Struthiomimus altus
Struthiomimus altus es la única especie conocida del género extinto Struthiomimus (lat., "imitador del avestruz") de dinosaurio terópodo ornitomímido[1] que vivió a finales del período Cretácico, hace 76 y 70 millones de años, durante el Campaniense, en lo que hoy es Norteamérica. Sus restos se han encontrado en Alberta, Canadá y Wyoming, Estados Unidos. Su nombre genérico se deriva del latín, pasando por el griego στρουθιον, strouthion, que significa «avestruz», y μιμος, mimos, que significa «mimo» o «imitador», en el sentido de «similar a un avestruz», en castellano estrutiomimo. El término específico altus, de Struthiomimus altus, proviene del latín, que significa «alto» o «noble».
| Struthiomimus altus | ||
|---|---|---|
| Rango temporal: Cretácico superior | ||
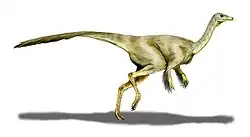 Recreación de S. altus. | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| Infraorden: | Ornithomimosauria | |
| Familia: | Ornithomimidae | |
| Género: |
Struthiomimus Osborn, 1917 | |
| Especie: |
S. altus (Lambe, 1902) | |
| Sinonimia | ||
| ||
Descripción
Struthiomimus medía más de 3 metros, sin embargo no pesaba mucho debido a sus huesos huecos y a la ligereza general de su esqueleto. Sus patas indican que pudo ser un excelente corredor, con gran afinidad con el avestruz actual. Struthiomimus es uno de los dinosaurios más comunes en el Parque Provincial del Dinosaurio. Su abundancia sugiere que fue herbívoro u omnívoro en vez de carnívoro.
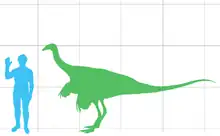
Un Struthiomimus adulto alcanzaba cerca de 4,3 metros de largo y 1,4 de alto hasta la cadera, pesando alrededor de 150 kilogramos,[2] un tamaño y masa típico de los ornitomímidos , que diferían de géneros estrechamente relacionados como Ornithomimus y Gallimimus en proporciones y detalles anatómico.[3]
En general el cuerpo exhibía una constitución frágil y delgada, presentando una cabeza proporcionalmente pequeña en cuellos largos, que constituían aproximadamente el 40% de la longitud del cuerpo por delante de las caderas.[4] El cráneo estaba conformado por unas órbitas, cuencas oculares, relativamente grandes. Tenía un hocico sin dientes que presentaba en la punta del premaxilar un pico recto. La mandíbula inferior poseía en su parte baja 2 pares diminutos de fenestras mandibulares. Con el cuello levantado hacia arriba hubiera alcanzado los 2 metros en altura. Tanto los miembros delanteros como los traseros eran largos, así como lo era su ya mencionado cuello, y su cola, que terminaba en una punta rígida. Las patas estaban diseñadas principalmente para la velocidad, y le hubieran servido de mucha ayuda ante los predadores.
