Plutella xylostella
La palomilla dorso de diamante (Plutella xylostella), conocida también como polilla de la col o polilla del repollo (diamondback moth o cabbage moth, en inglés), es una especie de insecto lepidóptero de la familia Plutellidae que ha llegado a ser verdaderamente cosmopolita y es la única de todo el orden Lepidoptera (Insecta) con una distribución continua y global. Se ha convertido en la plaga más destructiva de cosechas de plantas alimentarias de la familia Brassicaceae, como Brassica napus (colza) y distintas variedades de Brassica oleracea, como B. oleracea var. capitata (repollo) y B. oleracea var. botrytis (coliflor).[1] Es muy difícil de controlar debido a que ha desarrollado resistencia a numerosos plaguicidas. De hecho es la primera especie en desarrollar resistencia a diclorodifeniltricloroetano (DDT), durante la década de 1950,[2] y frente a toxinas de Bacillus thuringiensis (toxinas Bt), durante la década de 1990.[3]
| Palomilla dorso de diamante | ||
|---|---|---|
 | ||
 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Arthropoda | |
| Clase: | Insecta | |
| Orden: | Lepidoptera | |
| Superfamilia: | Yponomeutoidea | |
| Familia: | Plutellidae | |
| Género: | Plutella | |
| Especie: |
P. xylostella (Linnaeus, 1758) | |
| Distribución | ||
 | ||
| Sinonimia | ||
| ||
Clasificación
La clasificación que se muestra en la ficha proviene de Arctos,[4] una base de datos de referencia en taxonomía. Otra clasificación más detallada es la recogida en la base de datos "Taxonomy" del Centro Nacional para la Información Biotecnológica (NCBI):[5]
Distribución
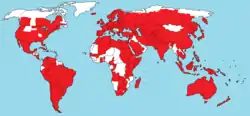
Se cree que el origen de P. xylostella es Europa, la región mediterránea y el sur de África.[6][7] Sin embargo hoy día puede encontrarse en toda América, Europa, Sudeste Asiático, Australia, Nueva Zelanda, India, y África.[8] A partir de datos y publicaciones disponibles, así como de localizaciones geográficas de cepas de P. xylostella empleadas en secuenciación genómica, You, M. et al, en su artículo de febrero de 2013,[9] realizaron un mapa de la distribución mundial de P. xylostella, el cual se puede consultar en la ficha de la especie.
Es una especie migratoria, capaz de cubrir largas distancias. Sus movimientos migratorios han sido estudiados en China.[10][11]
Biología
Descripción
Los huevos de P. xylostella tienen una forma oval aplanada y miden unos 0,44 mm de largo por 0,20 mm de ancho. Presentan un color que va desde el amarillo al verde pálido.[12]

Las larvas rara vez superan el tamaño promedio de 1,7 mm en el primer estadio larval, 3,5 mm en el segundo, 7 mm en el tercero y 12 mm en el cuarto. Son incoloras en el primer estadio pero adoptan un color verde a partir del segundo. Son estrechas en sus extremos y a lo largo del cuerpo de la larva se observan algunos pelos marcados por unas manchas de color blanco allí donde surgen. Presentan 5 pares de propatas, siendo característico el último par en la parte final posterior ya que se disponen en forma de “V”.[12]
El capullo tiene aspecto sedoso, es poco denso en fibras y suele medir de largo entre 7 y 9 mm.[12]

Los individuos adultos son pequeños y delgados, y miden unos 6 mm de largo. Tienen un color entre gris y marrón y presentan una banda a lo largo de la espalda de color crema o marrón claro. Dicha banda a veces se encuentra perfilada de tal manera que adopta la forma de uno o varios diamantes, lo que le confiere a Plutella xylostella su nombre vulgar de palomilla dorso de diamante.[12]

Ciclo de vida
El apareamiento de P. xylostella tiene lugar durante el anochecer (momento a partir del cual son más activos) del mismo día en el que los adultos emergen de la pupa. Las hembras comienzan la puesta de huevos poco después del apareamiento y puede durar hasta 4 días[1] (o hasta 10, según otras fuentes[12]), en los cuales se depositarán entre 11 y 188 huevos,[1] pudiendo llegar incluso a depositarse hasta 250-300.[12] Este proceso se ve favorecido por la falta de luz (es decir, en días poco luminosos). Sin embargo la puesta no parece inhibirse del todo cuando se hacen experimentos en los que se aplica luz durante el período nocturno. Otros factores que van a influenciar la puesta son la temperatura, la producción de compuestos volátiles y secundarios de la planta, y la presencia de tricomas y ceras en la superficie foliar.[1] Los huevos suelen depositarse preferentemente en depresiones de la hoja en vez de en regiones lisas.[1][12] El periodo de incubación de los huevos suele comprender entre 5 y 6 días, siendo la temperatura el principal factor que determina dicho período.[1][12]
