Alpha cell
Alpha cells (α cells) are endocrine cells that are found in the Islets of Langerhans in the pancreas. Alpha cells secrete the peptide hormone glucagon in order to increase glucose levels in the blood stream.[1]
| Alpha cell | |
|---|---|
 Pancreatic islets (islets of Langerhans). | |
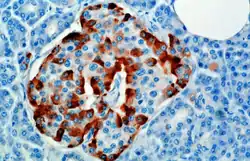 Alpha cells in red | |
| Details | |
| System | Endocrine |
| Location | Pancreatic islet |
| Function | Glucagon secretion |
| Identifiers | |
| TH | H3.04.02.0.00025 |
| FMA | 70585 |
| Anatomical terms of microanatomy | |
Discovery
Islets of Langerhans were first discussed by Paul Langerhans in his medical thesis in 1869.[2] This same year, Laguesse named them after Langerhans.[3] At first, there was a lot of controversy about what the Islets were made of and what they did.[3] It appeared that all of the cells were the same within the Islet, but were histologically distinct from acini cells.[3] Laguesse discovered that the cells within the Islets of Langerhans contained granules that distinguished them from acini cells.[3] He also determined that these granules were products of the metabolism of the cells in which they were contained.[3] Michael Lane was the one to discover that alpha cells were histologically different than beta cells in 1907.[3]
Before the function of alpha cells was discovered, the function of their metabolic product, glucagon, was discovered. The discovery of the function of glucagon coincides with the discovery of the function of insulin. In 1921, Banting and Best were testing pancreatic extracts in dogs that had had their pancreas removed. They discovered that "insulin-induced hypoglycemia was preceded by a transient, rather mild hyperglycemia..."[4] Murlin is credited with the discovery of glucagon because in 1923 they suggested that the early hyperglycemic effect observed by Banting and Best was due to "a contaminant with glucogenic properties that they also proposed to call 'glucagon,' or the mobilizer of glucose".[4] In 1948, Sutherland and de Duve established that alpha cells in the pancreas were the source of glucagon.[4]
Anatomy
Alpha cells are endocrine cells, meaning they secrete a hormone, in this case glucagon. Alpha cells store this glucagon in secretory vesicles that typically have an electron dense core and a grayish outer edge.[1] It is believed that alpha cells make up approximately 20% of endocrine cells within the pancreas.[1] Alpha cells are most commonly found on the dorsal side of the pancreas and are very rarely found on the ventral side of the pancreas.[1] Alpha cells are typically found in compact Islets of Langerhans, which are themselves typically found in the body of the pancreas.[1]
Function
Alpha cells function in the maintenance of blood glucose levels. Alpha cells are stimulated to produce glucagon in response to hypoglycemia, epinephrine, amino acids, other hormones, and neurotransmitters.[5]
Glucagon Secretion and Control of Gluconeogenesis
Glucagon functions to signal the liver to begin gluconeogenesis which increases glucose levels in the blood.[5] Glucagon will bind to the glucagon receptors on the plasma membranes of hepatocytes (liver cells). This ligand binding causes the activation of adenylate cyclase, which causes the creation of cyclic AMP (cAMP).[6] As the intracellular concentration of cAMP rises, protein kinase A (PKA) is activated and phosphorylates the transcription factor cAMP Response Element Binding (CREB) protein.[6] CREB then induces transcription of glucose-6-phosphatase and phosphoenolpyruvate carboxylase (PEPCK). These enzymes increase gluconeogenic activity.[6] PKA also phosphorylates phospho-fructokinase 2 (PFK2)/fructose 2,6-biphsophatase (FBPase2), inhibiting PFK2 and activating FBPase2.[6] This inhibition decreases intracellular levels of fructose 2,6-biphosphate and increases intracellular levels of fructose 6-phosphate which decreases glycolytic activity and increases gluconeogenic activity.[6] PKA also phosphorylates pyruvate kinase which causes an increase in intracellular levels of fructose 1,6-biphosphate and decreases intracellular levels of pyruvate, further decreasing glycolytic activity.[6] The most important action of PKA in regulating gluconeogenesis is the phosphorylation of phosphorylase kinase which acts to initiate the glycogenolysis reaction, which is the conversion of glycogen to glucose, by converting glycogen to glucose 1-phosphate.[6]
Alpha cells also generate Glucagon-like peptide-1 and may have protective and regenerative effect on beta cells. They possibly can transdifferentiate into beta cells to replace lost beta cells.[7]
Regulation of glucagon secretion