Su columna vertebral constaba de 10 vértebras del cuello, 13 vértebras del dorso, 6 vértebras de la cadera y unas 35 vértebras de la cola.[5] Sus colas eran relativamente rígidas y probablemente las usaban para mantener el equilibrio.[6] Tenían brazos y manos largos y delgados, con huesos del antebrazo inmóviles y oponibilidad limitada entre el primer dedo y los otros dos.[7] Como en otros ornitomímidos, pero inusualmente entre los terópodos, los tres dedos tenían aproximadamente la misma longitud y las garras estaban ligeramente curvadas. Henry Fairfield Osborn , al describir un esqueleto de S. altus en 1917, comparó el brazo con el de un perezoso.[6] Estas podrían haber sido adaptaciones para sostener las plumas de las alas.[8] Es probable que tuviera plumas por todo el cuerpo. Struthiomimus difería de sus parientes cercanos solo en aspectos sutiles de la anatomía. El borde del pico superior era cóncavo en Struthiomimus, a diferencia de Ornithomimus, que tenía bordes de pico rectos.[9] Struthiomimus tenía manos más largas en relación con el húmero que otros ornitomimidos, con garras particularmente largas.[4] Sus extremidades anteriores eran más robustas que las de Ornithomimus.[9]

Sus hábitos alimentarios son todavía desconocidos y sobre ellos circulan varias hipótesis. Lo más probable, según varios estudios, es que haya sido omnívoro o herbívoro. Además, en un espécimen se han hallado gastrolitos (piedras que ayudaban digerir la comida en el estómago cuando ésta no se trituraba), algo común en dinosaurios herbívoros, aunque esto no determinaría con certeza su alimentación al haberse encontrado también en dinosaurios carnívoros tales piedras. Tenía manos prensiles con 3 garras curvas, de las cuales se teoriza que tal vez pudieron ayudar a agarrar frutas o ramas con hojas; sin embargo, también pudieron servir para desgarrar a sus presas.[10]
Pudo haber estado recubierto por plumas, según se puede especular por su posición filogenética, pero esto no está demostrado con certeza, ya que no se han descubierto impresiones fosilizadas de algún integumento del estrutiomimo que sustenten esto. También se han hallado restos de varios individuos juntos, lo que lleva a creer que estos dinosaurios vivían en manadas.
Historia

En 1901, Lawrence Lambe encontró algunos restos incompletos, el holotipo CMN 930, que nombró al año siguiente como Ornithomimus altus, ubicándolos dentro del mismo género que los restos descritos por Othniel Marsh en 1890. Sin embargo, en 1914, un esqueleto casi completo, catalogado AMNH 5339, fue descubierto por Barnum Brown en los yacimientos del río Red Deer, y fue oficialmente descrito como un género separado por Henry Fairfield Osborn en 1916, aunque igualmente en su estudio lo consideró un subgénero del Ornithomimus.[6] Dale Russell convirtió a Struthiomimus en un género completo en 1972, al mismo tiempo que le refirió varios otros especímenes, entre ellos, AMNH 5375, AMNH 5385, AMNH 5421, CMN 8897, CMN 8902 y ROM 1790, todos esqueletos parciales.[11] La especie tipo, S. altus, se conoce a partir de varios esqueletos y cráneos,[4] En 1916, Osborn también cambió el nombre de Ornithomimus tenuis Marsh 1890 a Struthiomimus tenuis.[6] Este se considera hoy un nomen dubium. En 2016, ROM 1790 se convirtió en el holotipo de un nuevo género y especie, Rativates evadens.[12]
En los años siguientes, William Arthur Parks nombró otras cuatro especies de Struthiomimus. La primera de ellas en 1926 la llamo Struthiomimus brevetertius,[13] la cual pasó a ser Dromiceiomimus brevitertius.[11] Posteriormente, en 1928, nombró a Struthiomimus samueli,[14] que pasó a ser llamado Dromiceiomimus samueli.[11] En 1933, nombró a Struthiomimus currellii,[15] que fue recombinado como Ornithomimus curellii por Russell y Chamney en 1967,[16] luego fue mal escrito como Ornithomimus currelli por Eberth en 1997[17] y sinonimizado subjetivamente con Ornithomimus edmontonicus por Steel en 1970.[18] En ese mismo trabajo, nombró a Struthiomimus ingens,[15] fue recombinado como Ornithomimus ingens por Russell y Chamney en 1967,[16] luego fue sinonimizado subjetivamente con Dromiceiomimus brevetertius por Osmólska et al. en 1972[19] y, por último, fue sinonimizado subjetivamente con Ornithomimus edmontonicus por Makovicky et al. en 2004.[20] En 1997 Donald Glut mencionó el nombre Struthiomimus lonzeensis.[21] Este fue probablemente un lapsus calami, una equivocación involuntaria al escribir, un error de Ornithomimus lonzeensis.