Las larvas comienzan a alimentarse al poco tiempo de emerger de los huevos. Las larvas del primer estadio larval sólo se alimentan del mesófilo de las hojas, un tejido esponjoso. Por otro lado, las larvas más maduras se alimentan de toda la hoja comenzando por la superficie inferior hasta llegar a la capa cerosa y la epidermis superior, que quedan sin consumir, formando "ventanas" transparentes.[1][12] El tiempo de desarrollo medio para cada estadio larval es de 4,5 días para el primero, 4 días para el segundo, 4 días para el tercero y 5 días para el cuarto.[12] De nuevo aquí la temperatura va a determinar la duración, en este caso, de los cuatro estadios larvales, siendo el desarrollo de éstas más rápido en climas cálidos. Otro factor que influye en la tasa de desarrollo es la planta que elige la hembra como hospedadora de los huevos[1] (el desarrollo será más o menos rápido en función de la especie de crucífera en la que se hospedan).
Al final del cuarto estadio larval, la larva construye un capullo sobre la hoja de la que se ha alimentado. Entra así la larva en un estado de quiescencia denominado prepupa, que dura unos dos días, durante el cual muda su piel que se mantendrá aún unida a la región caudal de la pupa. La pupación suele durar entre 4 y 15 días,[1][12] dependiendo de la temperatura, tras la cual emerge un individuo adulto.[1]
Ecología
El origen de P. xylostella en un área determinada así como su capacidad de sobrevivir en dicha área durante los meses que no hay cosecha sigue siendo un tema de discusión hoy en día. Mientras que en regiones tropicales y subtropicales se pueden encontrar individuos de P. xylostella en cualquier estadio de su ciclo de vida a lo largo de todo el año, ya que las crucíferas (familia de plantas a las que P. xylostella está adaptada y a las que emplea como hospedadoras) están presentes todo el año en dichas regiones, en regiones templadas estas plantas no crecen todo el año, lo que ha llevado a algunos investigadores a pensar que las pupas y/o los adultos deben invernar en el interior de estas plantas hospedadoras.[1] La controversia ha dado lugar a estudios de laboratorio y de seguimiento en los que se ha evaluado la resistencia de P. xylostella a bajas temperaturas, determinándose que sí existe cierta resistencia y supervivencia[13][14][15][16] pero que no se da diapausa.[15] A pesar de que Dosdall, en 1994,[17] publicó un estudio en el que demostraban que P. xylostella invernaba, en un estadio posterior[13] reconoció que debía tratarse de un fenómeno aislado ya que no volvieron a encontrar nuevas evidencias; por otro lado afirman que la tendencia del calentamiento global puede conducir a un incremento de plagas de P. xylostella, seguramente por una mayor supervivencia durante los meses de invierno. Por tanto, en numerosas regiones templadas donde hay plagas de P. xylostella, esta no inverna y su origen parece ser migratorio[1][13][15] a pesar de que se ha documentado que pueden sobrevivir como poblaciones residentes que invernan.[16]
Una de las características que hacen de P. xylostella una de las plagas más globales es su capacidad de migración y de recorrer largas distancias, pudiéndose mantener en vuelo continuo en corrientes de aire desplazándose una distancia de hasta 1.000Km por día.[1] Es por ello que algunas poblaciones de P. xylostella surgen por movimientos migratorios de individuos adultos desde regiones cálidas a regiones templadas:[1][13][15] de Estados Unidos o México a Canadá, de regiones cálidas de Japón a regiones más frías, del Sur al norte de la China en primavera, y, seguramente, lo mismo ocurra en otras regiones templadas (Nueva Zelanda, Australia, Sudáfrica, Argentina y Chile, entre otras).
