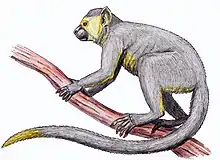
Pierolapithecus
Pierolapithecus catalaunicus is an extinct species of primate which lived around 12.5-13 million years ago during the Miocene in what is now Hostalets de Pierola, Catalonia, Spain. Some researchers believe that it is a candidate for common ancestor to the great ape clade, or is at least closer than any previous fossil discovery.[1] Others suggest it being a pongine,[2] or a dryopith.[3] On 16 October 2023, scientists reported the facial reconstruction of the great ape.[4][5]
| Pierolapithecus Temporal range: Miocene, | |
|---|---|
.jpg.webp) | |
| Pierolapithecus catalaunicus | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Primates |
| Suborder: | Haplorhini |
| Infraorder: | Simiiformes |
| Family: | Hominidae |
| Subfamily: | incertae sedis |
| Genus: | †Pierolapithecus Moyà-Solà et al., 2004 |
| Species: | †P. catalaunicus |
| Binomial name | |
| †Pierolapithecus catalaunicus | |
History
The splanchnocranium was discovered in 2002 and systematic excavations took place during May and June 2003.[6] The species was described by a team of Spanish paleoanthropologists led by Salvador Moyà-Solà on the basis of a fossil skeleton, IPS21350 (nicknamed Pau ("peace" in Catalan as it was announced alongside Spanish demonstrations against the Iraq War)[7]), discovered in December 2002. The finding was first reported in the journal Science on November 19, 2004. The skeleton is of an adult male individual, composed of 83 bones that make up the splanchnocranium, both maxillae, a complete set of cheek teeth, both canines, a right central incisor, zygomatics, lacrimals, a partial frontal, carpals, metacarpals, manual phalanges from two hands, tarsals, metatarsals, pedal phalanges, right patellar distal epiphysis, a left radius, some long bone diaphyses, two pelvic pieces, three vertebrae, two intact ribs, and twelve rib fragments of large size. They named their new genus after the nearby village Els Hostalets de Pierola, and Catalonia respectively.[8]
Description
Moyà-Solà et al. initially founded the species on a set of unique characteristics, of which are the following. The frontal processes of the face remain on the same plane, the nasals are flat and sit beneath the lower rims of the orbit, the glabella is posteriorly oriented, the face is low, the brows are thin, the zygomatic root is high, and the nasoalveolar clivus is high. The rear border of the incisive foramen is in line with the P3, the palate is deep and stout, the nasal aperture is widest at the base, the interorbital distance is wide, the zygomatics expand to the side, the P3 is similar in size to the P4, there is reduced cusp heteromorphy, all molars save from the M3 are elongated, the upper molars and premolars lack cingula, the lingual cusps of the upper molars are positioned peripherally, the M2 is large and has cusp heteromorphy, and the upper canine is large and compressed in the crown. The ribs are very curved to form a thorax that is anteroposteriorly compressed, the clavicle is robust, lacking a ventral keel on the mid-lumbars, the pedicles of the neural arch are robust and stout, the spinous processes are slightly caudally inclined, the pedicle-body inserts the transverse processes, dorsally oriented and pedicle-born transverse process, the metacarpals and phalanges are short, the os centrale are unfused, the triquetrum is small and non-articulating with the ulnar styloid, and the crevice inserting meniscus attachment and pisiform facet is distally shifted.[8]
The holotypic individual is estimated to have weighed 30 kg (66.13 lbs).[7]
Locomotion
Overall, the adduction and supination capacity of the wrist, specially built thorax, scapular shift to the back (which was inferred through the long, chimpanzee-like clavicles), and stiff lumbar vertebrae suggest that positional behavior and orthograde locomotion were emphasized. This type of movement is diagnostic for all extant apes including humans, but it is rarely documented in the fossil record. Other hominids that have this suite are Oreopithecus and, although less skeletally complete, Dryopithecus. Earlier taxa—Proconsul, Afropithecus, Equatorius, Nacholapithecus—retain basal characters and the similarly-aged Morotopithecus practiced orthograde locomotion but was probably sister to apes (based on facial structure). The shortened phalanges suggest ancestral palmigrade adaptations, but it is unlikely Pierolapithecus practiced much or any suspensory behavior. However, vertical climbing and suspension are independent abilities that are integral to ape evolution. Below-branch suspension may have evolved repeatedly or in convergence later and independently in the ape lineage.[8]

Further analysis by Almécija et al. (2009) suggests that very long and curved phalanges is decoupled with orthograde features related to vertical climbing being acquired. They find that the condition in this genus is related to a retained pronograde plan. Although the lumbars, ribs, and carpals are orthograde, the degree of this in the phalanges is only slight. They agree, however, that many traits were independently acquired, leading to new advances being superimposed and basal characters retained for an extended time. Pierolapithecus lacked adaptations for suspensory hanging, but unlike Moyà-Solà et al. report, they suggest that it was capable of doing so, only that it was not adaptively relevant.[9] Although, the latter remains disputed.[10]
