Herrerasaurus ischigualastensis
Herrerasaurus ischigualastensis (lat. "lagarto de Herrera de Ischigualasto") es la única especie conocida del género extinto Herrerasaurus de dinosaurio saurisquio herrerasáurido, que vivió en el Carniense, a principios del período Triásico superior, hace aproximadamente 231,4 millones de años, en lo que es hoy Sudamérica. Sus restos se encontraron en el Valle de la Luna del noroeste de Argentina. La especie tipo, Herrerasaurus ischigualastensis, fue descrita por Osvaldo Alfredo Reig en 1963[1] y es la única especie asignada al género.
| Herrerasaurus ischigualastensis | ||
|---|---|---|
| Rango temporal: 231,4 Ma - 228 Ma Triásico Superior | ||
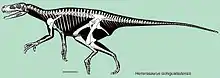
 Esqueleto montado, Museo Senckenberg de Historia Natural | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Familia: | Herrerasauridae | |
| Género: |
Herrerasaurus Reig, 1963 | |
| Especie: |
H. ischigualastensis Reig, 1963 | |
| Sinonimia | ||
| ||
Durante muchos años, la clasificación de Herrerasaurus no fue clara, ya que la descripción del animal se basaba en unos pocos restos. Esto conllevó la emisión de diversas hipótesis taxonómicas encuadrándolos como un terópodo basal, un sauropodomorfo basal, un saurisquio basal, o inclusive un no dinosaurio. Sin embargo, con el descubrimiento de un esqueleto completo con la cabeza,[2][3] Herrerasaurus fue clasificado como un terópodo temprano o un saurisquio temprano en por lo menos cinco exámenes recientes de la evolución de los terópodos. Este reptil bípedo de tamaño mediano es un miembro de Herrerasauridae, un grupo de animales similares que estaban entre los dinosaurios de la más temprana radiación de los dinosaurios.
Descripción
Herrerasaurus es uno de los dinosaurios más primitivos de los que se tengan registros, era un carnívoro bípedo ligeramente construido con una cola larga y una cabeza relativamente pequeña. Presenta un tamaño medio, con el fémur de unos 50 centímetros de longitud, y un largo total de entre 3 y 6 metros,[3] y una altura a las caderas de 1,1 metros,[4] llegando a pesar entre 210 a 350 kilogramos.[4] El mayor espécimen en un principio fue considerado un género separado, Frenguellisaurus, cuyo cráneo medía unos 56 centímetros de largo.[4] Posee una mezcla de características primitivas y derivadas que confundieron a los primeros estudiosos de este saurisquio acerca de su posición sistemática y sus relaciones filogenéticas. El cuello del Herrerasaurus no muestra signos de elongación. Es bastante corto y fuerte. En las vértebras dorsales están presentes algunos rasgos derivados interesantes, por ejemplo los centros vertebrales son altos y cortos, y la morfología derivada del arco neural, con espinas más bien cuadradas, con un engrosamiento dorsal. Estos rasgos indican que Herrerasaurus tenía características derivadas especiales y únicas en la columna vertebral. El hueso sacro es primitivo por las pocas vértebras que lo integran, solo tres. La reducción del tamaño de la cintura pectoral, muestra la lámina escapular angosta y corta, pero con un área coracoide-acromial expandida.

Herrerasaurus muestra rasgos que se encuentran en diferentes grupos de dinosaurios, y varios rasgos que se encuentran en los arcosaurios no dinosaurios. Aunque comparte la mayoría de las características de los dinosaurios, existen algunas diferencias, particularmente en la forma de los huesos de la cadera y las piernas. Su pelvis es como la de los dinosaurios saurisquios, pero tiene un acetábulo óseo, donde el fémur se encuentra con la pelvis, que estaba solo parcialmente abierto. El ilion, el hueso principal de la cadera, está soportado por solo dos vértebras sacras, un rasgo basal.[5] Sin embargo, el pubis apunta hacia atrás, un rasgo derivado como se ve en los dromeosáuridos y las aves. Además, el final del pubis tiene una forma de bota, como las de los aveterópodos y el centro vertebral tiene una forma de reloj de arena como el que se encuentra en el Allosaurus.[4]
Cráneo

Los adultos tenían cráneos de hasta 56 centímetros de largo y tenían hasta 6 metros de longitud total[3] y 350 kilogramos de peso.[4] Las muestras más pequeñas tenían la mitad del tamaño, con cráneos de solo 30 centímetros de largo.[5] El cráneo es estrecho y largo y carecía de casi todas las especializaciones que caracterizaron a los dinosaurios posteriores,[6] no tan diferente al de los arcosaurios anteriores como Euparkeria. Está provisto de una región preorbital, un hocico bien desarrollado, que representa más de la mitad del largo del cráneo. Posee cinco pares de aberturas en el cráneo, un par son las oculares y otra las abertura nasales. Entre estas hay una fenestra antorbital bien desarrollada y dos aberturas pequeñas, de un centímetro de largo con forma de ojal llamadas fenestra promaxilar.[7] detrás de los ojos hay una gran fenestra infratemporal. Estos huecos ayudan a reducir el peso del cráneo. La abertura preorbital es proporcionalmente grande y la proyección dorsal del maxilar muy ancha y fuerte, con un largo contacto con el nasal. El premaxilar es también fuerte con un contacto extenso pero no rígido con el maxilar. La abertura nasal externa es angosta.

Herrerasaurus poseía una articulación flexible entre los huesos dentarios y postdentarios, esto permitió que el animal resbalara su quijada inferior hacia adelante y hacia atrás y dar una mordedura de compresión.[6] La especialización craneal es inusual entre los dinosaurios pero se ha desarrollado independientemente en algunos lagartos.[8] La parte posterior de la quijada inferior también tenía ventanas. Las mandíbulas fueron equipadas de los dientes serrados grandes para morder y comer carne, y el cuello era delgado y flexible.[6][9] Los dientes superiores forman una larga hilera, de unas 20 piezas. Aquellos del premaxilar y de la porción anterior del maxilar son de tamaño medio, pero siguiendo hacia atrás hay de 4 a 5 dientes grandes y fuertes caniniformes, justo debajo de la fuerte proyección dorsal del maxilar. La hilera superior de dientes es completada por unas 10 piezas mucho más pequeñas que se extienden hasta debajo de la órbita. A raíz de estos caracteres se puede inferir que la acción de atrapar y matar a la presa era trabajo de los dientes anteriores.
La región temporal del cráneo es primitiva, con una cresta parietal larga y baja, una gran abertura supratemporal y una abertura infratemporal ancha y alta, con el cuadrado casi vertical.