There are several methods of control of the secretion of glucagon. The most well studied is through the action of extra-pancreatic glucose sensors, including neurons found in the brain and spinal cord, which exert control over the alpha cells in the pancreas.[5] Indirect, non-neuronal control has also been found to influence secretion of glucagon.[5]
Neuronal Control
The most well studied is through the action of extra-pancreatic glucose sensors, including neurons found in the brain, which exert control over the alpha cells in the pancreas.[5] The pancreas is controlled by both the sympathetic nervous system and the parasympathetic nervous system, although the method these two systems use to control the pancreas appears to be different.[8]
Sympathetic control of the pancreas appears to originate from the sympathetic preganglionic fibers in the lower thoracic and lumbar spinal cord.[9] According to Travagli et al. "axons from these neurons exit the spinal cord through the ventral roots and supply either the paravertebral ganglia of the sympathetic chain via communicating rami of the thoracic and lumbar nerves, or the celiac and mesenteric ganglia via the splanchnic nerves. The catecholaminergic neurons of these ganglia innervate the intrapancreatic ganglia, islets and blood vessels..."[9] The exact nature of the effect of sympathetic activation on the pancreas has been difficult to discern. However, a few things are known. It appears that stimulation of the splanchnic nerve lowers plasma insulin levels possibly through the action of α2 adrenoreceptors on beta cells.[9] It has also been shown that stimulation of the splanchnic nerve increases glucagon secretion.[9] Both of these findings together suggest that sympathetic stimulation of the pancreas is meant to maintain blood glucose levels during heightened arousal.[9]
Parasympathetic control of the pancreas appears to originate from the Vagus nerve.[8] Electrical and pharmacological stimulation of the Vagus nerve increases secretion of glucagon and insulin in most mammalian species, including humans. This suggests that the role of parasympathetic control is to maintain normal blood glucose concentration under normal conditions.[8]
Non-neuronal Control
Non-neuronal control has been found to be indirect paracrine regulation through ions, hormones, and neurotransmitters. Zinc, insulin, serotonin, γ-aminobutyric acid, and γ-hydroxybutyrate, all of which are released by beta cells in the pancreas, have been found to suppress glucagon production in alpha cells.[5] Delta cells also release somatostatin which has been found to inhibit glucagon secretion.[5]
Zinc is secreted at the same time as insulin by the beta cells in the pancreas. It has been proposed to act as a paracrine signal to inhibit glucagon secretion in alpha cells. Zinc is transported into both alpha and beta cells by the zinc transporter ZnT8. This protein channel allows zinc to cross the plasma membrane into the cell. When ZnT8 is under-expressed, there is a marked increase in glucagon secretion. When ZnT8 is over-expressed, there is a marked decrease in glucagon secretion. The exact mechanism by which zinc inhibits glucagon secretion is not known.[10]
Insulin has been shown to function as a paracrine signal to inhibit glucagon secretion by the alpha cells.[11] However, this is not through a direct interaction. It appears that insulin functions to inhibit glucagon secretion through activation of delta cells to secrete somatostatin.[12] Insulin binds to SGLT2 causing an increased glucose uptake into delta cells. SGLT2 is a sodium and glucose symporter, meaning that it brings glucose and sodium ions across the membrane at the same time in the same direction. This influx of sodium ions, in the right conditions, can cause a depolarization event across the membrane. This opens calcium channels, causing intracellular calcium levels to increase. This increase in the concentration of calcium in the cytosol activates ryanodine receptors on the endoplasmic reticulum which causes the release of more calcium into the cytosol. This increase in calcium causes the secretion of somatostatin by the delta cells.[12]
Somatostatin inhibits glucagon secretion through the activation of SSTR2, a membrane bound protein that when activated causes a hyperpolarization of the membrane. This hyperpolarization causes voltage gated calcium channels to close, leading to a decrease in intracellular calcium levels. This causes a decrease in exocytosis. In the case of alpha cells, this causes a decrease in the secretion of glucagon.[13]