Struthiomimus altus proviene de la Formación Oldman del Campaniano Superior, era Judithiano.[22] Restos de Struthiomimus provienen también de la Formación Dinosaur Park, que data de finales del Campaniense. Hay restos de Struthiomimus procedentes de la Formación Cañón Herradura, la cual data de finales del Campaniense y comienzos del Maastrichtiense , que forman la era Edmontoniana. Debido a que las faunas de dinosaurio muestran rápida productividad, es probable que estos especímenes del Struthiomimus de épocas más jóvenes pertenezcan a una nueva especie, pero hay muy poco material conocido para afirmar que estos especímenes pertenecen o no a S. altus, aunque no se les a dado un nuevo nombre.[22][9] De los especímenes hallados en Alberta, algunos fueron encontrados en y bajo pilas de ceniza, sugiriendo que un incendio forestal pudo haberlos matado.
Los especímenes adicionales de Struthiomimus de la Formación Lance inferior y equivalentes son más grandes, similares a Gallimimus en tamaño y tienden a tener garras más rectas y alargadas, similares a las que se ven en Ornithomimus. Un espécimen relativamente completo de la Formación Lance, BHI 1266, fue referido originalmente como Ornithomimus sedens, nombrado por Marsh en 1892[23] y luego clasificado como Struthiomimus sedens.[24] Un artículo de 2015 de van der Reest et al. enumeró BHI 1266 como Ornithomimus sp.,[25] mientras que otro artículo del mismo año consideró el espécimen Struthiomimus sp. pendiente de una revaluación de ambos géneros.[22]
Clasificación
Struthiomimus es un miembro de la familia Ornithomimidae, un grupo que también incluye Anserimimus, Archaeornithomimus, Dromiceiomimus, Gallimimus, Ornithomimus y Sinornithomimus.
Así como los restos fósiles del Struthiomimus fueron asignados erróneamente al Ornithomimus, la familia a la cual pertenece el género, Ornithomimidae, también sufrió cambios a través de los años. Por ejemplo, O.C. Marsh incluyó inicialmente a Struthiomimus en Ornithopoda, un gran clado de dinosaurios que no están estrechamente relacionados con los terópodos.[26] Cinco años después, Marsh clasificó a Struthiomimus en Ceratosauria,[27][28] En 1891, Baur colocó el género dentro de Iguanodontia,[29] Mas cerca de la actualidad, D A. Russell y Z. Dong en 1993, refirieron a Struthiomimus a Oviraptorosauria.[30] Pero posteriormente hubo numerosos estudios que los aceptaron a Struthiomimus dentro de Coelurosauria,[31][32][33] modificando así los taxones del grupo.[34] Reconociendo la diferencia de ornitomímidos comparada a otros terópodos, Rinchen Barsbold ubicó a los ornitomímidos dentro de su propio infraorden, Ornithomimosauria, en 1976.[35] [20]
La amplitud de Ornithomimidae y Ornithomimosauria variaron con los diferentes autores. Paul Sereno, por ejemplo, utilizó Ornithomimidae para incluir a todos los Ornithomimosauria en 1998, y subsecuentemente cambió a una definición más exclusiva que anidó a Ornithomimidae, los ornitomimosaurios avanzados, dentro del gran grupo más generaql de Ornithomimosauria,[36] un esquema de clasificación que fue adoptado por muchos otros autores a comienzos de los 2000s. Struthiomimus forma la familia junto con el Anserimimus, Archaeornithomimus, Dromiceiomimus, Gallimimus, Ornithomimus y Sinornithomimus, teniendo un poco más de similitud con el Dromiceiomimus y el Ornithomimus, con los cuales se ha confundido ya varias veces. Anatómicamente, los Ornithomimosauria representan el grupo de dinosaurios no avianos más estrechamente relacionados con las avestruces.