Extremidades

Herrerasaurus era completamente bípedo. Tenía miembros posteriores fuertes con muslos cortos y pies bastante largos, lo que indica que probablemente era un corredor veloz. El pie tenía cinco dedos, pero solo los tres medios, los dígitos II, III y IV, soportaban peso. Los dedos exteriores, los I y V, eran pequeños. El primer dedo tenía una pequeña garra. La cola, parcialmente rígida por las proyecciones vertebrales superpuestas, equilibró el cuerpo y también fue una adaptación para la velocidad.[5] Las extremidades anteriores de Herrerasaurus tenían menos de la mitad de la longitud de sus extremidades traseras. La parte superior del brazo y el antebrazo eran bastante cortos, mientras que la mano estaba alargada. Los primeros dos dedos y el pulgar terminaron en garras curvas y afiladas para agarrar la presa. Los dígitos cuarto y quinto eran pequeños y sin garras.[3][10]
El tamaño relativo de los miembros anteriores sugieren que era de hábitos mayormente bípedos. EL antebrazo y el brazo son cortos, con unas manos largas. Esta poseía tres dedos alargados, el tercero es el más largo con fuertes garras que se cerraban sobre la palma permitiéndole tomar a sus presas, los dedos IV y V son muy pequeños y sin garras. Lo cual represente una forma primitiva en algunos dinosaurios terópodos como los Herrerasáuridos y algunos Abelisaurios.[3][10] Herrerasaurus era bípedo. Tenía miembros traseros fuertes con un corto muslo y los pies algo largos, indicando este animal era un probable corredor rápido. La cola le daba equilibrio, se atiesaba parcialmente traslapando procesos vertebrales, también indica una adaptación para la velocidad.[5] En la extremidad posterior, los metatarsos muestran un buen grado de superposición. Los metatarsos de los dedos I y a pesar de estar bien desarrollados, son muy reducidos en grosor y longitud respecto a los metatarsos II, III y IV. Esto probablemente significa que sus pies eran funcionalmente tridáctilos.
Características anatómicas distintivas
El diagnóstico es una declaración de las características anatómicas de un organismo (o grupo) que colectivamente lo distinguen de todos los demás organismos. Algunas, pero no todas de las características en un diagnóstico también son autapomorfías. Una autapomorfía es una característica anatómica distintiva que es única de un organismo o grupo determinado. De acuerdo con Novas en 1993, Herrerasaurus puede distinguirse sobre la base de las siguientes características:[11] La presencia de una fenestra entre el premaxilar y el maxilar, y la parte dorsal de la fenestra laterotemporal es menos de un tercio del ancho de la parte ventral de la misma. La presencia de una cresta en la superficie lateral del hueso yugal, y una fosa supratemporal muy perforada que se extiende a través del proceso postorbital medio. El proceso escamosal subcuadrado ventral tiene una depresión lateral, y el hueso cuadratoyugal se superpone a la cara posterodorsal del hueso cuadrado. El proceso pterigoideo del cuadrado tiene un margen ventral curvo en forma de depresión, y cuenta con un proceso delgado en forma de canal del hueso dentario en su parte posterodorsal. El hueso surangular tiene un proceso anterior bifurcado para la articulación con el proceso posterodorsal del dentario. La tuberosidad interna del húmero se proyecta proximalmente y está separada de la cabeza del húmero por una profunda muesca, también presente en los celofisoideos. Posee manos alargadas, las cuales tienen un tamaño equivalente al 60% del húmero y el radio, y el entepocóndilo del húmero tiene forma de cresta con depresiones anteriores y posteriores. El borde posterior del pedúnculo del ilion forma un ángulo recto con el borde dorsal del eje del isquion.
De acuerdo con Paul Sereno 1993, Herrerasaurus puede ser distinguido con las siguientes características, todas las cuales son desconocidas en otros herrerasáuridos:[12] Una incisión circular presente en el ectepicóndilo del húmero, un rasgo también presente en Saturnalia Un cóndilo cubital en el húmero en forma de silla de montar, y la superficie articular para el hueso ulnar en el cúbito es convexa. La superficie articular del ulnar es más pequeña que la del cúbito, un rasgo desconocido en Staurikosaurus y Sanjuansaurus. El central se sitúa distalmente con respecto al radial. Un amplio proceso subnarial en el premaxilar, y una ancha depresión supratemporal, señalado por Sereno y Novas, 1993.[13] El tubérculo basal y el cóndilo occipital son de anchura casi igual señalado por Sereno y Novas, 1993.[13]
Descubrimiento e investigación

El primer espécimen de Herrerasaurus fue descubierto en 1959, en el valle de Ischigualasto, por una comisión paleontológica de la Universidad Nacional de Tucumán, encabezada por el Dr.Osvaldo Reig, quien lo estudió en 1963.[1] Victorino Herrera, un pastor local, un miembro argentino de la mencionada expedición paleontológica, fue quien encontró los primeros fósiles de esta especie, a quien O. Reig le dedicó el nombre genérico del dinosaurio. Luego fue estudiado por otros investigadores argentinos y extranjeros. Estas rocas, en las que más adelante se encontró el Eoraptor,[14] es parte de la Formación Ischigualasto y data del Ladiniense a Carniense temprano en el Triásico Superior.[15] O. Reig nombró un segundo dinosaurio de estas rocas en la misma publicación que el Herrerasaurus;[1] este dinosaurio, Ischisaurus cattoi, es hoy considerado sinónimo más moderno y un ejemplar juvenil de Herrerasaurus.[9] Otros dos esqueletos parciales, con material craneal, fueron llamados Frenguellisaurus ischigualastensis por Fernando Novas en 1986,[16] pero esta especie ahora es también probablemente un sinónimo.[9] Frenguellisaurus ischigualastensis fue descubierto en 1975, y fue descrito por Novas en 1986, quien lo consideró un saurisquio primitivo, y posiblemente un terópodo. Novas en 1992 y Sereno y Novas en 1992 examinaron los restos de Frenguellisaurus y los encontraron remitibles a Herrerasaurus.[1] Ischisaurus cattoi fue descubierto en 1960 y descrito por Reig en 1963. Novas en 1992 y Sereno y Novas en 1992 revisaron sus restos y los encontraron como referentes a Herrerasaurus.[9]
O. Reig consideró a Herrerasaurus como un ejemplo temprano de un carnosaurio,[1] pero esto fue sujeto de muchos debates por los siguientes treinta años, y el género fue varias veces clasificado durante esos años. En 1970, R. Steel clasificó a Herrerasaurus como un prosaurópodo.[17] En 1972, Peter Galton lo clasificó como indestinguible más allá de Saurischia.[18] Posteriormente usando el método cladístico de análisis, algunos investigadores colocaron a Herrerasaurus y Staurikosaurus en la base del árbol del dinosaurio antes de la separación entre los ornitisquios y los saurisquios.[19][20][21][22] Otros investigadores consideran los restos como pertenecientes a un no dinosaurio.[23]
Un cráneo completo de Herrerasaurus no fue encontrado hasta 1988, por un equipo de paleontólogos liderados por Paul Sereno.[3][24] Basándose en los nuevos fósiles, autores tales como Thomas Holtz[25] y José Fernando Bonaparte[26] Herrerasaurus es clasificado en la base del árbol de los saurisquios antes de la divergencia entre los sauropodomórfos y los terópodos. Sin embargo, P. Sereno esta a favor de clasificar a Herrerasaurus y Herrerasauridae, su familia como terópodos primitivos.

