Fecundación
La fecundación es el proceso por el cual dos gametos (masculino y femenino) se fusionan durante la reproducción sexual para crear un cigoto con un genoma derivado de ambos progenitores. Los dos fines principales de la fecundación son la combinación de genes derivados de ambos progenitores y la generación de un cigoto.
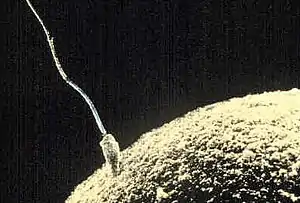
En el caso de las plantas con semillas, hay que diferenciar el fenómeno de la fecundación propiamente dicho (unión íntima de dos células sexuales hasta confundirse sus núcleos respectivos y, en mayor o menor grado, sus citoplasmas),[1] del proceso biológico que lo antecede: la polinización, en el que los granos de polen, desarrollados en las tecas que contiene cada antera de un estambre (órgano floral masculino), son transportados por el viento o los insectos a los estigmas, donde germinan emitiendo un tubo polínico que crece hacia el ovario. En este caso no se trata de gametos, sino de esporas, pues cada grano de polen contiene dos gametos o células reproductoras masculinas.
Proceso
Los detalles de la fecundación son tan diversos como las especies; sin embargo, existen cuatro eventos que son constantes en todas ellas:
- El primer contacto y reconocimiento entre el gameto masculino y el gameto femenino, es de gran importancia para asegurar que ambos sean de la misma especie.
- La regulación de la interacción entre el gameto masculino y el femenino. El gameto femenino solamente puede fecundarse con un gameto masculino. Esto puede lograrse permitiendo que solo un gameto masculino se fusione con el femenino, lo que impedirá que otros también lo hagan.[cita requerida]
- La fusión del material genético proveniente de ambos gametos.
- La formación del cigoto y el inicio de su desarrollo (la embriogénesis).
Modalidades
Según las similitudes y diferencias entre los gametos
- Fecundación isogámica: Unión de dos gametos idénticos en tamaño y estructura. Ocurre solamente en algunos grupos como los protozoos.
- Fecundación anisogámica: Unión de dos gametos distintos, tanto en tamaño como en estructura, uno masculino y otro femenino. Ocurre en la mayor parte de los grupos.
- Fecundación oogámica: gametos muy distintos: el femenino es grande e inmóvil y aporta todas las reservas nutritivas al cigoto, mientras que el masculino es pequeño y móvil.
Según los individuos participantes
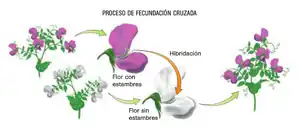
- Fecundación cruzada: fecundación en la que cada gameto procede de un individuo distinto. En algún raro caso, dos individuos se fecundan mutuamente, como ocurre en los caracoles terrestres (o. Pulmonata).
- Autofecundación: cuando los dos gametos proceden del mismo individuo. En las plantas angiospermas, cuyas flores suelen ser hermafrodita, es frecuente la autofecundación, casi siempre combinada con la fecundación cruzada. En algunas especies coexisten con las normales ciertas flores especiales que no se abren, y se produce la fecundación dentro del capullo (cleistogamia).
Según dónde se produce
- Fecundación interna: los espermatozoides llamados introespermatozoide, es un tipo de espermatozoide avanzado (su morfología es de una cabeza que contiene al núcleo y al acrosoma, un cuello donde están los centríolos, la pieza media que es el flagelo rodeado de vainas mitocondriales, y finalmente la pieza terminal que es la continuación del flagelo pero rodeado de vainas fibrosas), estos pasan al cuerpo de la hembra inyectados por órganos copuladores en el curso de un acoplamiento, o bien son tomados por la hembra en forma de un espermatóforo liberado previamente por el macho. Es cuando la unión de los dos gametos o células sexuales (espermatozoide y óvulo) se realiza dentro del cuerpo de la madre en el útero o matriz, de acuerdo a ello los animales se clasifican en:
- Ovíparos: fecundación interna y desarrollo embrionario externo dentro de un huevo provisto de nutrientes y cáscara calcárea, por ej., los Monotremas (ornitorrinco, equidnas), muchas especies de invertebrados, reptiles y aves.