Plantas hospedadoras
P. xylostella se alimenta sólo de plantas de la familia Brassicaceae, tanto cultivadas como silvestres. Este es un grupo muy variado de plantas, algunas cultivadas por su uso alimentario. Dentro de esta familia, el rango de plantas que se ven afectadas por P. xylostella se limita a aquellas que contienen glucosinolatos,[1][18] los cuales van a estimular la ingesta y la puesta de huevos. Algunas de las especies y variedades cultivadas de las que se alimenta son:[1]
- B. oleracea var. capitata (repollo)
- B. oleracea var. botrytis (coliflor)
- B. oleracea var. italica (broccoli)
- Raphanus sativus (rábano)
- B. rapa (nabo) y algunas variedades asiáticas
- B. oleracea var. gemmifera (col de Bruselas)
- B. oleracea var. gongylodes (colirrábano)
- B. juncea (mostaza castaña)
- B. napus (colza)
- B. oleracea var. medulosa (col de meollo)
- Nasturtium officinale (berro de agua)
- B. oleracea var. alboglabra (brécol chino)
También se alimenta de numerosas crucíferas consideradas como malas hierbas, pero solo en ausencia de plantas cultivadas que emplear como hospedadoras. Estas plantas, como hospedadoras, son muy importantes ya que ayudan al mantenimiento de las poblaciones de P. xylostella en regiones templadas durante la primavera cuando aún no ha comenzado la época de cultivo.[1]
Se ha observado que P. xylostella puede alimentarse de y reproducirse en otras crucíferas como:[1]
Arabis glabra, Armoracia lapathifolia, Barbarea stricta, Barbarea vulgaris, Basela alba, Beta vulgaris, Brassica caulorapa, Brassica kaber (Sinapis arvensis), Brassica napobrassica, Bunias orientalis, Capsella bursa-pastoris, Cardamine amara, Cardamine cordifolia, Cardamine pratensis, Cheiranthus cheiri, Conringa orientalis, Descurainia sophia, Erysimum cheiranthoides, Galinsoga ciliata, Galinsoga parviflora, Hesperis matronalis, lberis amara, Isatis tinctoria, Lepidium perfoliatum, Lepidium virginicum, Lobularia maritima, Mathiola incana, Sisymbrium altissimum (Norta altissima), Pringlea antiscorbutica, Raphanus raphanistrum, Rorippa amphibia, Rorippa islandica, Sinapis alba, Sisymbrium austriacum, Sisymbrium officinale y Thlaspi arvense.
P. xylostella como plaga

P. xylostella se consideró como una plaga menor durante mucho tiempo, ya que su daño estaba enmascarado por el deterioro que llevaban a cabo especies como Pieris rapae (blanquita de la col) o Trichoplusia ni (palomilla nocturna; gusano falso medidor). Sin embargo la abundancia de P. xylostella empezó a aumentar durante los años 1950, llegando a ser un verdadero problema para los cultivos de crucíferas ya en los años 1970 y constatándose en los años 1980 el desarrollo de resistencia a insecticidas tales como los piretroides.[12]
En los últimos años, P. xylostella se ha convertido en la plaga más destructiva de cultivos de crucíferas en el mundo,[1] con un costo de entre $4-5 mil millones por año en daños y control de la plaga.[19][20] La ausencia de enemigos naturales para P. xylostella se ha postulado como una de las razones de su éxito como plaga mundial. La ausencia de éstos, tales como parasitoides, puede deberse a que P. xylostella presenta una mejor capacidad de establecerse en nuevos cultivos de crucíferas que los primeros. En este sentido se ha documentado en algunos estudios la capacidad de P. xylostella de recorrer grandes distancias, no habiéndose encontrado datos similares para sus enemigos naturales. Otra posibilidad es que el uso indiscriminado de insecticidas en un área determinada haya conllevado la muerte de los enemigos naturales de P. xylostella, razón que justifica la falta de control por parte de éstos sobre P. xylostella.[1]
Resistencia a insecticidas
