): 104701. Retrieved 3 October 2023.
- Houben, A. J.; Goldberg, T.; Slomp, C. P. (2021). "Biogeochemical evolution and organic carbon deposition on the Northwestern European Shelf during the Toarcian Ocean Anoxic Event". Palaeogeography, Palaeoclimatology, Palaeoecology. 565 (1): 897–912. Bibcode:2021PPP...565k0191H. doi:10.1016/j.palaeo.2020.110191. S2CID 233805324. Retrieved 3 March 2022.
- Hofmann, Christa-Ch.; Odgerel, Nyamsambuu; Seyfullah, Leyla J. (2021). "The occurrence of pollen of Sciadopityaceae Luerss. through time". Fossil Imprint. 77 (2): 271–281. doi:10.37520/fi.2021.019. S2CID 245555379. Retrieved 27 December 2021.
- Madler, K.A. (1956). "Pollen analytical studies on the Posidonian Shale[ Pollenanalytische untersuchungen im Posidonienschiefer ]". PalZ (Paläontologische Zeitschrift). 30 (1): 18. Retrieved 3 March 2022.
- Suan, G.; Nikitenko, B.L.; Rogov, M.A.; Baudin, F.; Spangenberg, J.E.; Knyazev, V.G.; Glinskikh, L.A.; Goryacheva, A.A.; Adatte, T.; Riding, J.B.; Föllmi, K.B.; Pittet, B.; Mattioli, E.; Lécuyer, C. (2011). "Polar record of Early Jurassic massive carbon injection". Earth and Planetary Science Letters. 312 (1): 102–113. Bibcode:2011E&PSL.312..102S. doi:10.1016/j.epsl.2011.09.050. Retrieved 3 March 2022.
- Thiergart, F. (1944). "The plant remains of the Posidonia Shale [Die Pflanzenreste des Posidonienschiefers]". Archiv für Lagerstättenforschung, zur Paläogeographie und Bitumen-Fährung des Posidonihiefers im Deutschen Lias. 77 (1): 45–48.
- Wade-Murphy, J.; Kuerschner, W. M (2006). "A new technique to infer the botanical affinity of palynomorphs, and its application on Spheripollenites psilatus from the Toarcian of Bornholm, Denmark" (PDF). In 7 Th European Palaeobotany Palynology Conference (1–2): 153–154. Archived from the original (PDF) on 22 December 2021. Retrieved 13 October 2021.
- Salfeld, H. (1907). "Fossile Land-Pflanzen der Rät- und Juraformation Südwestdeutschlands" (PDF). Palaeontographica. 54 (4): 163–204. Retrieved 3 March 2022.
- Salfeld, H. (1909). "Beiträge zur Kenntnis jurassischer Pflanzenreste aus Norddeutschland". Palaeontographica. 56: 1–35. Retrieved 3 March 2022.
- Maubeuge, P. (1947). "Sur l'existence du genre Neocalamites dans le Toarcien du Grand-Duché de Luxembourg". Archives de l'Institut Gratul-Ducal de Luxembourg, Section des Sciences naturelles, physiques et mathématiques. Nouvelle Série. 17 (2): 59–64. Retrieved 3 March 2022.
- Böttcher, R. (1998). "Leben und Tod im Meer des Posidonienschiefers". PalZ (Paläontologische Zeitschrift). 25 (2): 83–96. Retrieved 3 March 2022.
- Kurr, J. G. (1845). "Beiträge zur fossilen Flora der Juraformation Württembergs". Schule zu Stuttgart den. 27 (6): 1–17. Retrieved 3 March 2022.
- Küpper, K. (1968). "Die Gattung Otozamites". Taxon. 17 (5): 548–552. doi:10.2307/1216063. JSTOR 1216063.
- Zijlstra, G.; van Konijnenburg-van Cittert, J. H. (2019). "Proposal to conserve the name Otozamites (fossil Cycadophyta: Bennettitales) against Otopteris". Taxon. 68 (4): 874–875. doi:10.1002/tax.12112. hdl:1874/389836. S2CID 213356496. Retrieved 3 March 2022.
- Keller, T.; Wilde, V. (2000). "Ein Koniferenrest aus dem Posidonienschiefer des Unteren Jura (Schwarzer Jura [epsilon], Unter-Toarcium) von Süddeutschland". Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie). 282 (2): 1–17. Retrieved 3 March 2022.
- Tollmann, A. (1976). "Analyse des klassischen nordalpinen Mesozoikums". Stratigraphie, Fauna und Fazies der Nördlichen Kalkalpen. 2 (1): 123–141. OCLC 3710249. Retrieved 3 March 2022.
- Süss, H.; Philippe, M. (1993). "Holzanatomische Untersuchungen an einem fossilen Holz, Circoporoxylon grandiporosum Müller-Stoll et Schultze-Motel, aus dem Unteren Jura von Frankreich". Feddes Repertorium. 104 (8): 451–463. doi:10.1002/fedr.19931040706. Retrieved 3 March 2022.
- Philippe, M.; Tchoumatchenco, P. (2008). "Palaeoecologically significant wood genus Xenoxylon discovered in the East Stara Planina Mts.(East Bulgaria) Balaban Formation (Toarcian, Early Jurassic)". Comptes rendus de l'Académie bulgare des Sciences. 61 (5): 633–638. Retrieved 3 March 2022.
- Schultze-Motel, J. (1960). "Anatomische Untersuchungen an mesozoischen Gymnospermen-Hölzern". Dissertation Pädagogische Hochschule Potsdam. 156 (1): 1–25.
- Mueller-Stoll, W. R. (1986). "Evolutionary trends in gymnospermous wood structures during Mesozoic-Protopinaceous woods in the German Jurassic" (PDF). Palaeobotanist. 35 (3): 233–235. Retrieved 3 March 2022.
- Bamford, M. K.; Philippe, M.; Thévenard, F. (2016). "Long overdue extinction of the Protopinaceae" (PDF). Review of Palaeobotany and Palynology. 234 (1): 25–30. Bibcode:2016RPaPa.234...25B. doi:10.1016/j.revpalbo.2016.06.006. Retrieved 3 March 2022.
- Müller-Stoll, W.R.; Schultze-Motel, J. (1990). "Gymnospermen-Hölzer des Deutschen Jura. Teil 3: Abietoid (modern) getüpfelte Hölzer". Zeitschrift der deutschen geologischen Gesellschaft. 141 (2): 61–77. doi:10.1127/zdgg/141/1990/61.
- Philippe, M.; Thevenard, F. (1996). "Distribution and palaeoecology of the Mesozoic wood genus Xenoxylon: palaeoclimatological implications for the Jurassic of Western Europe". Review of Palaeobotany and Palynology. 91 (4): 353–370. Bibcode:1996RPaPa..91..353P. doi:10.1016/0034-6667(95)00067-4. Retrieved 3 March 2022.