The patella was like extant apes in dimensions, which is typically regarded as having a mobile knee. Pierolapithecus differs from monkeys, hylobatids, and basal hominoids through thicker patellae. As such, a derived knee might be related to enhanced climbing, notably vertical climbing.[11] The pelvis shares an ancestral template with Proconsul nyanzae, which was modified for orthograde behavior (assuming that hypothesis is accepted), and suggests homoplasy in ape pelves.[12]
Classification
Pierolapithecus demonstrates derived facial features and apelike skeletal adaptations that suggest that it is an early member of the ape clade. This genus, 12.5-13 mya in age, postdates the hylobatid-hominid split, which occurred anywhere from 14.9±2 or 14.6±2.6 mya. Much of the skeleton is derived, but the shortened phalanges are indicative of palmigrade adaptations that are primitive. Moyà-Solà et al. note that this mosaic is important in ape evolution. The large amount of homoplasy in ape locomotion creates considerable taxonomic confusion. The late Middle Miocene is the farthest trace of a Pierolapithecus-like character group, and assuming that this identifies the earliest apes, is the farthest trace of hominids. As well, early hominids are substantially more primitive than estimated, which may explain why no early great apes were previously reported.[8] These early traits would have been maintained, overlaid, and modified to suite new adaptations that occurred independently.[9]
Begun (2009) suggested that Pierolapithecus and Anoiapithecus are synonymous with Dryopithecus, but this was not followed by Alba and Moyà-Solà (2010) who reasoned that both have enough craniofacial difference to justify separation. They do not regard this species as a dryopith due to a lack of diagnostic features. Instead, they support placement as a basal hominid inheriting thick enamel from the afropithecids, which may be ancestral to apes.[13] Other hypotheses are that the taxon represents a stem pongine.[2] Rather than a full common ancestor, it has been suggested that the species may be ancestral to humans, chimpanzees and gorillas but not orangutans, given certain facial characteristics.[14] Sevim-Erol et al. (2023) suggest that this genus is distinguished from pongines and share traits with extant hominines, suggesting a sister relationship with Dryopithecus (a tribe they call Dryopithecini, having thick and thin enamel much like Ardipithecus/australopiths and Pan/hominins).[3]
Paleoecology
Pierolapithecus bore thick enamel found in hard-object feeders, but its diet is not yet known aside from possibly having fed in trees like orangutans.[7][13] It was discovered at the locality of BCV1, which formed when the northwestern Mediterranean rifted to form a stretch between two mountain ranges. The proximal to distal-marginal alluvial-fan sediments cover the Miocene. It was discovered as a fossiliferous area by Guerín in the 1920s with an ape M2 mistaken as a suid, followed by the discovery of Dryopithecus fontani, Hispanopithecus laietanus, and 'Sivapithecus' occidentalis in the area. From the area hailing Pierolapithecus specifically was explored from the 1950s-1970s from a garbage dump. 19 large and small mammals were discovered at the site, almost 300 macroinvertebrate fossils, and 83 hominid fossils (composing the holotype skeleton).[6]
The fauna comprises the Pierolapithecus, megaherbivores like elephants, and various others like carnivores, artiodactyls, turtles, and small-medium fragments. Pierolapithecus bears evidence of scavenging whereas the other fossils show signs of being scattered across an alluvial plain. The micromammals show signs of digestion by predators, probably by barn owls and others. The environment was quite humid, warm, and forested. The fauna is most alike France and central Europe in composition. More insectivores, arboreal dormice, and flying squirrels support a humid environment, and the open woodlands of other sites would have made hominid occupation impossible.[6]
That Pierolapithecus would be ancestral to modern great apes is debated largely because this great ape was found in the Iberian Peninsula, while most of the fossil evidence of the evolution of hominids and hominins has been located in East Africa and Southeast Asia. Because the Mediterranean Sea contracted several times in the past, migration of terrestrial fauna between Africa and Europe was permitted.[15]
See also
- Anoiapithecus – Extinct genus of ape from the Miocene
- Chororapithecus – Extinct hominine genus from the Miocene
- Griphopithecus – Extinct genus of primates
- Nakalipithecus – Extinct species of ape
- Samburupithecus – Extinct genus of primate from Miocene Kenya
References
- Rincon, Paul (18 November 2004). "'Original' great ape discovered". BBC. Retrieved 23 January 2013.
- Pérez de los Ríos, Miriam; Moyà-Solà, Salvador; Alba, David M. (2012-09-01). "The nasal and paranasal architecture of the Middle Miocene ape Pierolapithecus catalaunicus (primates: Hominidae): Phylogenetic implications". Journal of Human Evolution. 63 (3): 497–506. doi:10.1016/j.jhevol.2012.05.012. ISSN 0047-2484. PMID 22819226.