Antes del uso de insecticidas sintéticos, introducidos a finales de los años 1940, P. xylostella no se consideraba una plaga. Ya a mediados de los años 1950, el uso indiscriminado de estos insecticidas sintéticos terminó por eliminar a los enemigos naturales de P. xylostella. Al eliminar los enemigos naturales de P. xylostella se incrementó aún más el uso de estos insecticidas, dando como resultado resistencias eventuales a insecticidas y fallos en el control de plagas de P. xylostella. Indirectamente se estaban seleccionando positivamente aquellos individuos que presentasen resistencia a insecticidas debido a la presión selectiva que se estaba ejerciendo en los mismos.[1] En 1953, P. xylostella se convirtió en el primer insecto que desarrolló resistencia al DDT[2] y hoy en día es resistente a muchos insecticidas sintéticos utilizados en la agricultura contra este insecto.[1] Es más, también ostenta ser la primera especie en desarrollar resistencia a toxinas de B. thuringiensis.[3][21][22][23][24][25][26][27][28]
Genómica, evolución y adaptación
Muchos herbívoros están adaptados a alimentarse de un determinado grupo de plantas que se encuentran genética y bioquímicamente relacionados, y P. xylostella no es una excepción. Sin embargo los mecanismos que determinan esa adaptación evolutiva a ese grupo de plantas relacionado aún no se conocen bien. En su trabajo, You et al (2013)[9] tratan de profundizar en el conocimiento de la herbivoría y la detoxificación así como de dilucidar cuáles son las adaptaciones que han permitido a P. xylostella convertirse en una plaga a escala mundial. Para ello secuenciaron el genoma de P. xylostella y analizaron su estructura comparando con otras especies de lepidópteros.
En primer lugar predijeron que el número de genes era de unos 18.071 codificantes para proteínas y otros 781 de ARN's no codificantes. De todos ellos, 1.683[genes eran específicos de los lepidópteros y otros 1.412 eran específicos de P. xylostella. El número de genes específicos de especie en P. xylostella era superior que en otras dos especies como Danaus plexippus (mariposa monarca, con 1.184 genes específicos de especie) y Bombyx mori (gusano de seda, con 463 genes específicos de especie). Esos 1.412 genes específicos de P. xylostella están, en su mayoría, implicados en rutas esenciales para el procesamiento de información ambiental, en replicación y/o reparación cromosómica, regulación transcripcional y metabolismo de carbohidratos y proteínas, lo que sugiere que P. xylostella tiene una capacidad intrínseca (genes específicos) de respuesta rápida a estrés ambiental y a daño genético.
Al realizar un análisis filogenético estimaron la época de divergencia de los diferentes órdenes de insectos hace unos 265-332 millones de años atrás,[9] época muy próxima a la época de divergencia entre monocotiledóneas y dicotiledóneas (304 millones de años atrás)[29] y consistente con la hipótesis de que insectos herbívoros y sus plantas hospedadoras habrían coevolucionado desde entonces. En este sentido, P. xylostella se habría convertido en huésped específico de crucíferas (familia, Brassicaceae) tras la divergencia de dicha familia de la familia Caricaceae hace unos 54-90 millones de años atrás,[30] poco después de la divergencia de P. xylostella de D. plexippus y de B. mori hace unos 124 millones de años atrás (fecha estimada por You et al).[9]
Existe un conjunto de 354 genes que están preferencialmente expresados en estadios larvales, siendo muchos de ellos los que posibilitan esa adaptación específica de P. xylostella a las crucíferas. Entre ellos se encuentran:[9]
- Genes implicados en el metabolismo del sulfato, como los que codifican para glucosinolato sulfatasas (GSS1 y GSS2),[9] que permiten a P. xylostella alimentarse de un amplio número de crucíferas, ya que transforman el glucosinolato (compuesto de defensa de la planta) en desulfoglucosinolato (impidiendo la formación de productos tóxicos de hidrólisis).[31]
- Genes que codifican para enzimas digestivas. P. xylostella presenta un conjunto de enzimas digestivas similar a otra especie polifágica como es Helicoverpa armigera. Dentro de este conjunto de genes hay numerosos genes que codifican para serín proteasas. Cada planta hospedadora intentará defenderse de P. xylostella secretando inhibidores enzimáticos para estas serín proteasas digestivas, pero P. xylostella puede eludir dichos inhibidores puesto que presenta en su genoma un número mayor de genes para serín proteasas que inhibidores tiene la planta para éstas, por lo que siempre puede disponer de serín proteasas funcionales que no estén afectadas por los inhibidores de la planta hospedadora.
