Núcleo sexualmente dimórfico
El núcleo sexualmente dimórfico (NSD) o (SDN por sus siglas sexually dimorphic nucleus) es un núcleo ovoide, de grandes células y densamente empaquetado que se localiza en el área preóptica medial del hipotálamo. Se cree que su función está relacionada con la conducta sexual en animales.
| Núcleo sexualmente dimórfico | ||
|---|---|---|
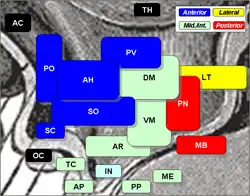 Área preóptica, del hipotálamo 'PO' en azul a la izquierda. | ||
| Latín | [TA]: Area praeoptica | |
| Es parte de | Hipotálamo | |
| Enlaces externos | ||
| NeuroLex ID | Núcleo sexualmente dimórfico del área preóptica | |
Para todas las especies de mamíferos en que se ha investigado el NSD presenta dimorfismo sexual siendo considerablemente mayor en machos (hasta dos veces mayor) que en las hembras. Además en humanos toma forma elongada en hombres mientras que es más esférico en hembras. A pesar de ello no se encuentran diferencias en el NSD humano en cuanto a la densidad de células o en el diámetro celular medio.[1] El volumen y el número de células en el NSD desciende considerablemente con la edad, sin embargo esta disminución es dependiente también del sexo y en hombres se encuentra un sustancial decremento entre los 50-60 años. La muerte celular en esta área es más común en mujeres que en hombres, especialmente en aquellas mayores de 70 años. Esta disminución puede alcanzar el 10-15 % con respecto a las encontradas en la niñez.
El NSD y sus homólogos existen en humanos, mamíferos y en el encéfalo de algunos otros animales, incluyendo:
- El tercer núcleo intersticial del hipotálamo anterior (INAH3) en humanos;
- El núcleo sexualmente dimórfico ovino, (NSDo) en el área preóptica medial del hipotálamo en ovejas;[2]
- Núcleo sexualmente dimórfico del área preóptica (NSD-APO) en ratas;
- Núcleo anterior hipotalámico (AHdc por sus siglas en inglés) en macacos;
- Área específica en el núcleo preóptico medial (APOm) en codornices; etc.[3][4]
Formación y organización del NSD en el área preóptica medial
Diferencias causadas por exposición prenatal a testosterona
El volumen del NSD en el área preóptica medial se modifica por la acción hormonal y se ha demostrado que la testosterona tiene una gran importancia en este proceso. De esta manera el mayor volumen en machos está correlacionado con la mayor concentración fetal de testosterona de estos. Además hay evidencia de que la testosterona actúa durante un periodo prenatal específico promoviendo el desarrollo de neuronas que expresarán aromatasa en el NSD de los machos (la testosterona se transforma en estradiol por la acción de la aromatasa). El efecto de la testosterona se refleja también por la influencia de la posición intrauterina en la morfología del NSD. Algunos estudios han demostrado que ratas macho gestadas entre otros dos fetos de macho (2M) tienen hasta dos veces mayor volumen en el NSD que aquellas gestadas entre dos fetos de hembras (2F). De la misma manera, los niveles de testosterona y de 17β-estradiol (producto de la aromatización de la testosterona) son significativamente mayores en los machos 2M que en los machos 2F en el día 21 de la gestación (la testosterona puede ser transferida desde fetos adyacentes a los fetos experimentales). A pesar de ello no hay evidencias de relación alguna entre el volumen del NSD y la posición fetal en hembras.[5][6]
Diferencias causadas por apoptosis postnatal
De acuerdo con algunos estudios, la diferencia de volumen en el NSD entre machos y hembras es debida a a la muerte celular programada (apoptosis) durante el desarrollo temprano tras el nacimiento. En ratas, la división central del núcleo preóptico medial (NSD-cmAPO) es un componente importante del NSD-APO y las evidencias muestran que el número de células apoptóticas es mayor en hembras que en machos entre el día postnatal (DP) 7 y el DP10. En el cmAPO, los niveles de algunas proteínas relacionadas con la apoptosis son diferentes en hembras que en machos, incluyendo las proteínas Bcl-2 y Bax. Los niveles de Bcl-2, una proteína antiapoptótica, son mucho mayores en machos de rata en el DP8 en comparación con los niveles encontrados en hembras, causando que el número de células apoptóticas a esa edad sean menores en machos. Por otra parte, Bax, una proteína proapoptótica, muestra menores niveles en estos mismos machos que en hembras. Además, el número de células inmunoreactivas para caspasa-3 es mayor en hembras que en machos, indicando un alto nivel de apoptosis en el cmAPO.