Serotonin inhibits the secretion of glucagon through its receptors on the plasma membrane of alpha cells. Alpha cells have 5-HT1f receptors which are triggered by the binding of serotonin. Once activated, these receptors suppress the action of adenylyl cyclase, which suppresses the production of cAMP. The inhibition of the production of cAMP in turn suppresses the secretion of glucagon.[5] Serotonin is considered a paracrine signal due to the close proximity of beta cells to alpha cells.[14]
Glucose can also have a somewhat direct influence on glucagon secretion as well. This is through the influence of ATP. Cellular concentrations of ATP directly reflects the concentration of glucose in the blood. If the concentration of ATP drops in alpha cells, this causes potassium ion channels in the plasma membrane to close. This causes depolarization across the membrane causing calcium ion channels to open, allowing calcium to flood into the cell. This increase in the cellular concentration of calcium causes secretory vesicles containing glucagon to fuse with the plasma membrane, thus causing the secretion of glucagon from the pancreas.[5]
Medical significance
High levels of glucagon secretion has been implicated in both Type I and Type II diabetes. In fact, high levels of plasma glucagon is considered an early sign of the development of both Type I and Type II diabetes.[15]
Type I Diabetes
It is thought that high glucagon levels and lack of insulin production are the main triggers for the metabolic issues associated with Type I diabetes, in particular maintaining normal blood glucose levels, formation of ketone bodies, and formation of urea.[16] One finding of note is that the glucagon response to hypoglycemia is completely absent in patients with Type I diabetes.[16] Consistently high glucagon concentrations in the blood can lead to diabetic ketoacidosis,[16] which is when ketones from lipid breakdown build up in the blood, which can lead to dangerously low blood glucose levels, low potassium levels, and in extreme cases cerebral edema.[17] It has been proposed that the reason for the high levels of glucagon found in the plasma of patients with Type I diabetes is the absence of beta cells producing insulin and the reciprocal effect this has on delta cells and the secretion of somatostatin.[16]
Type II Diabetes
Patients with Type II diabetes will have elevated glucagon levels during a fast and after eating.[18] These elevated glucagon levels over stimulate the liver to undergo gluconeogenesis, leading to elevated blood glucose levels.[18] Consistently high blood glucose levels can lead to organ damage, neuropathy, blindness, cardiovascular issues and bone and joint problems.[19] It is not entirely clear why glucagon levels are so high in patients with Type II diabetes. One theory is that the alpha cells have become resistant to the inhibitory effects of glucose and insulin and do not respond properly to them.[18] Another theory is that nutrient stimulation of the gastrointestinal tract, thus the secretion of gastric inhibitory polypeptide and Glucagon-like peptide-1, is a very important factor in the elevated secretion of glucagon.[18]
In other species
There is much controversy as to the effects of various artemisinin derivatives on α-cell-to-β-cell differentiation in rodents and zebrafish.[43] Li et al., 2017 find artemisinin itself forces α⇨β conversion in rodents (via gephyrin)[45] and zebrafish[46] while Ackermann et al., 2018 find artesunate does not[48] and van der Meulen et al., 2018 find the same absence of effect for artemether[49] (although artemether does inhibit ARX).[50] (Shin et al., 2019 further finds no such effect for GABA in rhesus macaque, although GABA is not an artemisinin but has a related action.)[51] Both Eizirik & Gurzov 2018[33] and Yi et al., 2020[36] consider it possible that these are all legitimately varying results from varying combinations of substance, subject, and environment. On the other hand, a large number of reviewers[52] are uncertain whether these are separate effects, instead questioning the validity of Li on the basis of Ackermann and van der Meulen – perhaps GABA receptor agonists as a whole are not β-cell-ergic.[53] Coppieters et al., 2020 goes further, highlighting Ackermann and van der Meulen as publications that catch an unreplicatable scientific result, Li.[47]
See also
References
- Islam, Md. Shahidul, ed. (2015). Islets of Langerhans. Dordrecht: Springer Netherlands. doi:10.1007/978-94-007-6686-0. ISBN 978-94-007-6685-3. S2CID 26869330.
- Jörgens, Viktor (2020). "Paul Langerhans: The Man Who Discovered the Islets". Unveiling Diabetes - Historical Milestones in Diabetology. Frontiers in Diabetes. 29: 25–35. doi:10.1159/000506551. ISBN 978-3-318-06733-0. S2CID 226502826.
- Lane, Michael (1907). "The Cytological Characters of the Areas of Langerhans". The American Journal of Anatomy. VII (3): 409–422. doi:10.1002/aja.1000070304.