Estas dos clasificaciones se han convertido en las más persistentes, con Rauhut en 2003[27] y Bittencourt y Kellner en 2004[28] favoreciendo la hipótesis del terópodo temprano, y Max Langer en 2004,[5] Langer y Benton en 2006,[29] y Randall Irmis y sus coautores en 2007[30] favoreciendo la hipótesis del saurisquio básico. Si Herrerasaurus fuera de hecho un terópodo, indicaría que los terópodos, los sauropodomórfos, y los ornitisquios divergieron incluso antes que los herrerasáuridos, previamente al Carniano medio, edad de Ischigualasto, y que " los tres linajes desarrollaron independientemente varias características dinosaurianas, tales como una articulación de tobillo más avanzada o un acetábulo abierto ".[31]

Esta visión es apoyada más a fondo por los expedientes de huellas fósiles que demuestran las grandes pisadas del tridáctilas que se pueden atribuir solamente a un dinosaurio terópodo, fechando en el Ladiniense, Triásico Medio de la Formación Los Rastros en la Argentina y que preceden a Herrerasaurus por 3 a 5 millones de años.[32][33]
La importancia de Herrerasaurus y Eoraptor está dada por el hecho de que sus restos apoyan directamente la idea de la monofíla de los dinosaurios, lo que es lo mismo decir que todos los dinosaurios tienen un ancestro común. La monofília de los dinosaurios fue explícitamente propuesta por Robert Bakker en los años de 1970,[34][35] y nueve características craneales y quince postcraneales sinapomorfias, características comunes derivadas de un mismo ancestro, han sido mencionadas. Sin embargo, un extensivo estudio de Herrerasaurus por P. Sereno indican que sólo una craneal y siete postcraneales sinapomorfias en la lista original de R. Bakker, son actualmente consideradas, agregando nuevas a la lista.[3]
Clasificación
Herrerasaurus le da el nombre a toda la familia, Herrerasauridae, de finales del Triásico, junto con sus parientes cercanos se ve enredados en un árbol evolucionario de los dinosaurios no muy claro. Existiendo la posibilidad de que sea un primitivo terópodo o inclusive un saurisquio basal y aun preceder la separación entre los saurisquios y los ornitisquios.[22] Otros miembros del clado incluyen al Eoraptor de la misma formación geológica que el Herrerasaurus, Staurikosaurus de la Formación Santa María del sur del Brasil,[36] Chindesaurus de la Formación Chinle de Arizona,[37] y posiblemente Caseosaurus de la Formación Dockum de Texas,[38] aunque las relaciones de estos animales no se entiendan completamente, y no todos los paleontólogos opinen igual. Otros terópodos básicos posibles, Alwalkeria del último Triásico Formación Maleri de la India,[39] y Teyuwasu, conocidos por restos muy fragmentarios del Triásico superior de Brasil, pudo haber estado relacionado.[40] Novas en 1992 definió el grupo como Herrerasaurus, Staurikosaurus, su ancestro común más cercano.[21] Sereno en 1998 define el grupo como el clado más inclusivo que contiene a H. ischigualastensis pero no a Passer domesticus.[41] Langer en 2004 crea un taxón de nivel más elevado, el infraorden Herrerasauria.[5]

No está claro si Herrerasaurus y sus parientes cercanos se encuentran en los primeros dinosaurios del árbol evolutivo. La mayoría de los análisis del siglo 21 se ha encontrado que son terópodos basales, aunque pueden, de hecho representen saurisquios basales, o incluso ser no dinosaurios, anterior a los Saurischia - Ornithischia. La mayoría de los análisis, como los que por Sterling Nesbitt y sus colegas en 2009 y 2011, han encontrado que Herrerasaurus y sus familiares pueden ser terópodos muy basales. Otros, como Ezcurra 2010, han encontrado que son basales del clado Saurischia, es decir, más cerca de la base del árbol de Saurischia de los verdaderos terópodos o sauropodomorfos, pero no miembros de ambos grupos. La situación se complica aún más por la incertidumbre en la correlación de las edades de finales del Triásico.