La apoptosis también ocurre en el núcleo periventricular anteroventral (avNP), que también es un área sexualmente dimórfica localizada en el área gris periventricular, en el extremo rostral del tercer ventrículo. A diferencia del NSD, el avNP tiene mayor tamaño en hembras que en machos. Se ha demostrado que en hembras los niveles de Bcl-2 en el avNP son mayores mientras que los niveles de Bax son menores que en machos. De esta forma, la apoptosis tiene un rol crucial en la formación del NSD de forma que el número de células apoptóticas correlaciona negativamente con el volumen del NSD.[7]
Rol en los comportamientos sexuales en machos
El comportamiento sexual se divide en dos fases: la fase apetitiva, que contiene una secuencia variable de comportamientos como la atracción y el cortejo; y la fase consumatoria, durante la cual se dan comportamientos de cópula estereotipados. Se sabe que el área preóptica medial del encéfalo controla la expresión de ambos aspectos del comportamiento sexual. Así, lesiones del NSD-APO interrumpen el comportamiento copulatorio en ratas. Además, lesiones del cuerpo celular en el NSD pars compacta, homólogo al NSD en jerbos, produce severas disrupciones en el comportamiento copulatorio de los machos. Además, un estudio del núcleo preóptico medial en codornices (POm) muestra que la activación del comportamiento sexual en machos requiere de la aromatización de los andrógenos (testosterona) en estrógenos (17β-estradiol). Como en ratas, la expresión de aromatasa es específica del núcleo medial preóptico y la intensidad del comportamiento copulatorio correlaciona positivamente con el número de células que expresan aromatasa en la parte caudal del núcleo.[3]
Los comportamientos apetitivos son también controlados en parte por el área preóptica medial. De esta manera, los ratones nulos para aromatasa muestran defectos en la motivación sexual. Sin embargo esta disrupción se da cuando se lesiona la parte rostral y no la caudal del área preóptica medial. Las lesiones en la parte rostral también disminuyen la preferencia de las ratas hembra por los machos. Además, mediante microdiálisis cerebral in vivo se ha demostrado que los niveles extracelulares de dopamina en el APOm incrementan a medida que progresan los comportamientos apetitivos o de aproximación. El rol del APOm en el control de los comportamientos sexuales apetitivos se ha confirmado también con manipulaciones farmacológicas del sistema dopaminérgico en la misma. En ratas, lesiones del NSD-APO pueden eliminar completamente el comportamiento copulatorio de machos, pero solo disminuyen los comportamientos apetitivos, lo que sugiere que hay otras partes del encéfalo responsables de estos comportamientos.[3]
Rol en la preferencia de pareja sexual
Rol en la preferencia de pareja sexual en machos
Investigaciones en el núcleo sexualmente dimórfico ovino demuestran que el volumen del NSDo varía en función preferencia de pareja en machos. En machos homosexuales (aproximadamente el 8 % de la población) el NSDo tiene aproximadamente la mitad de tamaño que en los heterosexuales.[2] En un estudio en ovejas, cuatro machos heterosexuales y nueve homosexuales fueron expuestos a dos hembras sexualmente receptivas y a dos machos, registrando el comportamiento copulatorio (montas y eyaculaciónes). Los machos heterosexuales mostraron más montas y eyaculaciones con hembras que con machos, mientras que los homosexuales mostraron el patrón contrario. El análisis del tejido cerebral en estos animales mostró que el NSDo es aproximadamente dos veces mayor en heterosexuales que en homosexuales y además tiene un mayor número de neuronas, aunque la densidad celular es similar en ambos grupos. Además, la cantidad de aromatasa es significativamente mayor en heterosexuales que en homosexuales.[2]
Otras especies muestran una relación similar entre la preferencia de pareja sexual y el volumen del NSD, por ejemplo, el INAH-3 en humanos (homólogo al NSDo) es significativamente mayor en hombres heterosexuales que en homosexuales.