- Gromada, Jesper; Franklin, Isobel; Wollheim, Claes B. (2007-02-01). "α-Cells of the Endocrine Pancreas: 35 Years of Research but the Enigma Remains". Endocrine Reviews. 28 (1): 84–116. doi:10.1210/er.2006-0007. ISSN 0163-769X. PMID 17261637.
- Yu, Qian; Shuai, Hongyan; Ahooghalandari, Parvin; Gylfe, Erik; Tengholm, Anders (July 2019). "Glucose controls glucagon secretion by directly modulating cAMP in alpha cells". Diabetologia. 62 (7): 1212–1224. doi:10.1007/s00125-019-4857-6. ISSN 0012-186X. PMC 6560012. PMID 30953108.
- Janah, Lina; Kjeldsen, Sasha; Galsgaard, Katrine D.; Winther-Sørensen, Marie; Stojanovska, Elena; Pedersen, Jens; Knop, Filip K.; Holst, Jens J.; Wewer Albrechtsen, Nicolai J. (January 2019). "Glucagon Receptor Signaling and Glucagon Resistance". International Journal of Molecular Sciences. 20 (13): 3314. doi:10.3390/ijms20133314. ISSN 1422-0067. PMC 6651628. PMID 31284506.
- Stanojevic, Violeta; Habener, Joel F. (2015-10-08). "Evolving Function and Potential of Pancreatic Alpha Cells". Best Practice & Research. Clinical Endocrinology & Metabolism. 29 (6): 859–871. doi:10.1016/j.beem.2015.10.002. ISSN 1521-690X. PMC 4690008. PMID 26696515.
- Verberne, Anthony J. M.; Mussa, Bashair M. (2022-06-01). "Neural control of pancreatic peptide hormone secretion". Peptides. 152: 170768. doi:10.1016/j.peptides.2022.170768. ISSN 0196-9781. PMID 35189258. S2CID 246906606.
- Babic, Tanja; Travagli, R. Alberto (2016-09-23). "Neural Control of the Pancreas". Pancreapedia: The Exocrine Pancreas Knowledge Base. doi:10.3998/panc.2016.27.
- Rutter, Guy A.; Chabosseau, Pauline; Bellomo, Elisa A.; Maret, Wolfgang; Mitchell, Ryan K.; Hodson, David J.; Solomou, Antonia; Hu, Ming (February 2016). "Intracellular zinc in insulin secretion and action: a determinant of diabetes risk?". Proceedings of the Nutrition Society. 75 (1): 61–72. doi:10.1017/S0029665115003237. ISSN 0029-6651. PMID 26365743. S2CID 13936539.
- Asplin, C. M.; Paquette, T. L.; Palmer, J. P. (1981-07-01). "In vivo inhibition of glucagon secretion by paracrine beta cell activity in man". The Journal of Clinical Investigation. 68 (1): 314–318. doi:10.1172/JCI110251. ISSN 0021-9738. PMC 370801. PMID 7019246.
- Vergari, Elisa; Knudsen, Jakob G.; Ramracheya, Reshma; Salehi, Albert; Zhang, Quan; Adam, Julie; Asterholm, Ingrid Wernstedt; Benrick, Anna; Briant, Linford J. B.; Chibalina, Margarita V.; Gribble, Fiona M. (2019-01-11). "Insulin inhibits glucagon release by SGLT2-induced stimulation of somatostatin secretion". Nature Communications. 10 (1): 139. Bibcode:2019NatCo..10..139V. doi:10.1038/s41467-018-08193-8. ISSN 2041-1723. PMC 6329806. PMID 30635569.
- Kailey, Balrik; van de Bunt, Martijn; Cheley, Stephen; Johnson, Paul R.; MacDonald, Patrick E.; Gloyn, Anna L.; Rorsman, Patrik; Braun, Matthias (2012-11-01). "SSTR2 is the functionally dominant somatostatin receptor in human pancreatic β- and α-cells". American Journal of Physiology. Endocrinology and Metabolism. 303 (9): E1107–E1116. doi:10.1152/ajpendo.00207.2012. ISSN 0193-1849. PMC 3492856. PMID 22932785.