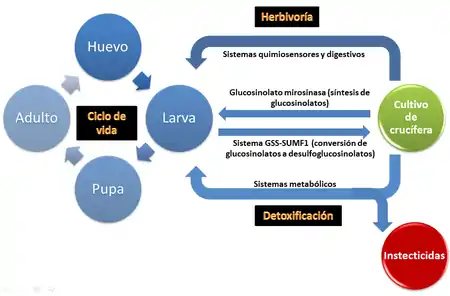
- Genes relacionados con el rango de plantas que puede hospedar. Dentro de este conjunto de genes se diferencian dos grupos:
- Genes encargados de la percepción de señales químicas provenientes de la planta hospedadora. Son cinco familias de genes que están relacionadas con la preferencia de ingesta en estadio larval y con la selección de plantas hospedadoras en adultos: receptores de odorantes (OR's), proteínas de unión a odorantes (OBP's), receptores gustativos (GR's), receptores ionotrópicos (IR's) y proteínas quimiosensoras (CSP's). De las cinco familias de genes se observa que existe expansión en concreto de los genes de la familia de los OR's, lo que va a conferir a P. xylostella un alto potencial de Adaptación biológica a señales químicas provenientes de las plantas hospedadoras.
- Genes de defensa contra compuestos secundarios de la planta. Son genes implicados en vías de detoxificación de compuestos de defensa de la planta que, a su vez, le van a ser útlies a P. xylostella para desarrollar tolerancia o resistencia a insecticidas (compuestos xenobióticos). Existe un mayor número de genes relacionados con la resistencia a insecticidas en P. xylostella que en B. mori (seguramente debido a que B. mori es una especie monofágica que ha tenido poca exposición a pesticidas). Muchos de los genes implicados en detoxificación van a estar duplicados en el genoma de P. xylostella, como ocurre en la mayoría de los genes que codifican para transportadores tipo ATP-binding cassette (transportadores ABC). Los genes de la familia de transportadores ABC son los más expandidos en el genoma de P. xylostella dentro de las familias de genes implicadas en vías de detoxificación comparando con el genoma de B. mori. Es más, en cepas resistentes a insecticidas los niveles de expresión de transportadores ABC están aumentados. Otras familias de genes implicadas en vías de detoxificación son los de las Citocromo P450 monooxigenasas (P450's), Glutatión-S-transferasas (GST's) y Carboxilesterasas (COE's). Dentro de las cuatro familias de genes implicadas en vías de detoxificación se observa un gran polimorfismo; por ejemplo, en genes de COE's hay un gran número de SNP's que conducen a cambios en la especificidad del sustrato y en la actividad catalítica. En general se observa un mayor número de transposones en genes involucrados en vías de detoxificación metabólica sensible a estrés externo (una tendencia general en lepidópteros).
En conclusión, la expansión de familias de genes implicados en percepción y detoxificación, así como el alto grado de variabilidad en dichos genes permiten a P. xylostella disponer de una batería de genes y alelos adaptados a las diversas y ubicuas toxinas que las crucíferas, como plantas hospedadoras, han desarrollado frente a esta y otras especies, pero también subraya la rápida capacidad que tiene P. xylostella de desarrollar resistencia a insecticidas.[9]
Referencias
- Talekar, N.S. & Shelton, A.M (1993). «Biology, ecology, and management of the diamondback moth». Annual Review of Entomology 38: 275-301. doi:10.1146/annurev.en.38.010193.001423.