Filogenia
El siguiente cladograma está basado en Turner, Clarke, Ericson y Norell, 2007.[37] El nombre de los clado sigue la definición de Sereno, 2005.[38]
| Ornithomimosauria |
| ||||||||||||||||||||||||||||||||||||||||||
Sin embargo Xu et al., 2011 propone:[39]
| Ornithomimidae |
| ||||||||||||||||||||||||||||||||||||
Paleobiología
En un estudio de 2001 realizado por Bruce Rothschild y otros paleontólogos, se examinaron cincuenta huesos del pie referidos a Struthiomimus en busca de signos de fractura por estrés, pero no se encontró ninguno.[40]
Postura
Struthiomimus fue uno de los primeros terópodos imaginados desde el principio con una postura horizontal. Osborn en 1916 dejó que el animal se representara intencionalmente con una cola elevada.[6] Esta vista más nueva creó una imagen que recuerda mucho más a las aves no voladoras modernas, como el avestruz al que se refiere el nombre de este dinosaurio, pero solo mucho más tarde se aceptaría para todos los terópodos.

Struthiomimus normalmente caminaba y corría con la cola horizontal, sin arrastrarla. Pero hay algunos grupos de terópodos avanzados que son capaces de mantener una postura con el cuerpo a 45 grados o menos de la dirección vertical y la cola arrastrándose a lo largo del suelo, una pose conocida como "trípode". Por tanto, Struthiomimus, al ser un ornitomímido, es un dinosaurio que podía mantener su postura de esa manera gracias a las articulaciones de su cadera, pero quizá solo por tiempos poco prolongados.

La antigua postura con la que se representaba a Struthiomimus era incorrecta al ser siempre trípode en la locomoción, ya que muy probablemente sólo la mantenía para tareas de vigilancia, realizar despliegues sexuales o simplemente observar a sus alrededores. Lo máximo que Struthiomimus pudo alzarse se muestra en la imagen derecha, pues si su postura giraba más grados se hubiera dislocado o debilitado varias articulaciones.
Alimentación

Hay mucha discusión acerca de los hábitos alimenticios de Struthiomimus.[41] Debido a su pico de bordes rectos, se cree que Struthiomimus pudo ser probablemente omnívoro o herbívoro. Algunas teorías sugieren que pudo ser un habitante de orillas en las que comía insectos, cangrejos, camarones y posiblemente huevos de otros dinosaurios. Algunos paleontólogos señalan que es más posible que sea un carnívoro, ya que está clasificado dentro del grupo de los terópodos.[42][11] Esta teoría no había sido rechazada hasta que Osborn, quien describió y nombró al dinosaurio, propuso que probablemente se alimentaba de yemas y brotes de árboles, arbustos y otras plantas,[3] usando sus miembros delanteros para agarrar ramas y su cuello largo para seleccionar objetos particulares. La hipótesis de que Struthiomimus era herbívoro encuentra sustento, además, en la estructura inusual de sus manos. El segundo y tercer dedo tenían la misma longitud, no podían funcionar independientemente y es probable estuviesen unidos por piel . Esto indica que la mano fue utilizada como "garfio", para poner ramas o frondas de helecho a su alcance. La estructura de la cintura escapular no permitía una gran elevación del brazo ni estaba optimizada para un alcance bajo. La mano no podía flexionarse por completo para un movimiento de agarre o extenderse para rastrillar. Esto indica que la mano se usaba como "gancho" o "abrazadera", para poner al alcance ramas o frondas de helecho a la altura de los hombros.[10] Sin embargo, estas adaptaciones podrían haber sido utilizadas para el apoyo de las plumas de las alas.[8]
Velocidad
Los miembros traseros de Struthiomimus fueron largos, muy parecidos a los de un avestruz, sobre todo porque la tibia era mucho más grande que el fémur, un rasgo notable en el esqueleto de las aves. Los músculos de las articulaciones garantizaban que las patas pudieran moverse rápido, y sus tres metatarsos estaban combinados entre sí para transferir la fuerza de la pisada desde las patas a las piernas y al resto del cuerpo. La supuesta velocidad del estrutiomimo fue, de hecho, su única defensa contra depredadores, como los dromeosáuridos, como Saurornitholestes y Dromaeosaurus y tiranosáuridos, como el Daspletosaurus y Gorgosaurus, los cuales vivieron en la misma época. Se ha estimado que pudieron ser capaces de correr entre 50 y 80 km/h,[3][43] y que cuando daba 2 zancadas por segundo la longitud máxima de estas pudo llegar a ser de 6 metros.