Herrerasaurus fue originalmente considerado como un género dentro de Carnosauria, que luego incluía formas similares a Megalosaurus y Antrodemus, este último es equivalente a Allosaurus,[42] aunque Herrerasaurus vivió muchos millones de años antes de ellos y conservó múltiples características primitivas . Esta clasificación carnosauriana fue modificada por Rozhdestvensky y Tatarinov en 1964, quienes clasificaron a Herrerasaurus dentro de la familia Gryponichidae dentro de Carnosauria. El mismo año, Walker publicó una opinión diferente de que Herrerasaurus, en cambio, estaba aliado con Plateosauridae, aunque difería en poseer una bota púbica. Walker también propuso que Herrerasaurus podría estar más cerca de Poposaurus, ahora considerado un pseudosuquio[43] y el terópodo anónimo del Grupo Dockum de Texas , ahora asignado al rauisuquio Postosuchus.[44] En 1985, Charig notó que Herrerasaurus era de clasificación incierta, mostrando similitudes tanto con los prosauropodos como con los carnosaurios. Romer en 1966, simplemente observó que Herrerasaurus era un prosauropodo posiblemente dentro de Plateosauridae. En la descripción de Staurikosaurus, Colbert notó que había muchas similitudes entre su taxón y Herrerasaurus, pero los clasificó en familias separadas, con Herrerasaurus en Teratosauridae. En 1970, Bonaparte también propuso similitudes entre Herrerasaurus y Staurikosaurus y al clasificarlos claramente en Saurischia, afirmó que parecían no estar en una familia actual. Benedetto lo apoyó aún más en 1973, quien nombró para los taxones la nueva familia Herrerasauridae, que clasificó como saurisquios, posiblemente dentro de Theropoda pero no en Sauropodomorpha..[45] Sin embargo, en 1977 Galton propuso que Herrerasauridae solo incluyera a Herrerasaurus , y encontró que era Saurischia incertae sedis.[46]

Propuesto en 1987 por Brinkman y Sues, Herrerasaurus a veces se ha considerado basal para Ornithischia y Saurischia, aunque Brinkmann y Sues todavía consideraban que estaba dentro de Dinosauria. Apoyaron esto sobre la base de que Herrerasaurus tiene un gran dígito V en el pie, y tiene una pared medial bien desarrollada en el acetábulo. Brinkmann y Sues consideraron que Staurikosaurus y Herrerasaurus no formaban un verdadero grupo llamado Herrerasauridae, y que, en cambio, eran formas más primitivas. Además, consideraron que los caracteres utilizados por Benedetto no eran válidos, en lugar de representar solo el estado plesiomórfico que se encontró en ambos taxones.[47] Esto fue rebatido en 1992 por Novas, quien declaró muchas sinapomorfias derivadasde Herrerasauridae, como una bota púbica distinta, pero aún las clasificó como basales de Ornithischia y Saurischia. Novas definió a la familia como el antepasado menos común de Herrerasaurus y Staurikosaurus y todos sus descendientes.[48] Una definición diferente de la propuesta por Sereno en 1998 que incluye a Herrerasaurus pero no de Passer domesticus y sigue más de cerca la inclusión original propuesta por Benedetto.[49] Otro grupo, Herrerasauria fue nombrado por Galton en 1985, y se definió como Herrerasaurus pero no Liliensternus o Plateosaurus por Langer en 2004, quienes usaron la definición basada en nodos para Herrerasauridae.[50] En una revisión de Dinosauria basal, Padian y May en 1993 discutieron la definición del clado y lo redefinieron como el último ancestro común de Triceratops y aves. También discutieron lo que esta definición haría con los taxones más basales, como Herrerasauridae y Eoraptor . Padian y May consideraron que, dado que tanto Herrerasauridae como Eoraptor carecen de muchas características diagnósticas de Saurischia u Ornithischia, no podían considerarse dentro de Dinosauria.[51]
Un estudio posterior realizado en 1994 por Novas, en cambio, clasificó a Herrerasaurus dentro de Dinosauria, y apoyó firmemente su posición dentro de Saurischia, así como una sinapomorfia compartida con Theropoda. Novas descubrió que las características primitivas de carecer de fosa corta y de tener solo dos vértebras sacras eran simplemente reversiones encontradas en el género.[52] En 1996, Novas fue más allá al apoyar una posición de terópodo para Herrerasaurus con un análisis filogenético, que lo colocó más cerca de Neotheropoda que de Eoraptor o Sauropodomorpha.[53] Langer en 2004 mencionó que esta hipótesis fue ampliamente aceptada, pero que los autores posteriores, en cambio, prefirieron ubicar a Herrerasaurus así como Eoraptor basal para Theropoda y Sauropodomorpha, un clado llamado Eusaurischia. Langer en 2004 realizó un análisis filogenético y descubrió que era mucho más probable que Herrerasaurus fuera un saurisquío basal, que un terópodo o un no dinosaurio.[50] La propuesta de Langer fue apoyada por múltiples estudios hasta el descubrimiento de Tawa , cuando Nesbitt et al. realizó un análisis más inclusivo, y el cladograma resultante colocó Herrerasauridae basal a Eoraptor, pero más cerca de Dilophosaurus que de Sauropodomorpha.[54][55] A diferencia de Nesbitt, Ezcurra en 2010 realizó un análisis filogenético para colocar su nuevo taxón Chromogisaurus y descubrió que Herrerasauridae era basal para Eusaurischia.[56]
En 2010, Alcocer y Martínez describieron un nuevo taxón de herrerasáurido, Sanjuansaurus. Se podría distinguir de Herrerasaurus por múltiples características. En el análisis filogenético, Herrerasaurus , Sanjuansaurus y Staurikosaurus estaban en una politomía y Herrerasauridae era el grupo más primitivo de saurisquios, fuera de Eusaurischia, Eoraptor y Guaibasaurus.[57] En 2011, Martínez et al. describió a Eodromaeus, un terópodo basal de la misma formación que Herrerasaurus . En un análisis filogenético, Eoraptor se colocó dentro de Sauropodomorpha, Herrerasauridae se colocó como los terópodos más basales y Eodromaeus se colocó como el más basal siguiente.[58] Un análisis más reciente, por Bittencourt et al. de 2014, colocaron a Herrerasauridae en una politomía con Theropoda y Sauropodomorpha, con Eoraptor también en una posición no resuelta.[59]
En marzo de 2017, un artículo publicado en la revista Nature por Matthew Baron, David Norman y Paul Barrett, incluyó un análisis en el cual los dinosaurios terópodos, estaban más cercanamente relacionados con los dinosaurios ornitisquios que a los Sauropodomorpha, el grupo al que pertenecen los dinosaurios saurópodos, que sería un clado hermano de Herrerasauridae dentro de Saurischia.[60]
Géneros relacionados
Otros miembros del clado pueden incluir a Eoraptor de la misma Formación Ischigualasto de Argentina en que se halló a Herrerasaurus, Staurikosaurus de la Formación Santa María del sur de Brasil,[61] Chindesaurus del Bosque Petrificado de la Formación Chinle de Arizona,[62] y posiblemente Caseosaurus de la Formación Tecovas del Grupo Dockum en Texas, en Estados Unidos[63] aunque las relaciones de estos animales no son bien entendidas, y no todos los paleontólogos están de acuerdo. Otros terópodos basales posibles, Alwalkeria del Triásico Superior de la Formación Maleri de la India,[64] y Teyuwasu, conocido a partir de restos muy fragmentarios del Triásico Superior de Brasil, podrían estar relacionados.[65] Gregory Paul en 1988 sugirió que se había señalado incorrectamente que Staurikosaurus pricei era un juvenil de Herrerasaurus. Esta afirmación fue refutada cuando los huesos pélvicos de un ejemplar joven de Herrerasaurus fueron descubiertos, que al ser examinados no revelaron ningún parecido particular a los de la pelvis de Staurikosaurus.[4]