- Almaça, Joana; Molina, Judith; Menegaz, Danusa; Pronin, Alexey N.; Tamayo, Alejandro; Slepak, Vladlen; Berggren, Per-Olof; Caicedo, Alejandro (2016-12-20). "Human Beta Cells Produce and Release Serotonin to Inhibit Glucagon Secretion from Alpha Cells". Cell Reports. 17 (12): 3281–3291. doi:10.1016/j.celrep.2016.11.072. ISSN 2211-1247. PMC 5217294. PMID 28009296.
- Smith, Tamar; Gerich, John E. (2003-01-01), "Glucagon Secretion, Regulation of", in Henry, Helen L.; Norman, Anthony W. (eds.), Encyclopedia of Hormones, New York: Academic Press, pp. 74–82, doi:10.1016/b0-12-341103-3/00116-9, ISBN 978-0-12-341103-7, retrieved 2022-03-22
- Bisgaard Bengtsen, Mads; Møller, Niels (August 2021). "Mini‐review: Glucagon responses in type 1 diabetes – a matter of complexity". Physiological Reports. 9 (16): e15009. doi:10.14814/phy2.15009. ISSN 2051-817X. PMC 8371343. PMID 34405569.
- "Diabetic ketoacidosis - Symptoms and causes". Mayo Clinic. Retrieved 2022-03-22.
- Lund, Asger; Bagger, Jonatan I.; Christensen, Mikkel; Knop, Filip K.; Vilsbøll, Tina (December 2014). "Glucagon and Type 2 Diabetes: the Return of the Alpha Cell". Current Diabetes Reports. 14 (12): 555. doi:10.1007/s11892-014-0555-4. ISSN 1534-4827. PMID 25344790. S2CID 6159190.
- "Hyperglycemia in diabetes - Symptoms and causes". Mayo Clinic. Retrieved 2022-03-22.
- Wang, Jigang; Xu, Chengchao; Wong, Yin Kwan; Li, Yujie; Liao, Fulong; Jiang, Tingliang; Tu, Youyou (2019). "Artemisinin, the Magic Drug Discovered from Traditional Chinese Medicine". Engineering. Chinese Academy of Engineering + Higher Education Press (Elsevier). 5 (1): 32–39. doi:10.1016/j.eng.2018.11.011. ISSN 2095-8099. S2CID 116733896.
- Nair, Gopika G.; Tzanakakis, Emmanuel S.; Hebrok, Matthias (2020-06-25). "Emerging routes to the generation of functional β-cells for diabetes mellitus cell therapy". Nature Reviews Endocrinology. Nature Portfolio. 16 (9): 506–518. doi:10.1038/s41574-020-0375-3. ISSN 1759-5029. PMC 9188823. PMID 32587391. S2CID 220064790.
- Kasaragod, Vikram B.; Schindelin, Hermann (2018-09-12). "Structure–Function Relationships of Glycine and GABAA Receptors and Their Interplay With the Scaffolding Protein Gephyrin". Frontiers in Molecular Neuroscience. Frontiers. 11: 317. doi:10.3389/fnmol.2018.00317. ISSN 1662-5099. PMC 6143783. PMID 30258351. S2CID 52187070.
- Gromada, Jesper; Chabosseau, Pauline; Rutter, Guy A. (2018-10-11). "The α-cell in diabetes mellitus". Nature Reviews Endocrinology. Nature Portfolio. 14 (12): 694–704. doi:10.1038/s41574-018-0097-y. ISSN 1759-5029. PMID 30310153. S2CID 52967093.
- Cito, Monia; Pellegrini, Silvia; Piemonti, Lorenzo; Sordi, Valeria (2018). "The potential and challenges of alternative sources of β cells for the cure of type 1 diabetes". Endocrine Connections. European Society of Endocrinology (Bioscientifica). 7 (3): R114–R125. doi:10.1530/ec-18-0012. ISSN 2049-3614. PMC 5861368. PMID 29555660. S2CID 4161356.
- Wang, Qinghua; Ren, Liwei; Wan, Yun; Prud'homme, Gerald J. (2019-01-28). "GABAergic regulation of pancreatic islet cells: Physiology and antidiabetic effects". Journal of Cellular Physiology. Wiley. 234 (9): 14432–14444. doi:10.1002/jcp.28214. ISSN 0021-9541. PMID 30693506. S2CID 59338821.