- Ovovivíparos: fecundación interna y desarrollo embrionario incompleto, abandonan el cuerpo de la madre cuando aún son fetos para completar su desarrollo fuera del cuerpo materno, por ej., los Marsupiales (comadrejas, zarigüeyas, canguros). El marsupio (bolsa membranosa) contiene las glándulas mamarias para la alimentación de las crías.
- Vivíparos: fecundación interna y desarrollo embrionario interno, por ej., los Euterios o mamíferos.
- Fecundación externa: propia de los animales acuáticos, existen dos tipos de espermatozoides para la fecundación externa: el acuaespermatozoide que es aquel emitido libremente por el organismo al medio acuático donde vive y que fecunda huevos también libres, y el espermatozoide endoacuático, que también es emitido al medio acuático pero es dirigido por las corrientes inhalantes o de alimentación de la hembra, para fecundar los huevos emitidos por esta y que se mantienen para su incubación fuera del tracto genital. Estos espermatozoides son de tipo primitivo (su morfología se basa en un acrosoma en forma de capuchón, un núcleo subesférico, un número pequeño de mitocondrias específicas con crestas mitocondiales y una cola o flagelo cuya organización es en microtúbulos en 9 pares externos y 1 interno que es originado por un centríolo distal). Es llevada a cabo por casi todos los invertebrados marinos y las siguientes especies:
- Peces: en la reproducción, los óvulos son abandonados por la hembra en el agua al azar y son inmediatamente fecundados por el macho. La fecundación es externa porque ocurre en el agua. Los huevos quedan flotando en el agua, algunos caen y se fijan en el fondo, pero la mayoría sirven de alimento a otros peces. Hay algunas especies de peces, como los tiburones y los peces martillo, que tienen fecundación interna, es decir, que se realiza dentro del cuerpo de la hembra.
- Anfibios: son ovíparos y efectúan una fecundación externa. Los machos abrazan a las hembras y estas al pasar uno o dos días, sueltan los óvulos en el agua. Luego el macho deposita sus espermatozoides sobre los óvulos para lograr la fecundación y el posterior desarrollo de los mismos. Los huevos son blandos y sin cáscara, como estos se secan rápidamente, los depositan en el agua o en sitios húmedos. Las crías no se parecen a sus padres, tienen aspecto de pececillos y respiran como los peces. Cambian de forma, es decir, sufren metamorfosis. Pasan de un estado de renacuajo, donde no tienen patas, a la forma adulta adquiriendo las cuatro patas.
Reconocimiento entre los gametos masculino y femenino
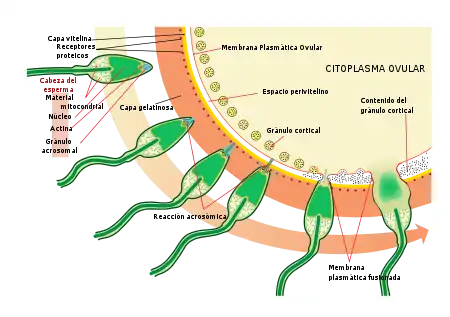
Se producen los siguientes pasos:
- Quimio-atracción desde el gameto femenino hacia el masculino, por medio de la secreción de moléculas solubles que atraen al espermatozoide. En un gran número de especies los espermatozoides son atraídos hacia el gameto femenino, a través de la secreción de una sustancia química por parte de este último.
- Exocitosis de la vesícula acrosómica del gameto masculino de modo que se puedan liberar las enzimas contenidas en esta vesícula.
- Unión del gameto masculino a la capa extracelular que recubre el óvulo. El espermatozoide se une primero al gameto femenino, ocurriendo después la liberación del contenido de la vesícula acrosómica.