Filogenia
Su posición es muy controvertida; por un lado Herrerasauridae se define como el clado más inclusivo que contiene a Herrerasaurus ischigualastensis (Reig, 1963) pero no a Passer domesticus (Linneo, 1758). Son todos los terópodos más cercanos al Herrerasaurus que a las aves verdaderas. El cladograma que sigue se basa en uno propuesto por Martín Ezcurra en 2010. En esta revisión, Herrerasaurus es un saurisquio primitivo, pero no un terópodo.[66]

| Dinosauria |
| ||||||||||||||||||||||||||||||||||||
El cladograma que sigue se basa en uno propuesto por Bittencourt et al. en 2014.[59]
| Dinosauria |
| |||||||||||||||||||||||||||||||||||||||
El cladograma que sigue se basa en la propuesta alternativa realizada por Matthew Baron, David Norman y Paul Barrett en 2017.[60]
| Dinosauromorpha |
| ||||||||||||||||||||||||||||||
Paleobiología
Los dientes de Herrerasaurus indican que eran carnívoros, su tamaño indica que habrían cazado pequeños y medianos animales. Se pudieron haber alimentado de otros dinosaurios, tales como el herbívoro Pisanosaurus. Sin embargo, ya que Herrerasaurus vivió durante una era cuando otros dinosaurios eran infrecuentes, habría incluido entre sus presas los más abundantes rincosáuridos y sinápsidos herbívoros.[67] A su vez los herrerasaurios en si debió haber sido presa de los gigantes rauisuquianos como Saurosuchus, habiéndose encontrado heridas de dientes en su cráneo.[6]
Se han descubierto coprolitos, estiércol fosilizado, que contienen pequeños huesos pero ningún rastro de fragmentos de plantas en la formación de Ischigualasto, asignándose a Herrerasaurus de acuerdo con su abundancia fósil. El análisis mineralógico y químico de estos coprolitos indica que el animal carnívoro tenía la capacidad de digerir los huesos.[68]
Las comparaciones entre los anillos esclerales de Herrerasaurus y las aves y reptiles modernos sugieren que puede haber sido catemeral, activo durante todo el día a intervalos cortos.[69]

Aunque los herrerasaurios compartían la forma del cuerpo de los dinosaurios carnívoros grandes, vivió cerca de hace 230 millones de años, una época en que los dinosaurios eran pequeños e insignificantes. Era la época de reptiles no-dinosaurianos, no de dinosaurios, y de un momento crucial en la ecología de la tierra. La fauna vertebrada de la Formación Ischigualasto y la levemente joven Formación Los Colorados consistidas principalmente en una variedad arcosaurios crurotarsianos y sinápsidos. [70][71] Para ese momento, en Ischigualasto los dinosaurios constituían solo el 6% del total de los fósiles.[72] Para finales del periodo Triásico, los dinosaurios se habían convertido en los animales dominates con los sinápsidos y crurotarsianos perdiendo diversidad.[73]
Paleopatología
En un estudio de 2001 llevado a cabo por Bruce Rothschild y otros paleontólogos, se examinaron 12 huesos de manos y 20 huesos de pies asignados a Herrerasaurus para buscar indicios de fracturas por estrés, pero no se halló ninguno.[74]
PVSJ 407, un ejemplar de Herrerasaurus ischigualastensis, tenía una incisión en un hueso craneal atribuida por Paul Sereno y Fernando Novas a una mordida. Dos agujeros más aparecen en el esplenial. Las áreas alrededor de los agujeros están hinchadas y son porosas, lo que sugiere que las heridas sufrieron una infección no letal de corta duración. Tomando en cuenta el tamaño y el ángulo de las heridas, es probable que fueran infligidas durante una lucha con otro Herrerasaurus.[75]
Paleoecología
Procedencia y ocurrencia
El holotipo de Herrerasaurus (PVL 2566) fue descubierto en la Cancha de Bochas Miembro de la Formación Ischigualasto , en San Juan, Argentina. Fue recogido en 1961 por Victorino Herrera, en los sedimentos que fueron depositados en la época del Carniano del período Triásico, entre hace aproximadamente 235 hasta 221 millones años. Con los años, en la Formación Ischigualasto se han descubierto otros fósiles que han sido referidos a Herrerasaurus. En 1958, AS Romer descubrió el espécimen MCZ 7063, originalmente asignado a Staurikosaurus que databa del Carniense. Los especímenes de Herrerasaurus PVL 2045 y MLP (4) 61 fueron recolectados en 1959 y 1960, respectivamente, en los sedimentos que fueron depositados en la época del Noriense del Triásico, hace aproximadamente 228 a 208 millones de años. En 1960, Scaglia recogió los especímenes MACN 18.060, originalmente el holotipo de Ischisaurus cattoi, en los sedimentos depositados en el Carniense. En 1961, Scaglia recogió un ejemplar de Herrerasaurus PVL 2558, en los lechos del Carniense de esta formación. En 1990, se hallaron en el Miembro Cancha de Bochas más ejemplares de Herrerasaurus, también de sus lechos del Carniano. El espécimen PVSJ 53, originalmente el holotipo de Frenguellisaurus fue recogido por Gargiulo y Oñate en 1975, en los sedimentos que fueron depositados en el Noriense.
Paleoambiente
Los estudios han sugerido que el paleoambiente de la Formación Ischigualasto era un terreno de aluvión volcánico activo cubierto por los bosques y conforme a precipitaciones estacionales fuertes. La vegetación consistía en helechos (Cladophlebis), equisetos, y coníferas gigantes (Protojuniperoxylon).[71] Las plantas formaban un bosque ribereño de tierras altas.[3] Los restos de Herrerasaurus parecen haber sido los más comunes entre los carnívoros de la Formación Ischigualasto.[15] Vivió en las selvas del Triásico Superior de Sudamérica junto a otro dinosaurio primitivo, Eoraptor de un metro de largo, así como Saurosuchus, un gigantesco rauisuquio terrestre[76] (carnívoros cuadrúpedos con cráneos semejantes a los de los terópodos), los más o menos similares, pero más pequeños Venaticosuchus, Ornithosuchus, y los sinápsidos depredadores Chiniquodontidae.[15] Los herbívoros eran mucho más abundantes que los carnívoros y estuvieron representados por rincosaurios como Hyperodapedon (un reptil con pico); los aetosaurios (reptiles acorazados); los dicinodontes kannemeyéridos (robustos animales cuadrúpedos con un picos) como Ischigualastia y los terápsidos Traversodontidae (algo similares en forma general a los dicinodontes, pero que carecían de picos), tales como Exaeretodon. Estos herbívoros no dinosaurios eran mucho más abundantes que los dinosaurios ornitisquios tempranos como Pisanosaurus.[77]
Fauna y hábitat
Aunque Herrerasaurus comparte la forma del cuerpo de los grandes dinosaurios carnívoros, vivió durante una época en que los dinosaurios eran pequeños e insignificantes. Era una época en que los reptiles no dinosaurios eran los dominantes del planeta, y fue un punto de inflexión importante en la ecología de la Tierra. La fauna de vertebrados de la Formación Ischigualasto y un poco más tarde Formación Los Colorados consistían principalmente en una variedad de arcosaurios crurotarsos y sinápsidos. En la Formación Ischigualasto, los dinosaurios constituían sólo alrededor del 6%[66] del número total de fósiles, pero al final del Triásico, los dinosaurios se estaban convirtiendo en los grandes animales terrestres dominantes y los otros arcosaurios y sinápsidos disminuyeron en variedad y cantidad.