Struthiomimus fue uno de los primeros coelurosaurios en organizar sus extremidades, músculos y colas como las aves, con un músculo caudofemoralis en la pierna y una cola que actuaba más como timón para cambiar de dirección a alta velocidad. Pero como Struthiomimus poseía una cola más pequeña y ligera, tendría más peso en la parte superior si no tuviera las patas debajo de su centro de equilibrio. Sostener el fémur de manera más horizontal traía sus patas hacia adelante y equilibraba al animal.
La velocidad estimada de Struthiomimus demuestra que pudo ser corredor. Sin embargo, muy probablemente no pudo alcanzar esta capacidad al ritmo que lo hacen las avestruces modernas, debido a que sus largas patas delanteras y su cola corta lo hacían más pesado, y por tanto su velocidad se hubiese limitado en la carrera.
Paleoecología
Los restos fósiles de S. altus solo se conocen de forma definitiva de la Formación Oldman, que data de hace entre 78 y 77 millones de años durante la etapa Campaniense del período Cretácico tardío.[22] Una especie más joven, que aún no ha sido nombrada, que aparentemente difería de S. altus por tener manos más largas y delgadas, se conoce a partir de varios especímenes encontrados en la Formación Horseshoe Canyon y la Formación Lance inferior, entre 69 y 67.5 hace millones de años, maastrichtiense temprano.[22] Struthiomimus habitó en lo que hoy es América del norte, en climas y ambientes muy diferentes a los actuales. El paisaje representado por los depósitos del Parque Provincial del Dinosaurio eran costas subtropicales bajas rodeadas de grandes ríos en las que tanto vida terrestre como marina debió proliferar. El estrutiomimo pudo estar fácilmente adaptado a este ambiente gracias al clima equilibrado y a la abundancia de comida.
Struthiomimus debió ser, por su velocidad, una presa difícil para muchos carnívoros como el Daspletosaurus y el Gorgosaurus. Para otros carnívoros más pequeños, como los dromeosáuridos como el Saurornitolestes y el Dromaeosaurus, la caza les habría resultado más sencilla gracias a su gran rapidez, comparable con la de Struthiomimus. Mientras en las despejadas praderas hubieran migrado manadas de ceratopsianos, como el Chasmosaurus y Styracosaurus y de hadrosáuridos, como el Parasaurolophus y Lambeosaurus. El ambiente donde vivía el Struthiomimus también contenía en sus playas, mares y ríos otros arcosaurios como los pterosaurios, plesiosaurios y mosasaurios.[44] También había numerosas clases de peces, anfibios, reptiles, insectos y crustáceos.
Struthiomimus en la cultura popular
- Representado mediante la técnica go motion por el técnico en efectos especiales Phil Tippett, Struthiomimus aparece robando y devorando huevos de Hadrosaurus en el documental de 1985 Dinosaurios.
- Struthiomimus aparece en la segunda parte de The Land Before Time, la película de dibujos animados The Land Before Time II: The Great Valley Adventure (de Roy Allen Smith, 1994, titulada en Hispanoamérica La tierra antes del tiempo 2: La aventura del gran valle y en España En busca del valle encantado 2: Aventuras en el gran valle). Hay una pareja de ellos como los principales villanos de esta película, en la que tratan de robar un huevo de tiranosaurio encontrado por los protagonistas, a los que también atacan.