- Paso del espermatozoide a través de la matriz extracelular (membrana vitelina en el erizo de mar; zona pelúcida en mamíferos) que recubre al óvulo.
- Fusión de las membranas celulares del gameto femenino y masculino.
Fecundación en plantas

Gimnospermas
Las gimnospermas tiene flores masculinas y femeninas.
La flor femenina tiene una bráctea, una escama y dos óvulos. Se agrupan alrededor de un eje floral, formando el cono femenino. El óvulo contiene un saco embrionario con dos arquegonios, con dos oósferas o gametos femeninos cada uno. Las flores masculinas forman conos masculinos alrededor de un eje floral. Tienen una escama y dos sacos polínicos o microsporangios en los que se forman las células madre que dan lugar a los granos de polen.
Dentro de los granos de polen hay dos anterozoides o gametos masculinos y dos sacos aéreos que favorecen su dispersión hasta llegar a la flor femenina.
El grano de polen tarda hasta un año en germinar tras alcanzar la flor femenina. El tubo polínico se abre lentamente a través de la nucela del óvulo. Al llegar al gametófito femenino atraviesa el cuello del arquegonio, penetra en la oosfera y descarga en ella su contenido. En ese momento se produce la fecundación, uno de los gametos se une con el núcleo de la oosfera para formar el cigoto (diploide). El otro gameto masculino, el núcleo vegetativo y las demás células del arquegonio, degeneran.
El embrión queda rodeado por el endospermo (células de reserva) y protegido por el tegumento del óvulo, que se lignifica.
El embrión no está totalmente maduro hasta dos años después de la aparición de las flores, cuando se liberan las semillas.
En la semilla de pino, el tegumento es diploide, producido por el esporófito maternal, el tejido de reserva (endosperma primario) es haploide pues forma parte del gametófito femenino. El embrión diploide que se forma tras la fecundación es el nuevo esporófito.
Angiospermas
Después de que el carpelo ha sido polinizado, el grano de polen germina en respuesta a un líquido azucarado (principalmente sacarosa) secretado por el estigma maduro. De cada grano de polen, surge un tubo polínico que crea un camino a través del estilo y se dirige hacia el saco embrionario o gametófito femenino de las angiospermas el cual se ubica dentro del óvulo.
A través del tubo polínico viajan los núcleos generativos o gametos masculinos hasta el micrópilo. El tubo polínico lo atraviesa y descarga su contenido cerca de una de las sinérgidas del saco embrionario. Una vez descargado su contenido, los núcleos generativos se fusionan con la oósfera y con los núcleos polares en un proceso conocido como doble fecundación.
Muchos granos de polen pueden alcanzar el estigma y germinar, pero solo uno producirá la fecundación.
Después de haber sido fertilizado, el ovario empieza a crecer y se convertirá en el fruto[3] En los frutos con varias semillas, son necesarios varios granos de polen para fusionarse con cada óvulo.
Fecundación en invertebrados
Entre los invertebrados existen diferentes métodos de reproducción
Reproducción asexual
Este tipo de reproducción se puede llevar a cabo mediante diferentes procesos: mitosis, fisión binaria, gemación y fragmentación, entre otros, seguido por el crecimiento y desarrollo del nuevo individuo.
Existen ventajas adaptativas conferidas a los organismos que presentan este tipo de reproducción. Al poder producir mayor número de descendientes, pueden tomar ventaja en condiciones favorables presentes en cierto ambiente; al explotar las diferentes fuentes alimenticias, el espacio disponible y demás recursos presentes en el medio. Un claro ejemplo de reproducción asexual en invertebrados formadores de colonias son los corales.
Reproducción sexual
Este tipo de reproducción incluye la formación de células haploides, gametos (huevo y esperma) mediante meiosis, y la subsiguiente fusión de dos de estas células para poder formar un cigoto diploide. La mayoría de los invertebrados liberan sus gametos al medio donde habitan, y este generalmente es el agua, y tiene lugar la fertilización externa.