Referencias
- Reig, O.A. (1963). «La presencia de dinosaurios saurisquios en los "Estratos de Ischigualasto" (Mesotriásico Superior) de las provincias de San Juan y La Rioja (República Argentina)». Ameghiniana 3 (1): 3-20.
- Sereno, P.C.; Novas, F.E.; Arcucci, A.B.; C. Yu (1988). «New evidence on dinosaur and mammal origins from the Ischigualasto Formation (Upper Triassic, Argentina)». Journal of Vertebrate Paleontology 8 (3, supplement): 26A.
- Sereno, P.C.; Novas, F.E. (1992). «The complete skull and skeleton of an early dinosaur». Science 258: 1137-1140. PMID 17789086. doi:10.1126/science.258.5085.1137.
- Paul, G.S. (1988). Predatory Dinosaurs of the World. New York: Simon and Schuster. pp. 248–250. ISBN 978-0-671-68733-5.
- Langer, Max C. (2004). «Basal Saurischia». En Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.), ed. The Dinosauria (2nd edición). Berkeley: University of California Press. pp. 25–46. ISBN 0-520-24209-2.
- Sereno, P.C.; Novas, F.E. (1993). «The skull and neck of the basal theropod Herrerasaurus ischigualastensis.». Journal of Vertebrate Paleontology 13: 451-476.
- Sereno, P.C. (2007). «The phylogenetic relationships of early dinosaurs: a comparative report». Historical Biology 19 (1): 145-155. doi:10.1080/08912960601167435. ISSN 0891-2963.
- McDowell, S.B., Jr.; and C.M. Bogert (1954). «The systematic position of Lanthanotus and the affinities of the anguinomorphan lizards». Bulletin of the American Museum of Natural History 105: 1-142.
- Novas, F. E. (1994). «New information on the systematics and postcranial skeleton of Herrerasaurus ischigualastensis (Theropoda: Herrerasauridae) from the Ischigualasto Formation (Upper Triassic) of Argentina». Journal of Vertebrate Paleontology 13 (4): 400-423.
- Sereno, P.C. (1993). «The pectoral girdle and forelimb of the basal theropod Herrerasaurus ischigualastensis.». Journal of Vertebrate Paleontology 13: 425-450.
- Novas, 1993. New information on the systematics and postcranial skeleton of Herrerasaurus ischigualastensis (Theropoda: Herrerasauridae) from the Ischigualasto Formation (Upper Triassic) of Argentina. Journal of Vertebrate Paleontology. 13, 400-423.
- Sereno, 1993. The pectoral girdle and forelimb of the basal theropod Herrerasaurus ischigualastensis. Journal of Vertebrate Paleontology. 13, 425-450.
- Sereno and Novas, 1993. The skull and neck of the basal theropod Herrerasaurus ischigualastensis. Journal of Vertebrate Paleontology. 13, 451-476.
- Sereno, P.C.; Forster, C.A.; Rogers, R.R.; Monetta, A.M. (1993). «Primitive dinosaur skeleton from Argentina and the early evolution of Dinosaurs». Nature 361 (6407): 64-66. doi:10.1038/361064a0.
- Rogers, R. R.; Swisher III, C.C.; Sereno, P.C.; Monetta, A.M.; Forster, C.A.; Martinez, R.N. (1993). «The Ischigualasto tetrapod assemblage (Late Triassic, Argentina) and 40Ar/39Ar dating of dinosaur origins». Science 260: 794-797. PMID 17746113. doi:10.1126/science.260.5109.794.
- Novas, F. E. (1986). «Un probable terópodo (Saurischia) de la Formación Ischigualasto (Triásico superior), San Juan, Argentina». IV Congreso Argentino de Paleontología y Bioestratigrafía, Mendoza, november 23-27 2: 1-6.
- Steel, R. (1970). «Part 14. Saurischia. Handbuch der Paläoherpetologie/Encyclopedia of Paleoherpetology». Gustav Fischer Verlag, Stuttgart: 1-87.
- Galton, P.M. (1973). «On the anatomy and relationships of Efraasia diagnostica (Huene) n.gen., a prosauropod dinosaur (Reptilia: Saurischia) from the Upper Triassic of Germany.». Paläontologische Zeitschrift 47 (3/4): 229-255.
- Paul, G.S. (1984). «The segnosaurian dinosaurs: relics of the prosauropod-ornithischian transition?». Journal of Vertebrate Paleontology 4: 507-515.
- Novas, F. E. (1989). «The tibia and tarsus in Herrerasauridae (Dinosauria, incertae sedis) and the evolution and origin of the dinosaurian tarsus». Journal of Paleontology 63: 677-690.
- Novas, F. E. (1992). «Phylogenetic relationships of the basal dinosaurs, the Herrerasauridae». Palaeontology 35: 51-62.
- Brinkman, D.B.; Sues, H.-D. (1987). «A staurikosaurid dinosaur from the Upper Triassic Ischigualasto Formation of Argentina and the relationships of the Staurikosauridae». Palaeontology 30: 493-503.
- Gauthier, J.A., Cannatella, D., Queiroz, K., Kluge, A.G., and Rowe, T. (1989), «Tetrapod phylogeny», en Fernholm, K. Bremer, & Jörnvall, H., ed., The Hierarchy of Life, Ámsterdam: Elsevier Science Publishers B. V., pp. 337-353.
- «The Nature of the Beast». The Dinosaurs!. Episodio 3. 1990.
- Holtz, T.R., Jr.; Padian, K. (1995). «Definition and diagnosis of Theropoda and related taxa». Journal of Vertebrate Paleontolology 15 (3, supplement): 35A.
- Bonaparte, J.F.; Pumares, J.A. (1995). «Notas sobre el primer cráneo de Riojasaurus incertus (Dinosauria, Prosauropoda, Melanorosauridae) del Triásico superior de La Rioja, Argentina». Ameghiniana 32: 341-349.
- Rauhut, O.W.M. (2003). «The interrelationships and evolution of basal theropod dinosaurs». Special Papers in Palaeontology 69: 1-213.
