Raza y genética
Los investigadores han investigado la relación entre raza y genética como parte de los esfuerzos para comprender cómo la biología puede contribuir o no a la categorización racial humana.
| Genética humana | ||
|---|---|---|
| Subtemas |
| |
Muchas construcciones de raza están asociadas con rasgos fenotípicos y ascendencia geográfica, y estudiosos como Carlos Linneo han propuesto modelos científicos para la organización de la raza desde al menos el siglo XVIII. Tras el descubrimiento de la genética mendeliana y el mapeo del genoma humano, las preguntas sobre la biología de la raza a menudo se han enmarcado en términos de genética.[1] Se ha empleado una amplia gama de métodos de investigación para examinar los patrones de variación humana y sus relaciones con la ascendencia y los grupos raciales, incluidos estudios de rasgos individuales,[2] estudios de grandes poblaciones y grupos genéticos[3] y estudios de factores genéticos de riesgo de enfermedad.[4]
La investigación sobre la raza y la genética también ha sido criticada por surgir o contribuir al racismo científico. Algunos han interpretado los estudios genéticos de rasgos y poblaciones como evidencia para justificar las desigualdades sociales asociadas con la raza,[5] a pesar de que se ha demostrado que los patrones de variación humana son principalmente clinales, siendo el código genético humano aproximadamente un 99,9% idéntico entre individuos y sin límites claros entre los grupos.[6][1]
Existe un debate científico en curso sobre la definición y el significado de raza en la investigación genética y biomédica. Algunos investigadores sostienen que la raza puede actuar como un "proxy" de la ascendencia genética porque los individuos de la misma categoría racial pueden compartir una ascendencia común, pero otros abogan por distinguir entre la biología y los factores sociales, políticos, culturales y económicos que contribuyen a la raza, como se entiende más comúnmente.[7]
Variación genética
La variación genética surge de mutaciones, de la selección natural, de la migración entre poblaciones (flujo genético) y de la reorganización de genes a través de la reproducción sexual.[8] Las mutaciones conducen a un cambio en la estructura del ADN, ya que se reorganiza el orden de las bases. Como resultado, se codifican diferentes proteínas polipeptídicas. Algunas mutaciones pueden ser positivas y pueden ayudar al individuo a sobrevivir de manera más eficaz en su entorno. La variación se contrarresta mediante la selección natural y la deriva genética; nótese también el efecto fundador, cuando un pequeño número de fundadores iniciales establece una población que, por tanto, comienza con un grado correspondientemente pequeño de variación genética.[9] La herencia epigenética implica cambios hereditarios en el fenotipo (apariencia) o la expresión génica, causados por mecanismos distintos a los cambios en la secuencia del ADN.[10]
Los fenotipos humanos son altamente poligénicos (dependen de la interacción de muchos genes) y están influenciados tanto por el medio ambiente como por la genética.
La diversidad de nucleótidos se basa en mutaciones únicas, polimorfismos de nucleótido único (SNP). La diversidad de nucleótidos entre humanos es de aproximadamente 0,1 % (una diferencia por mil nucleótidos entre dos humanos elegidos al azar). Esto equivale a aproximadamente tres millones de SNP (ya que el genoma humano tiene alrededor de tres mil millones de nucleótidos). Se estima que hay diez millones de SNP en la población humana.[11]
La investigación ha demostrado que la variación no SNP (estructural) explica más variación genética humana que la diversidad de un solo nucleótido. La variación estructural incluye la variación del número de copias y los resultados de eliminaciones, inversiones, inserciones y duplicaciones. Se estima que aproximadamente del 0,4 al 0,6 por ciento de los genomas de personas no relacionadas difieren.[12][13]
¿Existe una base genética para la raza?
Gran parte de la investigación científica se ha organizado en torno a la cuestión de si existe o no una base genética para la raza. Según Luigi Luca Cavalli-Sforza,
Desde un punto de vista científico, el concepto de raza no ha logrado obtener ningún consenso; ninguno es probable, dada la variación gradual que existe. Se puede objetar que los estereotipos raciales tienen una coherencia que permite incluso al lego clasificar a los individuos. Sin embargo, los principales estereotipos, todos basados en el color de la piel, el color y la forma del cabello y los rasgos faciales, reflejan diferencias superficiales que no son confirmadas por un análisis más profundo con rasgos genéticos más confiables y cuyo origen se remonta a la evolución reciente principalmente bajo el efecto del clima y quizás selección sexual.[14][15][16][17][18][19]
Métodos de búsqueda
Los científicos que investigan la variación humana han utilizado una serie de métodos para caracterizar cómo varían las diferentes poblaciones.