Daños en el NSD y cambios en la preferencia sexual en machos
Lesiones bilaterales del NSD del área preóptica medial en hurones macho sexualmente experimentados cambian la preferencia de los mismos a patrones similares a los que se dan en hembras. De esta forma es probable que el NSD juegue un papel importante en el apareamiento y la reproducción.[8]
Rol del NSD en la preferencia de pareja sexual en hembras
A pesar de que el NSD muestra una relación con la preferencia de pareja sexual en machos no muestra la misma relación en hembras. Hay evidencias de que el NSD no es necesario para la expresión de comportamientos sexuales típicos o para la preferencia de pareja en hembras. Parte de la evidencia proviene del estudio de hembras de Macaco japonés donde se examina el núcleo anterior hipotalámico (NAH, homólogo al NSD. En esta especie la hembra rutinariamente corteja, monta, compite por e incluso prefiere ciertas parejas sexuales femeninas antes que a algunos machos. Comparando el NAH de Macacos japoneses con el de Monos rhesus, una especie cercana, no se encuentran diferencias de volumen entre los machos típicos y las hembras típicas.[4]
Rol del NSD en la orientación sexual humana
EN 1991 LeVay publicó un estudio en el que analizaba a 16 hombres Heterosexuales, 19 hombres Homosexuales y 6 mujeres heterosexuales Post mortem buscando diferencias en el núcleo intersticial anterior del hipotálamo (INAH 1, 2, 3, 4). Este estudió encontró que el INAH-3era mayor en hombres heterosexuales en comparación con hombres homosexuales y mujeres heterosexuales. Este estudio mostró por primera vez el NSD como área dimórfica entre hombres homosexuales y heterosexuales.[9] Su estudio sin embargo fue criticado porque una parte de sus pacientes eran enfermos de SIDA, pudiendo alterar sus resultados.
Un estudio posterior determinó que el tamaño del INAH3 en hombres homosexuales tiene un tamaño intermedio entre el de los hombres y el de las mujeres heterosexuales. También determinó que la razón de esta diferencia de tamaño es debida a que los hombres homosexuales mostraban una densidad neuronal mayor que la de los hombres heterosexuales pero no se encontraron diferencias en el área neuronal. Este estudio mostró que las diferencias obtenidas por LeVay no se debían a la infección por VIH.[10]
Estos estudios no corroboraron que los hombres homosexuales tuvieran un hipotálamo totalmente femenino. De esta forma aunque el INAH3 es más pequeño en hombres homosexuales, no llega a tener el tamaño típico de mujeres heterosexuales.[11] Por otro lado, el núcleo supraquiasmático (SCN) de hombres homosexuales es mayor que el de heterosexuales; en este caso tanto el número de neuronas como el volumen del núcleo es casi el doble en hombre homosexuales. A pesar de ello, estas áreas del hipotálamos no han sido estudiadas en mujeres homosexuales ni en hombres o mujeres bisexuales. En definitiva, los estudios contemporáneos a Levay dudaron de la hipótesis inicial en la que se afirmaba que los hombres homosexuales tienen un hipotálamo femenino y de que la clave de esta diferenciación a un "cerebro masculíno desde un cerebro inicialmente femeníno" es la influencia epigenética de la testosterona durante el desarrollo prenatal.[12][13]
Efectos de proteínas y otras moléculas en el NSD del área preóptica medial
NELL2
NELL2 es una proteína del sistema nervioso específica de algunos tejidos. Contiene un dominio similar al factor de crecimiento epidérmico (FCE)y su expresión génica está regulada por estrógenos. NELL2 también tiene un rol neuroprotector en el hipocampo en la rata. Cuando se bloquea la síntesis de NELL2 neonatal mediante la inyección de un oligodesoxinucleótido antisentido en ratas (DP0-DP5) el tamaño del NSD disminuye. Ya que el volumen está relacionado con la testosterona es posible que esto se deba al efecto neuroprotector del estradiol, aromatizado desde la testosterona.[14]
Somatostatina
La transcripción específica de cada sexo del gen de la somatostatina coincide con el establecimiento de las diferencias en el NSD entre machos y hembras. Desde el DP8 hasta el DP35 el área de las células que expresan ARNm de somatostatina es significativamente mayor en machos que en hembras, siendo el mayor tamaño en el DP15. Es posible que la somatostatina esté relacionada con la organización dependiente de estrógenos del NSD.[15]