- Struthiomimus también hace aparición en la película Dinosaurio (producida en 2000 por Walt Disney Pictures) como un cazador solitario del desierto.
- El grupo de música Fourth Grade Security Risk ha creado una canción titulada The Struthiomimus Strut.
Véase también
Referencias
- P. J. Currie. 1987. Theropods of the Judith River Formation of Dinosaur Provincial Park, Alberta, Canada. In P. J. Currie & E. H. Koster (eds.), Fourth Symposium on Mesozoic Terrestrial Ecosystems. Tyrrell Museum of Paleontology, Drumheller, Alberta 52-60
- Paul, Gregory S. (1988). «Ornithomimus altus». Predatory Dinosaurs of the World. New York: Simon & Schuster. pp. 387–389. ISBN 978-0-671-61946-6.
- Paul, Gregory S. (1988). «Genus Ornithomimus». Predatory Dinosaurs of the World. New York: Simon & Schuster. pp. 384–394. ISBN 978-0-671-61946-6.
- Currie, Philip J. (2005). «Theropods, Including Birds». En Currie, Phillip J.; Koppelhus, Eva, eds. Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Bloomington: Indiana University Press. pp. 367–397. ISBN 978-0-253-34595-0.
- Makovicky, Peter J.; Kobayashi, Yoshitsugu; Currie, Philip J. (2004). «Ornithomimosauria». En Weishampel, David B.; Dodson, Peter; Osmólska, Halszka, eds. The Dinosauria (2nd edición). Berkeley: University of California Press. pp. 137–150. ISBN 978-0-520-24209-8.
- Osborn, Henry Fairfield (1917). «Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus». Bulletin of the American Museum of Natural History 35: 733-771.
- Nicholls, Elizabeth L.; Russell, Anthony P. (1985). «Structure and function of the pectoral girdle and forelimb of Struthiomimus altus (Theropoda: Ornithomimidae)». Palaeontology 28: 643-677.
- Zelenitsky, D. K.; Therrien, F.; Erickson, G. M.; Debuhr, C. L.; Kobayashi, Y.; Eberth, D. A.; Hadfield, F. (2012). «Feathered Non-Avian Dinosaurs from North America Provide Insight into Wing Origins». Science 338 (6106): 510-514. Bibcode:2012Sci...338..510Z. PMID 23112330. doi:10.1126/science.1225376.
- Longrich, N (2008). «A new, large ornithomimid from the Cretaceous Dinosaur Park Formation of Alberta, Canada: Implications for the study of dissociated dinosaur remains». Palaeontology 51 (4): 983-997. doi:10.1111/j.1475-4983.2008.00791.x. Parámetro desconocido
|doi-access=ignorado (ayuda) - Nicholls, E. and Russell, A. Structure and function of the pectoral girdle and forelimb of Struthiomimus altus (Theropoda: Ornithomimidae). Palaeontology (Oxford) 28(4); 643-677 (1985).
- D. A. Russell. 1972. Ostrich dinosaurs from the late Cretaceous of western Canada. Canadian Journal of Earth Sciences 9:375-402
- McFeeters, Bradley; Ryan, Michael J.; Schröder-Adams, Claudia; Cullen, Thomas M. (2016). «A new ornithomimid theropod from the Dinosaur Park Formation of Alberta, Canada». Journal of Vertebrate Paleontology 36 (6): e1221415. doi:10.1080/02724634.2016.1221415.
- Parks, W. Trans. Roy. Soc. Canada 20 (3); 65-70 (1926). Struthiomimus brevetertius, a new species of dinosaur from the Edmonton formation of Alberta.
- Parks, W.A. (1928). «Struthiomimus samueli, a new species of Ornithomimidae from the Belly River Formation of Alberta». University of Toronto Studies, Geology Series 26: 1-24.