Estudios iniciales de rasgos, proteínas y genes
Los primeros intentos de clasificación racial midieron los rasgos de la superficie, particularmente el color de la piel, el color y la textura del cabello, el color de los ojos y el tamaño y la forma de la cabeza. (Las mediciones de este último a través de la craneometría fueron desacreditadas repetidamente a fines del siglo XIX y mediados del XX debido a la falta de correlación de los rasgos fenotípicos con la categorización racial.[20]) En realidad, la adaptación biológica juega el papel más importante en estas características corporales y tipo de piel. Un puñado relativo de genes explica los factores heredados que dan forma a la apariencia de una persona.[21][22] Se estima que los seres humanos tienen entre 19 000 y 20 000 genes codificadores de proteínas humanas.[23] Richard Sturm y David Duffy describen 11 genes que afectan la pigmentación de la piel y explican la mayoría de las variaciones en el color de la piel humana, las más importantes de las cuales son MC1R, ASIP, OCA2 y TYR.[24] Existe evidencia de que hasta 16 genes diferentes podrían ser responsables del color de ojos en humanos; sin embargo, los dos genes principales asociados con la variación del color de los ojos son OCA2 y HERC2, y ambos están localizados en el cromosoma 15.[25]
Análisis de proteínas sanguíneas y genética entre grupos
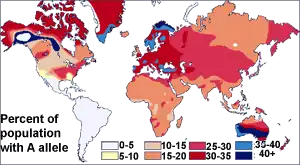
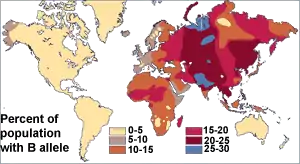
Antes del descubrimiento del ADN, los científicos usaban proteínas sanguíneas (los sistemas de grupos sanguíneos humanos) para estudiar la variación genética humana. La investigación de Ludwik y Hanka Herschfeld durante la Primera Guerra Mundial encontró que la incidencia de los grupos sanguíneos A y B difería según la región; por ejemplo, entre los europeos, el 15 por ciento eran del grupo B y el 40 por ciento del grupo A. Los europeos del este y los rusos tenían una mayor incidencia del grupo B; la gente de la India tuvo la mayor incidencia. Los Herschfelds concluyeron que los humanos comprenden dos "razas bioquímicas", que se originan por separado. Se planteó la hipótesis de que estas dos razas se mezclaron más tarde, lo que resultó en los patrones de los grupos A y B. Esta fue una de las primeras teorías de las diferencias raciales en incluir la idea de que la variación humana no se correlaciona con la variación genética. Se esperaba que los grupos con proporciones similares de grupos sanguíneos estuvieran más estrechamente relacionados, pero en cambio, a menudo se encontró que los grupos separados por grandes distancias (como los de Madagascar y Rusia), tenían incidencias similares.[26] Más tarde se descubrió que el sistema de grupos sanguíneos ABO no solo es común a los humanos, sino que se comparte con otros primates,[27] y probablemente es anterior a todos los grupos humanos.[28]
En 1972, Richard Lewontin realizó un análisis estadístico FST utilizando 17 marcadores (incluidas las proteínas del grupo sanguíneo). Encontró que la mayoría de las diferencias genéticas entre humanos (85,4 por ciento) se encontraron dentro de una población, el 8,3 por ciento se encontró entre poblaciones dentro de una raza y el 6,3 por ciento se encontró para diferenciar razas (caucásicos, africanos, mongoloides, aborígenes del sur de Asia, amerindios, aborígenes australianos y provenientes de Oceanía en su estudio). Desde entonces, otros análisis han encontrado valores de FST del 6 al 10 por ciento entre grupos humanos continentales, del 5 al 15 por ciento entre diferentes poblaciones en el mismo continente y del 75 al 85 por ciento dentro de las poblaciones.[29][30][31][32][33] Este punto de vista ha sido confirmado por la Asociación Americana de Antropología y la Asociación Americana de Antropólogos Físicos desde entonces.[34]