En estos organismos la sincronía en la liberación de los gametos es crítica, debido a que deben liberarse tanto huevos como esperma al mismo tiempo para que pueda ocurrir la fertilización. Algunos de los factores que se cree están relacionados con la sincronía de la liberación de gametos en los animales marinos son la temperatura del agua, la luz, la abundancia de fitoplancton, el ciclo lunar y la presencia de organismos de otras especies.
Es importante mencionar también que, aunque la mayoría de los invertebrados siguen este patrón de fecundación externa, también existen los que llevan a cabo este proceso internamente. Este tipo de organismos deben tener un mayor desarrollo de sus sistemas reproductores, que permitan facilitar este proceso.
Fecundación en mamíferos
La fecundación en mamíferos es interna. El papel que juega el tracto reproductivo femenino es muy importante porque facilita que los espermatozoides lleguen al extremo de la trompa de Falopio, gracias al movimiento muscular que ejerce el útero.
Cuando los espermatozoides alcanzan la ampolla, adquieren competencia (la pueden perder si se quedan alrededor de esta demasiado tiempo). Los espermatozoides pueden tener diferentes porcentajes de supervivencia dependiendo de su localización dentro del tracto reproductivo femenino.[4]
En mamíferos, la fertilización hace que se active el proceso por el cual se completa la meiosis en el huevo y el juego de cromosomas de la madre se convierte en el pronúcleo del huevo.
El huevo está cubierto de varias capas: la membrana plasmática, gránulos corticales y la zona pelúcida. El esperma es móvil, está diseñado para activar el huevo y al mismo tiempo insertar su núcleo al citoplasma del huevo. Tanto el huevo como el espermatozoide están estructuralmente especializados para la fertilización. El huevo está especializado en prevenir la fertilización de más de un espermatozoide, mientras que el espermatozoide está especializado en acercarse lo máximo posible a la membrana del óvulo. Al fusionarse el espermatozoide y el óvulo, se activa un mecanismo de bloqueo en el huevo en el cual se previene que otro espermatozoide se fusione (bloqueo de polispermia). Esto es necesario porque si más de un espermatozoide se fusionara con el huevo, habría juegos extra de cromosomas y centrosomas, resultando en un desarrollo anormal.
Proceso de capacitación
Después de la entrada del esperma en el tracto reproductivo femenino ocurre un proceso de capacitación, el cual busca facilitar la fertilización eliminando ciertos factores inhibitorios y barreras. La capacitación como tal, se refiere a los cambios fisiológicos por los cuales el espermatozoide llega a ser competente para fecundar el gameto femenino. Los cambios moleculares de la capacitación son todavía muy desconocidos. Hasta el momento se han identificado cuatro cambios importantes:
- Alteración de la membrana celular del espermatozoide, eliminación de colesterol por proteínas de albúmina en el tracto reproductivo femenino. Esto al parecer aumenta el pH que permite que el espermatozoide experimente acción acrosómica.
- También se pierden hidratos de carbono en la superficie espermática, lo cual al parecer facilita el reconocimiento para las proteínas de la zona pelúcida.
- El potencial de la membrana celular del espermatozoide se vuelve más negativo cuando los iones de potasio dejan el espermatozoide. Al parecer facilita que los canales de calcio se abran y así entra el calcio al espermatozoide. Los iones de bicarbonato y calcio están relacionados con la producción de cAMP y facilitan los eventos de fusión de membrana.
- Fosforilación de proteínas
Se ha documentado (Timothy Smith 1998 y Susan Suárez 1998) que antes de ingresar a la ampolla de la trompa de Falopio donde ocurre la fertilización los espermatozoides sin capacitarse se unen a las membranas de las células de la trompa en el istmo hasta que completan la capacitación y se desprenden. Al parecer esta unión temporal alarga la vida de los espermatozoides y vuelve más lenta la capacitación. Esto podría tener como objetivo evitar la poliespermia, y maximizar la probabilidad de que haya espermatozoides en la ampolla para encontrar el gameto femenino cuando la eyaculación ocurrió días antes de la ovulación.[4]
Motilidad de los espermatozoides
En distintas regiones del tracto reproductivo femenino se secretan distintas moléculas, las cuales pueden influenciar la motilidad de los espermatozoides. Por ejemplo, en algunos casos de roedores, los espermatozoides al pasar del útero a las tropas se Falopio se hiperactivan, nadando a velocidades mayores. La hiperactivación parece estar relacionada con cAMP de un canal de calcio en la cola de los espermatozoides. Esto facilita la motilidad a través de fluidos viscosos como los que se encuentran en las trompas de Falopio. La hiperactividad y la hialuronidasa permite que los espermatozoides atraviesen la capa del cúmulo.