- Parks, W.A. (1933). «New species of dinosaurs and turtles from the Upper Cretaceous formations of Alberta». University of Toronto Studies, Geological Series 34: 1-33.
- D. A. Russell and T. P. Chamney. 1967. Notes on the biostratigraphy of dinosaurian and microfossil faunas in the Edmonton Formation (Cretaceous), Alberta. National Museum of Canada Natural History Papers 35:1-22
- D. A. Eberth. 1997. Edmonton Group. In P. J. Currie & K. Padian (ed.), Encyclopedia of Dinosaurs 199-204
- R. Steel. 1970. Part 14. Saurischia. Handbuch der Paläoherpetologie/Encyclopedia of Paleoherpetology. Gustav Fischer Verlag, Stuttgart 1-87
- Osmólska H, Roniewicz E; Barsbold R (1972). «A new dinosaur, Gallimimus bullatus n. gen.,n. sp. (Ornithomimidae) from the Upper Cretaceous of Mongolia». Paleontol. Polonica 27: 103-143. Parámetro desconocido
|name-list-style=ignorado (ayuda); Parámetro desconocido|title-link=ignorado (ayuda) - Makovicky, P. J.; Kobayashi, Y.; Currie, P. J. (2004). «Ornithomimosauria». En Weishampel, D. B.; Dodson, P.; Osmólska, H., eds. The Dinosauria (2nd edición). Berkeley: University of California Press. pp. 137–150. ISBN 9780520242098. doi:10.1525/california/9780520242098.003.0008.
- Glut, D., 1997, Dinosaurs - The Encyclopedia. McFarland Press, Jefferson, NC. 1076 pp
- Claessens, L.; Loewen, Mark A. (2015). «A redescription of Ornithomimus velox Marsh, 1890 (Dinosauria, Theropoda)». Journal of Vertebrate Paleontology 36: e1034593. doi:10.1080/02724634.2015.1034593.
- Marsh, O.C. (1892). «Notice of new reptiles from the Laramie Formation». American Journal of Science. Series 3 43 (257): 449-453. Bibcode:1892AmJS...43..449M. doi:10.2475/ajs.s3-43.257.449.
- Farlow, J.O., 2001, "Acrocanthosaurus and the maker of Comanchean large-theropod footprints", In: Tanke, Carpenter, Skrepnick and Currie (eds). Mesozoic Vertebrate Life: New Research Inspired by the Paleontology of Philip J. Currie. pp. 408-427
- Aaron, J.; van der Reest, Alexander P. Wolfe; Currie, Philip J. (2016). «[2015] A densely feathered ornithomimid (Dinosauria: Theropoda) from the Upper Cretaceous Dinosaur Park Formation, Alberta, Canada». Cretaceous Research 58: 108-117. doi:10.1016/j.cretres.2015.10.004.
- O. C. Marsh. 1890. Additional characters of the Ceratopsidae, with notice of new Cretaceous dinosaurs. American Journal of Science 39:418-426
- O. C. Marsh. 1895. On the affinities and classification of the dinosaurian reptiles. American Journal of Science.