- Ankersmit, G.W (1953). «DDT-resistance in Plutella maculipennis (Curt.) (Lep.) in Java». Bulletin of Entomological Research 44: 421-425. doi:10.1017/S0007485300025530.
- Heckel, D.G., Gahan, L.J., Liu, Y.B. & Tabashnik, B.E (1999). «Genetic mapping of resistance to Bacillus thuringiensis toxins in diamondback moth using biphasic linkage analysis». Proceedings of the National Academy of Sciences 96: 8373-8377. doi:10.1073/pnas.96.15.8373.
- http://arctos.database.museum/name/Plutella%20xylostella
- http://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi
- Wei, Shu-Jun; Shi, Bao-Cai; Gong, Ya-Jun; Jin, Gui-Hua; Chen, Xue-Xin; Meng, Xiang-Feng (2013). «Genetic Structure and Demographic History Reveal Migration of the Diamondback Moth Plutella xylostella (Lepidoptera: Plutellidae) from the Southern to Northern Regions of China». PLoS ONE 8 (4). doi:10.1371/journal.pone.0059654.
- Kfir, R. (1998). «Origin of the diamondback moth (Lepidoptera: Plutellidae)». Annals of the Entomological Society of America 91: 164-167. Archivado desde el original el 3 de marzo de 2014. Consultado el 2 de marzo de 2014.
- Hardy J. E. (1938). «Plutella maculipennis, (Curt.), its natural and biological control in England». Bulletin of Entomological Research 29: 343-372. doi:10.1017/S0007485300026274.
- You, M. et al (2013). «A heterozygous moth genome provides insights into herbivory and detoxification». Nature genetics 45 (2): 220-225. doi:10.1038/ng.2524.
- Xing, K. et al. Evidence of long distance migration of diamondback moth (DBM) Plutella xylostella: a review
- Yang, J. et al. Insight into the Migration Routes of Plutella xylostella in China Using mtCOI and ISSR Markers
- Capinera, J. L. (2000). Diamondback Moth, Plutella xylostella (Linnaeus)(Insecta: Lepidoptera: Plutellidae). University of Florida/The Institute of Food and Agricultural Sciences (IFAS).
- Dosdall, L. M., Mason, P. G., Olfert, O., Kaminski, L., & Keddie, B. A. (Noviembre de 2001). «The origins of infestations of diamondback moth, Plutella xylostella (L.), in canola in western Canada». Proceedings of the Fourth International Workshop on the Management of Diamondback Moth and Other Crucifer Pests. Melbourne, Australia. pp. 95-100.
- Nguyen, C., Bahar, M. H., Baker, G., & Andrew, N. R. (2014). «Thermal Tolerance Limits of Diamondback Moth in Ramping and Plunging Assays». PloS one 9 (1): e87535. doi:10.1371/journal.pone.0087535.
- Honda, K. I. (Diciembre de 1992). «Hibernation and migration of diamondback moth in northern Japan». Diamondback Moth and Other Crucifer Pests: Proceedings of the Second International Workshop. AVRDC, Taiwan. pp. 43-50.
- Walker, M. K., Davidson, M. M., Wallace, A. R., & Walker, G. P. (Abril de 2001). «Monitoring of diamondback moth in a cold-winter climate, South Island, New Zealand». The Sixth International Workshop on Management of the Diamondback Moth and Other Crucifer Insect Pests. Nakon Pathom, Tailandia. pp. 51-57.
- Dosdall LM (1994). «Evidence for successful overwintering of diamondback moth, Plutella xylostella (L.) (Lepidoptera: Plutellidae), in Alberta». The Canadian Entomologist 126: 183-185. doi:10.4039/Ent126183-1.
- Badenes-Pérez, F., Riechelt, R.M., Gershenzon, J. & Hechel, D.G. (Abril de 2001). «Importance of glucosinolates in determining diamondback moth preference and host range». The Sixth International Workshop on Management of the Diamondback Moth and Other Crucifer Insect Pests. Nakon Pathom, Tailandia. pp. 63-66.