Críticas al análisis de proteínas en sangre
Si bien reconoce la observación de Lewontin de que los humanos son genéticamente homogéneos, A. W. F. Edwards en su artículo de 2003 Human Genetic Diversity: Lewontin's Falacy argumentó que la información que distingue a las poblaciones entre sí está oculta en la estructura de correlación de las frecuencias alélicas, lo que hace posible clasificar a los individuos utilizando métodos matemáticos. Edwards argumentó que incluso si la probabilidad de clasificar erróneamente a un individuo en función de un solo marcador genético es tan alta como el 30 por ciento (como informó Lewontin en 1972), la probabilidad de clasificación errónea se acerca a cero si se estudian simultáneamente suficientes marcadores genéticos. Edwards vio el argumento de Lewontin como basado en una postura política, negando las diferencias biológicas para defender la igualdad social.[35] El artículo de Edwards se reimprime, comentado por expertos como Noah Rosenberg y se le da más contexto en una entrevista con el filósofo de la ciencia Rasmus Grønfeldt Winther en una antología reciente.[36]
Edwards criticó el artículo de Lewontin, ya que tomó 17 rasgos diferentes y los analizó de forma independiente, sin mirarlos junto con ninguna otra proteína. Por lo tanto, hubiera sido bastante conveniente para Lewontin llegar a la conclusión de que el naturalismo racial no es sostenible, según su argumento.[37] Sesardic también fortaleció la visión de Edwards, ya que usó una ilustración que se refiere a cuadrados y triángulos, y mostró que si miras un rasgo de forma aislada, lo más probable es que sea un mal predicador de a qué grupo pertenece el individuo.[38] En contraste, en un artículo de 2014, reimpreso en el volumen de Edwards Cambridge University Press de 2018, Rasmus Grønfeldt Winther sostiene que "La falacia de Lewontin" es efectivamente un nombre inapropiado, ya que en realidad hay dos conjuntos diferentes de métodos y preguntas en juego al estudiar la estructura poblacional genómica de nuestra especie: "partición de varianza" y "análisis de agrupamiento". Según Winther, son "las dos caras de la misma moneda matemática" y ninguna "implica necesariamente nada sobre la realidad de los grupos humanos".[39]
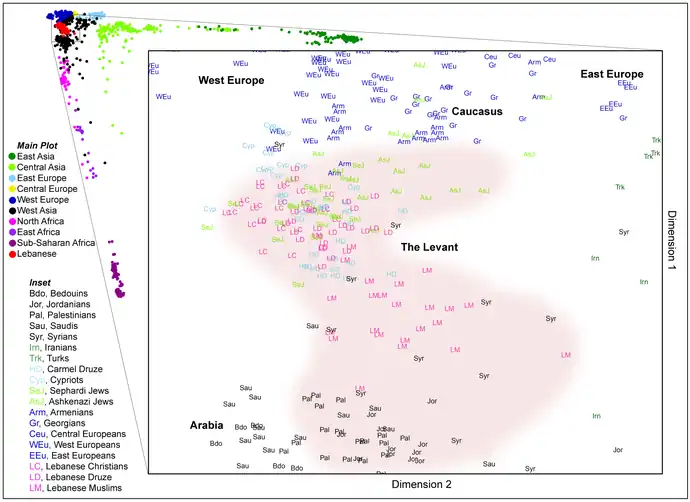
Estudios actuales de genética de poblaciones
Los investigadores actualmente usan pruebas genéticas, que pueden involucrar cientos (o miles) de marcadores genéticos o todo el genoma.
Estructura
Existen varios métodos para examinar y cuantificar subgrupos genéticos, incluido el análisis de conglomerados y el análisis de componentes principales. Los marcadores genéticos de los individuos se examinan para encontrar la estructura genética de una población. Si bien los subgrupos se superponen cuando se examinan variantes de un solo marcador, cuando se examinan varios marcadores, los diferentes subgrupos tienen una estructura genética promedio diferente. Un individuo puede describirse como perteneciente a varios subgrupos. Estos subgrupos pueden ser más o menos distintos, dependiendo de cuánta superposición haya con otros subgrupos.[40]
En el análisis de conglomerados, el número de conglomerados para buscar K se determina de antemano; lo distintos que son los grupos varía.