- O. C. Marsh. 1896. The dinosaurs of North America. United States Geological Survey, 16th Annual Report, 1894-95 55:133-244
- G. Baur. 1891. Remarks on the reptiles generally called Dinosauria. The American Naturalist 25(293):434-454
- D. A. Russell and Z.-M. Dong. 1993. The affinities of a new theropod from the Alxa Desert, Inner Mongolia, People's Republic of China. Canadian Journal of Earth Sciences 30(10-11):2107-2127
- J. A. Gauthier and K. Padian. 1985. Phylogenetic, functional, and aerodynamic analyses of the origin of birds and their flight. In M. K. Hecht, J. H. Ostrom, G. Viohl, and P. Wellnhofer (eds.), The Beginnings of Birds: Proceedings of the International Archaeopteryx Conference, Eichstätt 1984. Freunde des Jura-Museums Eichstätt, Eichstätt 185-197
- F. E. Novas. 1992. La evolución de los dinosaurios carnívoros [The evolution of carnivorous dinosaurs]. In J. L. Sanz and A. D. Buscalioni (eds.), Los Dinosaurios y Su Entorno Biótico: Actas del Segundo Curso de Paleontología in Cuenca. Instituto "Juan Valdez", Cuenca, Argentina 126-163
- P. C. Sereno, J. A. Wilson, H. C. E. Larsson, D. B. Dutheil, and H.-D. Sues. 1994. Early Cretaceous dinosaurs from the Sahara. Science 266(5183):267-271
- P. J. Makovicky, Y. Kobayashi, and P. J. Currie. 2004. Ornithomimosauria. In D. B. Weishampel, P. Dodson, & H. Osmólska (eds.), The Dinosauria (second edition). University of California Press, Berkeley 137-150
- R. Barsbold. 1976. K evolyutsii i sistematike pozdnemezozoyskikh khishchnykh dinozavrov [The evolution and systematics of late Mesozoic carnivorous dinosaurs]. In N. N. Kramarenko, B. Luvsandansan, Y. I. Voronin, R. Barsbold, A. K. Rozhdestvensky, B. A. Trofimov & V. Y. Reshetov (eds.), Paleontology and Biostratigraphy of Mongolia. The Joint Soviet-Mongolian Paleontological Expedition, Transactions 3:68-75
- P. C. Sereno. 1998. A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 210(1):41-83
- Turner, A.H., Pol, D., Clarke, J.A., Erickson, G.M., and Norell, M. (2007). "Supporting online material for: A basal dromaeosaurid and size evolution preceding avian flight". Science, 317: 1378-1381. doi:10.1126/science.1144066 (supplement)
- Sereno, P. C. 2005. Stem Archosauria—TaxonSearch Archivado el 25 de octubre de 2014 en Wayback Machine. [version 1.0, 2005 November 7]
- Xu, L.; Kobayashi, Y.; Lü, J.; Lee, Y. N.; Liu, Y.; Tanaka, K.; Zhang, X.; Jia, S.; Zhang, J. (2011). «A new ornithomimid dinosaur with North American affinities from the Late Cretaceous Qiupa Formation in Henan Province of China». Cretaceous Research 32 (2): 213. doi:10.1016/j.cretres.2010.12.004.
- Rothschild, B., Tanke, D. H., and Ford, T. L., 2001, Theropod stress fractures and tendon avulsions as a clue to activity: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 331-336.
- Makovicky PJ, Kobayashi Y, Currie PJ (2004). «Ornithomimosauria». En Weishampel DB, Osmólska H, Dodson P, ed. The Dinosauria (2nd Edition). University of California Press. pp. 149. ISBN 0-520-24209-2.
- Osmólska H, Roniewicz E & Barsbold R (1972). «A new dinosaur, Gallimimus bullatus n. gen.,n. sp. (Ornithomimidae) from the Upper Cretaceous of Mongolia». Paleontol. Polonica 27: 103-143.
- Paul, G.S. 1988. Predatory Dinosaurs of the World. New York: Simon & Schuster.
- P. J. Currie. 2005. History of research. In P. J. Currie & E. B. Koppelhus. Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press, Bloomington 3-33
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikispecies tiene un artículo sobre Struthiomimus altus.
Wikispecies tiene un artículo sobre Struthiomimus altus. Wikimedia Commons alberga una categoría multimedia sobre Struthiomimus altus.
Wikimedia Commons alberga una categoría multimedia sobre Struthiomimus altus.- Descripción sencilla del Struthiomimus en Duiops
- Ilustraciones del Struthiomimus en The Natural History Museum's Dino Directory (en inglés)
| Control de autoridades |
|
|---|
 Datos: Q310535
Datos: Q310535- Multimedia: Struthiomimus / Q310535
- Especies: Struthiomimus