Otros factores secretados en el oviducto proporcionan el componente direccional del movimiento de los espermatozoides, se especula que estos factores quimiotácticos son secretados por el folículo ovárico. También se ha visto que solo los espermatozoides capacitados logran ser atraídos por el fluido folicular quimiotáctico.[4]
.jpg.webp)
Barreras a superar por los espermatozoides
La capacitación permite al espermatozoide sobrepasar varias barreras y lograr la fecundación. La primera barrera al llegar al huevo es una capa de células cúmulus en ácido hialurónico. La actividad de la hialuronidasa en la superficie de la cabeza del espermatozoide lo ayuda a penetrar esta barrera. La segunda barrera es la zona pelúcida, la cual es una capa de glucoproteínas. El espermatozoide logra penetrar esta barrera gracias a la reacción acrosómica (liberación de contenidos de la vesícula acrosómica ubicada en la cabeza del espermatozoide).[5]
La zona pelúcida
La zona pelúcida juega un papel análogo a la membrana vitelina de los invertebrados. Esta matriz extracelular, la cual es sintetizada por el ovocito tiene dos funciones principales: unir el espermatozoide y comenzar la reacción acrosómica. Esta capa de la zona pelúcida tiene tres glucoproteínas principales ZP1, ZP2 y ZP3. Esta última es un receptor especie-específico para la unión del espermatozoide.
Unión del espermatozoide con el huevo
La membrana celular que recubre la cabeza del espermatozoide tiene varias proteínas (un ejemplo es la SED1), estas proteínas se pueden unir cientos de glucoproteínas de ZP3 de la zona pelúcida (por cadenas de carbohidratos enlazadas por serina y treonina. Al experimentar la reacción acrosómica sobre la zona pelúcida, los espermatozoides concentran sus enzimas proteolíticas sobre el punto de adhesión y digieren un agujero a través de esta capa extracelular. Cuando ZP3 se enlaza a los receptores en la membrana celular del espermatozoide se activa la reacción acrosómica. Una de las proteínas del espermatozoide enlazada es la galactosiltranferasa-I, una enzima intramembranosa cuyos sitios activos miran hacia el exterior y se une a los residuos de carbohidratos de la ZP3. Esto a la vez activa las proteínas G específicas de la membrana del espermatozoide, las cuales activan una cascada que abre los canales de calcio y provoca la exocitosis del contenido acrosómico, esto está mediado por el calcio de la vesícula acrosómica.
El contenido acrosómico incluye β-N-acetilglucosaminidasa y varias proteasas las cuales rompen cadenas de oligosacáridos de las glucoproteínas de la zona pelúcida. Esto permite que el espermatozoide perfore la zona pelúcida y se acerque a la membrana plasmática del huevo. Para la perforación continua sin perder la adhesión con la matriz extracelular se debe logar una unión secundaria a la zona pelúcida, a través de ZP2.[5]
La reacción acrosómica también expone unas proteínas de la superficie del espermatozoide que se pueden unir con la membrana del huevo y las cuales permiten la fusión de ambas membranas. Otro componente importante es la proteína fertilina la cual se une a un receptor tipo integrina en la membrana. Un receptor del huevo para reconocer el espermatozoide es la proteína CD9 la cual inicia la integración huevo-espermatozoide y que luego es un factor crítico en la fusión de los miocitos (precursores musculares) para formar el miotubo multinucleado del músculo estriado. En mamíferos el espermatozoide no contacta al gameto femenino en su extremo, sino sobre el costado de la cabeza, en la región dominio ecuatorial de la cabeza del espermatozoide.[4]
Para evitar la polispermia, apenas llega el primer espermatozoide a la membrana plasmática del huevo y comienza la integración, se liberan los gránulos corticales los cuales contienen enzimas que evitan la unión de otros espermatozoides con la zona pelúcida. A diferencia de otros organismos como los erizos de mar, en mamíferos no se presenta un cambio de potencial de membrana.