- Baeyens, Luc; Lemper, Marie; Staels, Willem; De Groef, Sofie; De Leu, Nico; Heremans, Yves; German, Michael S.; Heimberg, Harry (2018-07-01). "(Re)generating Human Beta Cells: Status, Pitfalls, and Perspectives". Physiological Reviews. American Physiological Society. 98 (3): 1143–1167. doi:10.1152/physrev.00034.2016. ISSN 0031-9333. PMC 6088144. PMID 29717931. S2CID 24141451.
- Jacobson, David A.; Shyng, Show-Ling (2020). "Ion Channels of the Islets in Type 2 Diabetes". Journal of Molecular Biology. Elsevier. 432 (5): 1326–1346. doi:10.1016/j.jmb.2019.08.014. ISSN 0022-2836. PMC 7720859. PMID 31473158. S2CID 201715726. NIHMSID 1538588.
- Kasaragod, Vikram Babu; Hausrat, Torben Johann; Schaefer, Natascha; Kuhn, Maximilian; Christensen, Nikolaj Riis; Tessmer, Ingrid; Maric, Hans Michael; Madsen, Kenneth Lindegaard; Sotriffer, Christoph; Villmann, Carmen; Kneussel, Matthias; Schindelin, Hermann (2019). "Elucidating the Molecular Basis for Inhibitory Neurotransmission Regulation by Artemisinins". Neuron. Cell Press. 101 (4): 673–689.e11. doi:10.1016/j.neuron.2019.01.001. ISSN 0896-6273. PMID 30704910. S2CID 72332955.
- Cobo-Vuilleumier, Nadia; Gauthier, Benoit R. (2020). "Time for a paradigm shift in treating type 1 diabetes mellitus: coupling inflammation to islet regeneration". Metabolism. Elsevier. 104: 154137. doi:10.1016/j.metabol.2020.154137. ISSN 0026-0495. PMID 31904355. S2CID 209894697.
- Lorenzo, Petra I; Cobo-Vuilleumier, Nadia; Gauthier, Benoit R (2018). "Therapeutic potential of pancreatic PAX4-regulated pathways in treating diabetes mellitus". Current Opinion in Pharmacology. Elsevier. 43: 1–10. doi:10.1016/j.coph.2018.07.004. ISSN 1471-4892. PMID 30048825. S2CID 51723745.
- Zhang, Jingjing; Liu, Feng (2020). "The De-, Re-, and trans-differentiation of β-cells: Regulation and function". Seminars in Cell & Developmental Biology. Elsevier. 103: 68–75. doi:10.1016/j.semcdb.2020.01.003. ISSN 1084-9521. PMID 31948775. S2CID 210702856.
- Kasaragod, Vikram Babu; Schindelin, Hermann (2019-08-07). "Structure of Heteropentameric GABAA Receptors and Receptor-Anchoring Properties of Gephyrin". Frontiers in Molecular Neuroscience. Frontiers. 12: 191. doi:10.3389/fnmol.2019.00191. ISSN 1662-5099. PMC 6693554. PMID 31440140. S2CID 199465436.
- Eizirik, Decio L.; Gurzov, Esteban N. (2018-09-25). "Can GABA turn pancreatic α-cells into β-cells?". Nature Reviews Endocrinology. Nature Portfolio. 14 (11): 629–630. doi:10.1038/s41574-018-0101-6. ISSN 1759-5029. PMID 30254299. S2CID 52820991.
- Yu, Xin-Xin; Xu, Cheng-Ran (2020-04-01). "Understanding generation and regeneration of pancreatic β cells from a single-cell perspective". Development. The Company of Biologists. 147 (7). doi:10.1242/dev.179051. ISSN 1477-9129. PMID 32280064. S2CID 215749163.
- Lin, Eugene E.; Scott-Solomon, Emily; Kuruvilla, Rejji (2021). "Peripheral Innervation in the Regulation of Glucose Homeostasis". Trends in Neurosciences. Cell Press. 44 (3): 189–202. doi:10.1016/j.tins.2020.10.015. ISSN 0166-2236. PMC 7904596. PMID 33229051. S2CID 227063235. NIHMSID 1643882.