Fenitrotión
El Fenitrotión es una forma de organofosfato que puede afectar al desarrollo del sistema reproductivo en machos de rata. El tratamiento con fenitrotión en machos de rata causa regresión de los órganos que dependen de andrógenos ya que es un potente antagonista competitivo del receptor de andrógenos. Su efecto en el NSD-APO es diferente en machos y en hembras de rata. En machos la exposición prenatal causa un incremento significativo en el NSD-APO; en hembras la exposición prenatall disminuye significativamente el volumen del mismo núcleo. Una explicación posible es que el fenitrotión cambie el comportamiento de la aromatasa, alterando entonces la cantidad de estrógeno que son convertidos a andrógenos.[16]
Dietilestilbestrol
El Dietilestilbestrol (DES) es un estrógeno sintético no esteroideo utilizado para prevenir posibles abortos. El DES tiene además efectos en el desarrollo del sistema reproductivo. A bajas dosis disminuye la concentración de testosterona plasmática de ratas macho mientras que promueve la maduración folicular en hembras. Se ha demostrado que el tratamiento con DES aumenta el volumen del NSD en hembras. En cambio no altera a bajas dosis el tamaño del NSD en machos de rata.[17]
Otras áreas sexualmente dimórficas en el encéfalo
El núcleo ventromedial del hipotálamo (NVM) ha sido considerado un núcleo sexualmente dimórfico. Es una región importante para regular el comportamiento sexual en hembras de roedor. Las neuronas del NVM son dependientes de estrógenos y muestran gran plasticidad estructural. La organización sináptica del NVM es sexualmente dimórfica y las hembras tienen más sinapsis químicas dendritícas en el NVM mientras que los machos muestran más sinapsis somáticas. Además, el tamaño de las densidades postsinápticas axodendríticas y axosomáticas es también sexualmente dimórfico, siendo mayor en machos que en hembras.
Los estrógenos también juegan un rol importante modulando la conectividad en el NVM. Estos estrógenos varían durante el ciclo estral siendo altos en el proestro de ratas y bajos en el diestro. De la misma forma, el volumen celular en el NVM en hembras de rata en proestro y en machos es mayor que en hembras en diestro. Además, las ratas en proestro tienen una mayor densidad de sinapsis en el NVM que aquellas en diestro. Por otra parte, el ácido gamma-butírico juega un rol en el desarrollo del NVM y en la diferenciación sexual.[19][20]
Véase también
Referencias
- Hofman, M.A.; Swaab D.F. (1989). «The sexually dimorphic nucleus of the preoptic area in the human brain: a comparative morphometric study». Journal of Anatomy 164: 55-72. PMC 1256598. PMID 2606795.
- Roselli, C.; Larkin, K.; Resko, J.; Stellflug, J.; Stormshak, F. (2004). «Volume of a Sexually Dimorphic Nucleus in the Ovine Medial Preoptic Area/Anterior Hypothalamus Varies with Sexual Partner Preference». Endocrinology 145 (2): 478-483. PMID 14525915. doi:10.1210/en.2003-1098.
- Balthazart J, Ball G (2007). «Topography in the preoptic region: Differential regulation of appetitive and consummatory male sexual behaviors». Frontiers in Neuroendocrinology 28 (4): 161-178. PMC 2100381. PMID 17624413. doi:10.1016/j.yfrne.2007.05.003.
- Vasey, P., Pfaus, J. (2005). «A sexually dimorphic hypothalamic nucleus in a macaque species with frequent female=female mounting and same-sex sexual partner preference». Behavioural Brain Research 157 (2): 265-272. PMID 15639177. doi:10.1016/j.bbr.2004.07.005.
- Pei, M.; Matsuda, K.; Sakamoto, H.; Kawata, M. (2006). «Intrauterine proximity to male fetuses affects the morphology of the sexually dimorphic nucleus of the preoptic area in the adult rat brain». European Journal of Neuroscience 23 (5): 1234-1240. PMID 16553785. doi:10.1111/j.1460-9568.2006.04661.x.
- Roselli, C., Stadelman, H., Reeve, R., Bishop, C., Stormshak, F. (2007). «The ovine sexually dimorphic nucleus of the medial preoptic area is organized prenatally by testosterone». Endocrinology 148 (9): 4450-4457. PMID 17540718. doi:10.1210/en.2007-0454.