- Sevim-Erol, Ayla; Begun, D. R.; Sözer, Ç Sönmez; Mayda, S.; van den Hoek Ostende, L. W.; Martin, R. M. G.; Alçiçek, M. Cihat (2023-08-23). "A new ape from Türkiye and the radiation of late Miocene hominines". Communications Biology. 6 (1): 842. doi:10.1038/s42003-023-05210-5. ISSN 2399-3642. PMC 10447513. PMID 37612372.
- Johnson, Mark (24 October 2023). "Scientists reconstructed the face of a 12 million-year-old great ape - Fossils of the extinct species Pierolapithecus catalaunicus may reveal clues about our origins". The Washington Post. Archived from the original on 24 October 2023. Retrieved 25 October 2023.
- Pugh, Kelsey D.; et al. (16 October 2023). "The reconstructed cranium of Pierolapithecus and the evolution of the great ape face". PNAS. 120 (44): e2218778120. doi:10.1073/pnas.2218778120. PMID 37844214. Archived from the original on 24 October 2023. Retrieved 25 October 2023.
- Casanovasvilar, I; Alba, D; Moyasola, S; Galindo, J; Cabrera, L; Garces, M; Furio, M; Robles, J; Kohler, M; Angelone, C (2008). "Biochronological, taphonomical, and paleoenvironmental background of the fossil great ape Pierolapithecus catalaunicus (Primates, Hominidae)". Journal of Human Evolution. 55 (4): 589–603. doi:10.1016/j.jhevol.2008.05.004. PMID 18691737.
- Bezanson, Michele; MacKinnon, Katherine C; Riley, Erin; Campbell, Christina J; Nekaris, K.A.I (Anna); Estrada, Alejandro; Di Fiore, Anthony F; Ross, Stephen; Jones‐Engel, Lisa E, eds. (2016-06-14). The International Encyclopedia of Primatology (1 ed.). Wiley. doi:10.1002/9781119179313.wbprim0216. ISBN 978-0-470-67337-9.
- Moya-Sola, S.; Köhler, M.; Alba, D. M.; Casanovas-Vilar, I.; Galindo, J. (2004). "Pierolapithecus catalaunicus, a New Middle Miocene Great Ape from Spain" (PDF). Science. 306 (5700): 1339–1344. Bibcode:2004Sci...306.1339M. doi:10.1126/science.1103094. PMID 15550663. S2CID 129576842.
- Almécija, Sergio; Alba, David M.; Moyà-Solà, Salvador (2009). "Pierolapithecus and the functional morphology of Miocene ape hand phalanges: paleobiological and evolutionary implications". Journal of Human Evolution. 57 (3): 284–297. doi:10.1016/j.jhevol.2009.02.008. PMID 19631964.
- Alba, David M.; Almécija, Sergio; Moyà-Solà, Salvador (2010-07-01). "Locomotor inferences in Pierolapithecus and Hispanopithecus: Reply to Deane and Begun (2008)". Journal of Human Evolution. 59 (1): 143–149. doi:10.1016/j.jhevol.2010.02.002. ISSN 0047-2484. PMID 20510436.
- Pina, Marta; Almécija, Sergio; Alba, David M.; O'Neill, Matthew C.; Moyà-Solà, Salvador (2014). "The Middle Miocene Ape Pierolapithecus catalaunicus Exhibits Extant Great Ape-Like Morphometric Affinities on Its Patella: Inferences on Knee Function and Evolution". PLOS ONE. 9 (3): e91944. Bibcode:2014PLoSO...991944P. doi:10.1371/journal.pone.0091944. ISSN 1932-6203. PMC 3956854. PMID 24637777.
- Hammond, Ashley S.; Alba, David M.; Almécija, Sergio; Moyà-Solà, Salvador (2013). "Middle Miocene Pierolapithecus provides a first glimpse into early hominid pelvic morphology". Journal of Human Evolution. 64 (6): 658–666. doi:10.1016/j.jhevol.2013.03.002. PMID 23545221.
- Alba, D. M.; Fortuny, J.; Moyà-Solà, S. (2010). "Enamel thickness in the Middle Miocene great apes Anoiapithecus , Pierolapithecus and Dryopithecus". Proceedings of the Royal Society B: Biological Sciences. 277 (1691): 2237–2245. doi:10.1098/rspb.2010.0218. ISSN 0962-8452. PMC 2880156. PMID 20335211.
- Begun, D. R. (2015). Fossil record of Miocene hominoids (2nd ed.). Berlin: Springer. pp. 1304–1306. ISBN 978-3-642-39980-0.
- Roegl, Fred (1999). "Mediterranean and Paratethys. Facts and hypotheses of an Oligocene to Miocene paleogeography (short overview)". Geol. Carpathica. 50: 339–349.
External links
| |||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||