- Yi, Zhao; Waseem Ghani, Muhammad; Ghani, Hammad; Jiang, Wu; Waseem Birmani, Muhammad; Ye, Li; Bin, Liu; Cun, Lang Guan; Lilong, An; Mei, Xiao (2020-01-16). "Gimmicks of gamma‐aminobutyric acid (GABA) in pancreatic β‐cell regeneration through transdifferentiation of pancreatic α‐ to β‐cells". Cell Biology International. International Federation for Cell Biology (Wiley). 44 (4): 926–936. doi:10.1002/cbin.11302. ISSN 1065-6995. PMID 31903671. S2CID 209894386.
- Guney, Michelle A; Lorberbaum, David S; Sussel, Lori (2020). "Pancreatic β cell regeneration: to β or not to β". Current Opinion in Physiology. Elsevier. 14: 13–20. doi:10.1016/j.cophys.2019.10.019. ISSN 2468-8673. PMC 7454996. PMID 32864533. NIHMSID 1545870.
- Hartig, Sean M.; Cox, Aaron R. (2020). "Paracrine signaling in islet function and survival". Journal of Molecular Medicine. 98 (4): 451–467. doi:10.1007/s00109-020-01887-x. PMC 7899133. PMID 32067063. S2CID 211138647.
- Nasteska, Daniela; Viloria, Katrina; Everett, Lewis; Hodson, David J. (2019). "Informing β-cell regeneration strategies using studies of heterogeneity". Molecular Metabolism. Elsevier. 27: S49–S59. doi:10.1016/j.molmet.2019.06.004. ISSN 2212-8778. PMC 6768502. PMID 31500831. S2CID 202406520.
- Krentz, Nicole A J; Shea, Lonnie D; Huising, Mark O; Shaw, James A M (2021). "Restoring normal islet mass and function in type 1 diabetes through regenerative medicine and tissue engineering". The Lancet Diabetes & Endocrinology. The Lancet (Elsevier). 9 (10): 708–724. doi:10.1016/s2213-8587(21)00170-4. ISSN 2213-8587. PMID 34480875. S2CID 237417470.
- Kim, Jeong Joo; Hibbs, Ryan E. (2019). "Bridges between Antimalarials and Synaptic Transmission". Neuron. Cell Press. 101 (4): 546–547. doi:10.1016/j.neuron.2019.01.057. ISSN 0896-6273. PMID 30790532. S2CID 72334998.
- Saleh, Mohamed; Gittes, George K.; Prasadan, Krishna (2021-12-09). "Alpha-to-beta cell trans-differentiation for treatment of diabetes". Biochemical Society Transactions. The Biochemical Society. 49 (6): 2539–2548. doi:10.1042/bst20210244. ISSN 0300-5127. PMC 8786296. PMID 34882233. S2CID 245122056.
- [20] [21] [22] [23] [24] [25] [26] [27] [28] [29] [30] [31] [32] [33] [34] [35] [36] [37] [38] [39] [40] [41] [42]
- Pacios-Michelena, Anabel; Kasaragod, Vikram Babu; Schindelin, Hermann (2021). "Artemisinins and their impact on inhibitory neurotransmission". Current Opinion in Pharmacology. Elsevier. 59: 19–25. doi:10.1016/j.coph.2021.04.008. ISSN 1471-4892. PMID 34051675. S2CID 235248600.
- [41] [44]
- [20] [33] [34] [35] [37] [39] [40] [41] [42] [44]
- Coppieters, Ken; von Herrath, Matthias; Homann, Dirk (2020). "Animal Models of Organ-Specific Autoimmune Disease". In Rose, Noel R.; Mackay, Ian R. (eds.). The Autoimmune Diseases. Elsevier. pp. 493–511. doi:10.1016/b978-0-12-812102-3.00027-0. ISBN 9780128121023. S2CID 243055640.
- [21] [25] [33] [35] [36] [37] [39] [40] [41] [44] [47]
- [33] [35] [36] [39] [40] [41] [47]
- [20] [21] [37] [44] [42]
- [36] [37] [40]
- [27] [28] [29] [32] [33] [34] [35] [38] [39] [44]
- [27] [34]
Further reading
- Quesada, Ivan; Tudurí, Eva; Ripoll, Cristina; Nadal, Ángel (2008-10-01). "Physiology of the pancreatic α-cell and glucagon secretion: role in glucose homeostasis and diabetes". Journal of Endocrinology. 199 (1): 5–19. doi:10.1677/JOE-08-0290. ISSN 0022-0795. PMID 18669612.