Activación del huevo a partir de la fertilización
La activación del huevo a partir de la fertilización activa una serie de eventos que resultan en el comienzo del desarrollo. Los eventos principales son: el huevo completa meiosis, se unen los núcleos del huevo y el espermatozoide para formar un cigoto diploide, y el huevo fertilizado entra a mitosis. En el caso de ratones y humanos las membranas de los pronúcleos desaparecen antes de la unión de estos.
Al igual que en erizos de mar la activación del huevo está relacionado con la liberación de iones libres de calcio en el huevo (produciendo una ola de calcio la cual es necesaria y suficiente para comenzar el desarrollo). La ola de calcio comienza en el punto por el cual entró el espermatozoide y cruza todo el huevo. Hay oscilaciones en la concentración de calcio por varias horas después de la fertilización. El mecanismo por el cual se inicia la liberación de calcio no es conocido, pero se cree que el espermatozoide induce factores específicos de proteína que inicia la liberación de calcio después de la fusión de los gránulos corticales. Al aumentar las concentraciones de calcio se inicia el desarrollo del huevo fertilizado al activar proteínas relacionadas con el ciclo celular.[5]
Resumen
Este proceso se desarrolla mediante los siguientes pasos:
- Se considera que los espermatozoides son atraídos quimio-tácticamente hacia el gameto femenino, mediante moléculas emitidas por células de los folículos alrededor de estas.
- Los espermatozoides, con el acrosoma intacto, atraviesan la zona del cúmulo y se unen de manera específica a la zona pelúcida o cubierta extracelular del óvulo. Se han identificado tres proteínas que están relacionas con la unión del espermatozoide a esta matriz; ZP1, ZP2 y ZP3. Esta última actúa como el principal y más importante receptor de los gametos masculinos, además de la habilidad que tiene de inducir la reacción acrosómica.
- Después de la unión, el espermatozoide llevará a cabo la reacción acrosómica, a través de una exocitosis celular.
- Debido a esta reacción, el espermatozoide puede ahora perforar la zona pelúcida, pasar a través de ella y llegar a la membrana celular del óvulo. El paso por medio de las diferentes capas de membranas extracelulares del óvulo depende únicamente del movimiento propio del gameto masculino, ayudado por enzimas del acrosoma.
- Al finalizar el proceso, el espermatozoide se une a la membrana plasmática del óvulo y se fusiona con esta.
Fecundación en el ser humano
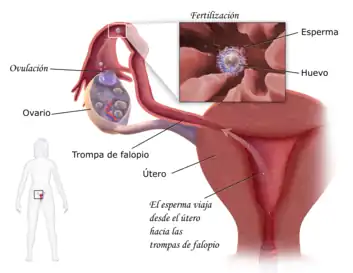
El proceso de fecundación se inicia con el contacto entre los gametos. Dicho encuentro ocurre habitualmente en la región de la ampolla que ocupa el tercio externo de la trompa uterina.
Primero el espermatozoide penetra la corona radiada del ovocito, hasta entrar en contacto con la zona pelúcida. Se genera en la cabeza del espermatozoide, la reacción acrosómica que le permite entrar a la zona pelúcida.
Tanto la cola del espermatozoide como enzimas de la mucosa tubárica contribuyen con la hialuronidasa acrosómica para abrirle el paso al espermatozoide por la zona pelúcida.[cita requerida]
Además de la hialuronidasa, otras enzimas del acrosoma pueden contribuir a la penetración de la zona pelúcida: ciertas esterasas, acrosinas como la arrocina y la neuraminidasa.