- Furlong, M. J., Wright, D. J., & Dosdall, L. M. (2013). «Diamondback moth ecology and management: problems, progress, and prospects». Annual review of entomology 58: 517-541. doi:10.1146/annurev-ento-120811-153605.
- Zalucki, M.P. et al (2012). «Estimating the economic cost of one of the world’s major insect pests, Plutella xylostella (Lepidoptera: Plutellidae): just how long is a piece of string?». Journal of Economic Entomology 105: 1115-1129. doi:10.1603/EC12107.
- Tabashnik, B. E. et al (1996). «Cross-resistance of the diamondback moth indicates altered interactions with domain II of Bacillus thuringiensis toxins». Applied and environmental microbiology 62: 2839-2844.
- Meyer, S. K., Tabashnik, B. E., Liu, Y. B., Wirth, M. C., & Federici, B. A. (2001). «Cyt1A from Bacillus thuringiensis lacks toxicity to susceptible and resistant larvae of diamondback moth (Plutella xylostella) and pink bollworm (Pectinophora gossypiella)». Applied and environmental microbiology 67: 462-463.
- Zhao, J. Z., LI, Y. X., Collins, H. L., Cao, J., Earle, E. D., & Shelton, A. M. (2001). «Different cross-resistance patterns in the diamondback moth (Lepidoptera: Plutellidae) resistant to Bacillus thuringiensis toxin Cry1C». Journal of economic entomology 94: 1547-1552. doi:10.1603/0022-0493-94.6.1547.
- Hernández-Martínez, P., Hernández-Rodríguez, C. S., Krishnan, V., Crickmore, N., Escriche, B., & Ferré, J. (2012). «Lack of Cry1Fa binding to the midgut brush border membrane in a resistant colony of Plutella xylostella moths with a mutation in the ABCC2 locus». Applied and environmental microbiology 78: 6759-6761.
- Baxter, S. W., et al (2011). «Parallel evolution of Bacillus thuringiensis toxin resistance in Lepidoptera». Genetics 189: 675-679. doi:10.1534/genetics.111.130971.
- Shelton, A. M., et al (1993). «Resistance of diamondback moth (Lepidoptera: Plutellidae) to Bacillus thuringiensis subspecies in the field». Journal of Economic Entomology 86: 697-705.
- Ferré, J., Real, M. D., Van Rie, J., Jansens, S., & Peferoen, M. (1991). «Resistance to the Bacillus thuringiensis bioinsecticide in a field population of Plutella xylostella is due to a change in a midgut membrane receptor». Proceedings of the National Academy of Sciences 88: 5119-5123. doi:10.1073/pnas.88.12.5119.
- Tabashnik, B. E., Finson, N., Groeters, F. R., Moar, W. J., Johnson, M. W., Luo, K., & Adang, M. J. (1994). «Reversal of resistance to Bacillus thuringiensis in Plutella xylostella». Proceedings of the National Academy of Sciences 91: 4120-4124.
- Zimmer, A., Lang, D., Richardt, S., Frank, W., Reski, R., & Rensing, S. A. (2007). «Dating the early evolution of plants: detection and molecular clock analyses of orthologs». Molecular Genetics and Genomics 278 (4): 393-402. doi:10.1007/s00438-007-0257-6.
- Wang, X. et al' (2011). «The genome of the mesopolyploid crop species Brassica rapa». Nature genetics 43 (10): 1035-1039. doi:10.1038/ng.919.
- Ratzka, A., Vogel, H., Kliebenstein, D. J., Mitchell-Olds, T., & Kroymann, J. (2002). «Disarming the mustard oil bomb». Proceedings of the National Academy of Sciences 99 (17): 11223-11228. doi:10.1073/pnas.172112899.
Enlaces externos
- Cornell University. Palomilla Dorso de Diamante (DDM)
- Instituto Nacional de Investigaciones Forestales de México. Manejo Integrado de la Palomilla Dorso de Diamante Plutella xylostella (L.)
| Control de autoridades |
|
|---|
 Datos: Q1050343
Datos: Q1050343 Multimedia: Plutella xylostella / Q1050343
Multimedia: Plutella xylostella / Q1050343 Especies: Plutella xylostella
Especies: Plutella xylostella