- Bittencourt, J.; Kellner, W.A. (2004). «The phylogenetic position of Staurikosaurus pricei from the Triassic of Brazil». Journal of Vertebrate Paleontology 24 (3, supplement): 39A.
- Langer, M.C; Benton, M.J. (2006). «Early dinosaurs: a phylogenetic study». Journal of Systematic Palaeontology 4 (4): 309-358. doi:10.1017/S1477201906001970.
- Irmis, Randall B.; Nesbitt, Sterling J.; Padian, Kevin; Smith, Nathan D.; Turner, Alan H.; Woody, Daniel; Downs, Alex (2007). «A Late Triassic dinosauromorph assemblage from New Mexico and the rise of dinosaurs». Science 317: 358-361. PMID 17641198. doi:10.1126/science.1143325.
- White, A.T.; Kazlev, M.A. (2003). «Theropoda: Basal Theropods». Palaeos.com. Archivado desde el original el 11 de julio de 2007. Consultado el 20 de julio de 2007.
- Forster, C.A.; Arcucci, A.B.; Marsicano, C.A.; Abdala, F.; May, C.L. (1995). «New vertebrate material from the Los Rastros Formation (Middle Triassic), La Rioja province, northwestern Argentina». Journal of Vertebrate Paleontology 15 (3, supplement): 29A.
- Arcucci, A. C.; Forster, C.; May, Abdala, F.; Marsicano, C. (1998). «Una nueva icnofauna de la Formación Los Rastros, Triásico Medio, en la Quebrada de Ischichuca Chica (Provincia de La Rioja, Argentina)». Acta Geológica Lilloana 18: 152.
- Bakker, R.T.; Galton, P.M. (1974). «Dinosaur monophyly and a new class of vertebrates». Nature 248: 168-172. doi:10.1038/248168a0.
- Bonaparte, J. F. (1976). «Pisanosaurus mertii Casimiquela and the origin of the Ornithischia». Journal of Paleontology 50: 808-820.
- Colbert, E.H. (1970). «A saurischian dinosaur from the Triassic of Brazil». American Museum Novitates 2405: 1-39.
- Long, R.A.; Murry, P.A. (1995). «Late Triassic (Carnian and Norian) Tetrapods from the Southwestern United States». New Mexico Museum of Natural History and Science, Bulletin 4.: 1-254.
- Hunt, A.P.; Lucas, S.G.; Heckert, A.B.; Sullivan, R.M.; Lockley, M.G. (1998). «Late Triassic Dinosaurs from the Western United States». Geobios 31 (4): 511-531. doi:10.1016/S0016-6995(98)80123-X.
- Chatterjee, S.; Creisler, B.S. (1994). «Alwalkeria (Theropoda) and Morturneria (Plesiosauria), new names for preoccupied Walkeria Chatterjee, 1987 and Turneria Chatterjee and Small, 1989». Journal of Vertebrate Paleontology (1): 142. Texto «volume14» ignorado (ayuda)
- Kischlat, E.-E. (1999). «A new dinosaurian “rescued” from the Brazilian Triassic: Teyuwasu barberenai, new taxon». 1st Simp. Brasil. Pal. Vert, Paleontologia em Destaque 14 (26): 58.
- Sereno, P.C. (1998). «A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria». Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 210 (1): 41-83.
- Gilmore, Charles W. (1920). «Osteology of the carnivorous dinosauria in the United States National Museum, with special reference to the genera Antrodemus (Allosaurus) and Ceratosaurus». Bulletin of the United States National Museum 110 (110): 1-159. doi:10.5479/si.03629236.110.i. hdl:2027/uiug.30112032536010.
- Gauthier, J.A.; Nesbitt, S.J.; Schachner, E.R.; Bever, G.S.; Joyce, W.G. (2011). «The bipedal stem crocodilian Poposaurus gracilis: inferring function in fossils and innovation in archosaur locomotion». Bulletin of the Peabody Museum of Natural History 52 (1): 107-126. doi:10.3374/014.052.0102. Archivado desde el original el 24 de septiembre de 2015.
- Chatterjee, Sankar (1985). «Postosuchus, a new Thecodontian reptile from the Triassic of Texas and the origin of Tyrannosaurs». Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 309 (1139): 395-460. Bibcode:1985RSPTB.309..395C. doi:10.1098/rstb.1985.0092.
- Benedetto, J.L. (1973). «Herrerasauridae, nueva familia de saurisquios triásicos». Ameghiniana 10 (1): 89-102.
- Galton, P.M. (1977). «On Staurikosaums pricei, an early saurischian dinosaur from the Triassic of Brazil, with notes on the Herrerasauridae and Poposauridae». Paläontologische Zeitschrift 51 (3): 234-245. ISSN 0031-0220. doi:10.1007/BF02986571.
- Brinkman, D.B.; Sues, H.D. (1987). «A staurikosaurid dinosaur from the Upper Triassic Ischigualasto Formation of Argentina and the relationships of the Staurikosauridae». Palaeontology 30 (3): 493-503. Archivado desde el original el 8 de diciembre de 2015. Consultado el 10 de abril de 2019.
- Novas, F.E. (1992). «Phylogenetic relationships of the basal dinosaurs, the Herrerasauridae». Palaeontology 35 (1): 51-62. Archivado desde el original el 8 de diciembre de 2015. Consultado el 10 de abril de 2019.
- Sereno, P.C. (1998). «A rational for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen 210 (1): 41-83.
- Langer, M. (2004). «Basal Saurischia». En Weishampel, David B.; Osmólska, Halzska; Dodson, Peter, eds. The Dinosauria (2nd ed.). Indiana University Press. pp. 40–42. ISBN 978-0-520-25408-4.
- Padian, K.; May, C.L. (1993). The Earliest Dinosaurs. En Lucas, Spencer G.; Morales, M., eds. «The Nonmarine Triassic». New Mexico Museum of Natural History & Science Bulletin 3. pp. 379-381.
- Novas, F.E. (1994). «New information on the systematics and postcranial skeleton of Herrerasaurus ischigualastensis (Theropoda: Herrerasauridae) from the Ischigualasto Formation (Upper Triassic) of Argentina». Journal of Vertebrate Paleontology 34 (4): 400-423. doi:10.1080/02724634.1994.10011523.
- Novas, F.E. (1996). «Dinosaur Monophyly». Journal of Vertebrate Paleontology 16 (4): 723-741. JSTOR 4523770. doi:10.1080/02724634.1996.10011361.