Se necesita más de un espermatozoide para lograr fecundar al ovocito (efecto colaborativo). La red de la zona granulosa no es fácil de atravesar para un espermatozoide. Los espermatozoides tienen haluiorinasa para facilitar el paso hasta llegar a la zona pelúcida. Algunos espermatozoides van soltando la cápsula de la vesícula acrosómica dejando un camino. Es imprescindible poseer el acrosoma intacto para pefrorar el ovocito, pues sin acrosoma el espermatozoide no podrá atravesar la membrana del óvulo. Por lo tanto se necesitan espermatozoides que liberen sus enzimas antes de llegar al ovocito para que estas degraden la zona granulosa y así algún espermatozoide consiga llegar a la zona pelúcida con su acrosoma intacto y pueda entonces unirse al ovocito.[cita requerida]
Cuando el espermatozoide se encuentra con la zona pelúcida se une a ella. Se produce entonces la reacción acrosómica inducida o la proteína ZP3. La membrana celular del espermatozoide se fusiona con la membrana exterior del acrosoma y el contenido se libera a través de unos poros. Las enzimas liberadas (acrosina principalmente, parecida a la tripsina) van disolviendo la zona pelúcida y permitiendo el paso del espermatozoide empujado por el flagelo a una velocidad de 1 micra (μm) por minuto. La zona pelúcida tiene 17±4.0 μm de grosor y los espermatozoides humanos tardan atravesarla entre 10 y 30 minutos.[cita requerida]
La unión a la zona pelúcida es un paso decisivo de la fecundación. Cuando la reacción ha terminado el espermatozoide está recubierto por la membrana interna del acrosoma. Este cambio es esencial para el contacto posterior con el ovocito. La zona postacrosómica entra en contacto con las microvellosidades del ovocito. A continuación se funden las membranas y entran en contacto los citoplasmas. El contenido del espermatozoide entra dentro del citoplasma del ovocito. [cita requerida]
Sin una correcta reacción acrosómica la zona postacrosómica no entra en contacto de forma adecuada con el ovocito.
Tanto la pieza media como el flagelo del espermatozoide pueden entrar en el ovocito. Tan pronto como un espermatozoide aborda el ovocito se debe evitar la entrada de otro para evitar la polispermia. Esto se provoca con dos mecanismos:[cita requerida]
- La unión dispara una veloz ola despolarizadora en el ovolema que altera la superficie. Una entrada masiva de iones Na+ impide nuevas fusiones de membrana. En un bloqueo temprano, inmediato y transitorio.
- Una segunda oleada despolarizadora provocada por iones Ca++ provoca el vaciado de miles de vesículas corticales al espacio perivitelino. Son lisosomas que contienen numerosas enzimas que endurecen la zona pelúcida. El endurecimiento impide de forma permanente la entrada de más espermatozoides y protege al cigoto.
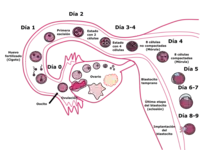
Es notable la capacidad que tienen los zooides para levantar el segundo bloqueo meiótico, que imposibilitaba al ovocito II a continuar con el proceso de meiosis. Una vez que el zooide penetra la zona pelúcida y toma contacto con la membrana plasmática del ovocito II, se produce una intensificación del metabolismo respiratorio de esta célula, se forma el segundo cuerpo polar, que es una célula más pequeña y sin material genético, producto de la conclusión del proceso meiótico.
A partir del momento de la fecundación se restablece el número cromosómico y se define el sexo del embrión, según si el espermatozoide porta un cromosoma X o un cromosoma Y (los ovocitos solo pueden llevar un cromosoma X).