- Tsukahara, S., Kakeyama, M., Toyofuku, Y. (2006). «Sex Differences in the level of Bcl-2 Family Proteins and Caspase-3 Activation in the Sexually Dimorphic Nuclei of the Preoptic Area in Postnatal Rats». Journal of Neurobiology 66 (13): 1411-1419. PMID 17013925. doi:10.1002/neu.20276.
- Alekseyenko, O., Waters, P., Zhou, H., Baum, M. (2007). «Bilateral damage to the sexually dimorphic medial preoptic area/anterior hypothalamus of male ferrets causes a female-typical preference for and a hypothalamic Fos response to male body odors». Physiology & Behavior 90 (2–3): 438-449. PMC 2265004. PMID 17118411. doi:10.1016/j.physbeh.2006.10.005.
- LeVay, S. (1991). «A difference in hypothalamic structure between heterosexual and homosexual men». Science 253 (5023): 1034-1037. PMID 1887219. doi:10.1126/science.1887219.
- Byne, W.; Lasco, M. S.; Kemether, E.; Shinwari, A.; Edgar, M. A.; Morgello, S.; Jones, L. B.; Tobet, S. (21 de febrero de 2000). «The interstitial nuclei of the human anterior hypothalamus: an investigation of sexual variation in volume and cell size, number and density». Brain Res 856 (1-2): 254-8. ISSN 0006-8993. PMID 10677635. doi:10.1016/S0006-8993(99)02458-0.
- Poiani, Aldo (19 de agosto de 2010). Animal Homosexuality: A Biosocial Perspective (illustrated edición). Cambridge University Press, 2010. pp. 212–-3. ISBN 0521196752.
- http://www.hiim.unizg.hr/images/knjiga/CNS41.pdf - Judaš, M., Kostović, I., The Fundamentals of Neuroscience, ch. 41, «Neurobiology of emotions and sexuality», p. 408. (en croata)
- «Gender and sexual orientation in relation to hypothalamic structures.». Horm Res. 38 Suppl 2: 51-61. 1992. PMID 1292983. doi:10.1159/000182597.
- Ellis, S., Mouihate, A., Pittman, Q. (2006). «Neuroendocrine programming and sexual differentiation». Frontiers in Neuroendocrinology 27: 95-99. doi:10.1016/j.yfrne.2006.03.217.
- Chitose, O., Yasuhiko, K., Sakuma, Y. (2007). «Transient transcription of the somatostatin gene at the time of estrogen-dependent organization of the sexually dimorphic nucleus of the rat preoptic area». Endocrinology 148 (3): 1144-1149. PMID 17138650. doi:10.1210/en.2006-1214.
- Struve, M., Turner, K., Dorman, D. (2007). «Preliminary investigation of changes in the sexually dimorphic nucleus of the rat medial preoptic area following prenatal exposure to fenitrothion». Journal of Applied Toxicology 27 (6): 631-636. PMID 17582585. doi:10.1002/jat.1267.
- Yamamoto, M., Shirai, M., Tamura, A., Kobayashi, T., Kohara, S., Murakami, M., Arishima, K. (2005). «Effects of maternal exposure to a low dose of diethylstilbestrol on sexual dimorphic nucleus volume and male reproductive system in rat offspring». The Journal of Toxicological Sciences 30 (1): 7-18. PMID 15800398. doi:10.2131/jts.30.7.
- Slamberova, R., Vathy, I., Hnatczuk, O. (2004). «Expression of proopiomelanocortin and proenkephalin mRNA in sexually dimorphic brain regions are altered in adult male and female rats treated prenatally with morphine». Journal of Applied Toxicology 63 (5): 399-408. PMID 15140157. doi:10.1111/j.1399-3011.2004.00134.x.
- Susana, I., Dulce Madeira, M. (2005). «Estrogen Modulates the Sexually Dimorphic Synaptic Connectivity of the Ventromedial Nucleus». The Journal of Comparative Neurology 484 (1): 68-79. PMID 15717306. doi:10.1002/cne.20451.
- Zhou, J., Pfaff, D., Chen, G. (2005). «Sex Differences in estrogenic regulation of neuronal activity in neonatal cultures of ventromedial nucleus of the hypothalamus». PNAS 102 (41): 14907-14912. PMC 1253608. PMID 16204378. doi:10.1073/pnas.0507440102.
| Control de autoridades |
|
|---|
 Datos: Q7458854
Datos: Q7458854