- Langer, M.C.; Ezcurra, M.D.; Bittencourt, J.S.; Novas, F.E. (2010). «The origin and early evolution of dinosaurs». Biological Reviews 85 (1): 55-110. PMID 19895605. doi:10.1111/j.1469-185X.2009.00094.x.
- Nesbitt, S.J.; Smith, N.D.; Irmis, R.B.; Turner, A.H.; Downs, A.; Norell, M.A. (2009). «A Complete Skeleton of a Late Triassic Saurischian and the Early Evolution of Dinosaurs». Science 326 (5959): 1530-1533. Bibcode:2009Sci...326.1530N. ISSN 0036-8075. PMID 20007898. doi:10.1126/science.1180350.
- Ezcurra, M.D. (2010). «A new early dinosaur (Saurischia: Sauropodomorpha) from the Late Triassic of Argentina: a reassessment of dinosaur origin and phylogeny». Journal of Systematic Palaeontology 8 (3): 371-425. doi:10.1080/14772019.2010.484650.
- Alcober, Oscar A.; Martínez, Ricardo N. (2010). «A new herrerasaurid (Dinosauria, Saurischia) from the Upper Triassic Ischigualasto Formation of northwestern Argentina». ZooKeys (63): 55-81. PMC 3088398. PMID 21594020. doi:10.3897/zookeys.63.550.
- Martínez, R.N.; Sereno, P.C.; Alcober, O.A.; Colombi, C.E.; Renne, P.A.; Montañez, I.P.; Currie, B.S. (2011). «A Basal Dinosaur from the Dawn of the Dinosaur Era in Southwestern Pangaea». Science 331 (6014): 206-210. Bibcode:2011Sci...331..206M. ISSN 0036-8075. PMID 21233386. doi:10.1126/science.1198467.
- Bittencourt, J.S.; Arcucci, A.B.; Marsicano, C.A.; Langer, M.C. (2014). «Osteology of the Middle Triassic archosaur Lewisuchus admixtus Romer (Chan~ares Formation, Argentina), its inclusivity, and relationships amongst early dinosauromorphs». Journal of Systematic Palaeontology 0 (3): 189-219. doi:10.1080/14772019.2013.878758.
- Baron, M.G., Norman, D.B., and Barrett, P.M. (2017). A new hypothesis of dinosaur relationships and early dinosaur evolution. Nature, 543: 501–506. doi 10.1038/nature21700
- Colbert, E.H. (1970). «A saurischian dinosaur from the Triassic of Brazil». American Museum Novitates 2405: 1-39.
- Long, R.A.; Murry, P.A. (1995). «Late Triassic (Carnian and Norian) Tetrapods from the Southwestern United States». New Mexico Museum of Natural History and Science, Bulletin 4.: 1-254.
- Hunt, A.P.; Lucas, S.G.; Heckert, A.B.; Sullivan, R.M.; Lockley, M.G. (1998). «Late Triassic Dinosaurs from the Western United States». Geobios 31 (4): 511-531. doi:10.1016/S0016-6995(98)80123-X.
- Chatterjee, S.; Creisler, B.S. (1994). «Alwalkeria (Theropoda) and Morturneria (Plesiosauria), new names for preoccupied Walkeria Chatterjee, 1987 and Turneria Chatterjee and Small, 1989». Journal of Vertebrate Paleontology 14 (1): 142. doi:10.1080/02724634.1994.10011546.
- Kischlat, E.-E. (1999). «A new dinosaurian "rescued" from the Brazilian Triassic: Teyuwasu barberenai, new taxon». Paleontologia em Destaque, Boletim Informativo da Sociedade Brasileira de Paleontologia 14 (26): 58.
- Ezcurra, M.D. (2010). "A new early dinosaur (Saurischia: Sauropodomorpha) from the Late Triassic of Argentina: a reassessment of dinosaur origin and phylogeny." Journal of Systematic Palaeontology, 8: 371-425.
- Sues, H.D. (1990). «Staurikosaurus and Herrerasauridae». En Weishampel, D.B.; Dodson, P.; Osmólska, H., ed. The Dinosauria. University of California Press. pp. 143-47. ISBN 0-520-06726-6.
- Hollocher, K.T.; Alcober, O.A.; Colombi, C.E.; Hollocher, T.C. (2005). «Carnivore coprolites from the Upper Triassic Ischigualasto Formation, Argentina: chemistry, mineralogy, and evidence for rapid initial mineralization». PALAIOS 20: 51-63. doi:10.2110/palo.2003.p03-98.
- Schmitz, L.; Motani, R. (2011). «Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology». Science 332 (6030): 705-8. Bibcode:2011Sci...332..705S. PMID 21493820. doi:10.1126/science.1200043.
- Bonaparte, J. F. (1982). «Faunal replacement in the Triassic of South America». Journal of Vertebrate Paleontology 2: 362-371.
- Bonaparte, J.F. (1979). «Faunas y paleobiogeografía de los tetrápodos mesozoicos de América del Sur». Ameghiniana, Revista de la Asociación Paleontológica Argentina 16 (3–4): 217-238.
- Benton, Michael J. (1999), «Origin and early evolution of dinosaurs», en Farlow, James O.; Brett-Surman, M.K., eds., The Complete Dinosaur, Indiana University Press, pp. 204-215, ISBN 0-253-21313-4.
- Parrish, J. Michael (1999), «Evolution of the archosaurs», en Farlow, James O.; Brett-Surman, M.K., eds., The Complete Dinosaur, Indiana University Press, pp. 191-203, ISBN 0-253-21313-4.
- Rothschild, B., Tanke, D. H., and Ford, T. L., 2001, Theropod stress fractures and tendon avulsions as a clue to activity: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, pp. 331–336.
- Molnar, R. E., 2001, Theropod paleopathology: a literature survey: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, pp. 337–363.
- Sill, W.D. (1974). «The anatomy of Saurosuchus galilei and the relationships of the rauisuchid thecodonts». Bulletin of the Museum of Comparative Zoology 146: 317-362.
- Bonaparte, J.F. (1970). «Annotated list of the South American Triassic tetrapods». Gondwana Symposium Proceedings and Papers 2: 665-682.
Enlaces externos
 Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Wikimedia Commons alberga una categoría multimedia sobre Herrerasaurus ischigualastensis.
Wikimedia Commons alberga una categoría multimedia sobre Herrerasaurus ischigualastensis. Wikispecies tiene un artículo sobre Herrerasaurus ischigualastensis.
Wikispecies tiene un artículo sobre Herrerasaurus ischigualastensis.- en duiops
- en Thescelosaurus
| Control de autoridades |
|
|---|
 Datos: Q20718404
Datos: Q20718404- Especies: Herrerasaurus ischigualastensis