Es común la idea de que para fecundar a un único ovocito se necesita un solo espermatozoide. Se sabe que es necesaria la aportación de varios espermatozoides para poder fecundar un ovocito. La hialurasa se secreta solamente si el espermatozoide llega a la zona pélucida, pero a veces hay espermatozoides que llevan a cabo la reacción acrosómica antes de tiempo, de forma que van degradando el ácido hiarulónico que rodea al ovocito, van despejando el camino a otros espermatozoides. Así pues, se necesitan varios espermatozoides para llegar a fecundar un único ovocito. Además, el movimiento hiperactivo conjunto se cree que también ayuda en la penetración en el ovocito.
- Huevo o cigoto
Después de haber ocurrido la fecundación el cigoto (previamente llamado óvulo cuando el espermatozoide hace el ingreso y se ha reanudado la metafase II) comienza a experimentar una serie de sucesos como la segmentación
Los términos fecundación y concepción
Si bien de unos años a la fecha se ha querido distanciar estos términos para referirse como etapas distintas del proceso de gestación,[6] los términos fecundación y concepción han sido considerados como sinónimos, en tanto que la palabra fecundación hace referencia a todo el proceso desde que los espermatozoides entran al útero, viajan y encuentran al óvulo.[7] En cambio, concepción es el momento exacto en el que el espermatozoide entra en el ovocito y desencadena una serie de cambios que darán lugar al desarrollo del embrión.[7]
En el Diccionario médico-biológico, histórico y etimológico, editado por la Universidad de Salamanca, se define a la concepción como el comienzo del embarazo, abarcando la unión del óvulo y el espermatozoide, y en anidamiento o implantación del huevo en el útero. Su origen viene del latín "con- unión, contacto, acción completa; cep- coger, recibir; y tion- acción.[8]
Véase también
Referencias
- Font Quer, P. 1977. Diccionario de botánica. Barcelona: Ed. Labor. p. 459.
- Pierce, Benjamin A. (7 de julio de 2009). Genética: Un enfoque conceptual. Ed. Médica Panamericana. ISBN 978-84-9835-216-0. Consultado el 12 de marzo de 2023.
- Johnstone, Adam (2001). Biology: facts & practice for A level. Oxford University Press. p. 95. ISBN 0-19-914766-3.
- Gilbert, S. 2005. Biología del desarrollo. 7.ª Edición. Sinauer Assoc., Sunderland. Capítulo 7 Fecundación: el comienzo de un nuevo organismo; páginas 207-221.
- Wolpert, L. 2007. Principles of Development. Third Edition, Oxford, United States Chapter 11. Germ cells, fertilization, and sex, pages 432-436.
- Mundo de Obstetricia en Blogspot (23 de julio de 2013). «Diferencia entre concepción y fecundación». Consultado el 23 de julio de 2013.
- «Embriología».
- Universidad de Salamanca (23 de julio de 2013). «definición de concepción y su relación con la fecundación». Diccionario médico-biológico, histórico y etimológico. Archivado desde el original el 15 de julio de 2013. Consultado el 23 de julio de 2013.
Bibliografía
- Austin, C. (1974). Scientific and clinical aspects of fertilization and implantation. Proc. Roy. Soc. Med., 67.
- Brusca, R. y Brusca, G. (2002). Invertebrates. 2a. ed. Sinauer Assoc., Sunderland.
- Gilbert, S. F. (2005). Biología del desarrollo. 7a. ed. México: Panamericana.
- Kiipker, W., Diedrich, K. y Edwards, R. (1998). Principles of mammalian fertilization. Human Reproduction, 13.
- Longo, F. J. (1997). Fertilization. 2nd. ed. London: Chapman & Hall.
- Wassarman, P., Jovine, L., Litschere. 2001. A profile of fertilization in mammals. Nature Cell Biology, 3.
Enlaces externos
 Wikcionario tiene definiciones y otra información sobre fecundación.
Wikcionario tiene definiciones y otra información sobre fecundación.
El Diccionario de la Real Academia Española tiene una definición para fecundación.
| Control de autoridades |
|
|---|
 Datos: Q14890574
Datos: Q14890574 Multimedia: Fertilisation / Q14890574
Multimedia: Fertilisation / Q14890574