Los resultados obtenidos de los análisis de conglomerados dependen de varios factores:
- Un gran número de marcadores genéticos estudiados facilita la búsqueda de grupos distintos.[41]
- Algunos marcadores genéticos varían más que otros, por lo que se requieren menos para encontrar grupos distintos.[42] Los marcadores informativos de ascendencia exhiben frecuencias sustancialmente diferentes entre poblaciones de diferentes regiones geográficas. Con AIM, los científicos pueden determinar el continente de origen ancestral de una persona basándose únicamente en su ADN. Los AIM también se pueden utilizar para determinar las proporciones de mezcla de una persona.[43]
- Cuantos más individuos se estudien, más fácil será detectar grupos distintos (se reduce el ruido estadístico).
- La baja variación genética hace que sea más difícil encontrar grupos distintos. Una mayor distancia geográfica generalmente aumenta la variación genética, lo que facilita la identificación de grupos.[44]
- Se observa una estructura de grupo similar con diferentes marcadores genéticos cuando el número de marcadores genéticos incluidos es suficientemente grande. La estructura de agrupamiento obtenida con diferentes técnicas estadísticas es similar. Se encuentra una estructura de conglomerados similar en la muestra original con una submuestra de la muestra original.[45]
Se han publicado estudios recientes utilizando un número creciente de marcadores genéticos.[42][45][46][47][48][49]
Distancia
La distancia genética es la divergencia genética entre especies o poblaciones de una especie. Puede comparar la similitud genética de especies relacionadas, como humanos y chimpancés. Dentro de una especie, la distancia genética mide la divergencia entre subgrupos. La distancia genética se correlaciona significativamente con la distancia geográfica entre poblaciones, un fenómeno conocido a veces como "aislamiento por distancia".[50] La distancia genética puede ser el resultado de límites físicos que restringen el flujo de genes, como islas, desiertos, montañas o bosques. La distancia genética se mide mediante el índice de fijación (FST). El FST es la correlación de alelos elegidos al azar en un subgrupo con una población más grande. A menudo se expresa como una proporción de la diversidad genética. Esta comparación de la variabilidad genética dentro (y entre) poblaciones se utiliza en genética de poblaciones. Los valores van de 0 a 1; cero indica que las dos poblaciones se cruzan libremente y uno indicaría que dos poblaciones están separadas.
Muchos estudios sitúan la distancia FST media entre razas humanas en aproximadamente 0,125. Henry Harpending argumentó que este valor implica a escala mundial un "parentesco entre dos individuos de la misma población humana es equivalente al parentesco entre abuelos y nietos o entre medio hermanos". De hecho, las fórmulas derivadas del artículo de Harpending implican que dos individuos no emparentados de la misma raza tienen un coeficiente de parentesco más alto (0.125) que un individuo y su medio hermano de raza mixta (0.109).[51]
Críticas a FST
Aunque se reconoce que el FST sigue siendo útil, varios científicos han escrito sobre otros enfoques para caracterizar la variación genética humana.[52][53][54] Long y Kittles afirmaron que el FST no logró identificar variaciones importantes y que cuando el análisis incluye solo humanos, FST = 0.119, pero agregar chimpancés lo aumenta solo a FST = 0.183. Se ha argumentado que una estimación de FST de 0,10-0,15 no descarta una base genética para las diferencias fenotípicas entre grupos y que una estimación de FST baja implica poco sobre el grado en que los genes contribuyen a las diferencias entre grupos. Pearse y Crandall escribieron que las cifras de FST no pueden distinguir entre una situación de alta migración entre poblaciones con un tiempo de divergencia prolongado y una de historia compartida relativamente reciente pero sin flujo de genes en curso. En su artículo de 2015, Keith Hunley, Graciela Cabana y Jeffrey Long (que habían criticado previamente la metodología estadística de Lewontin con Rick Kittles[34]) recalculan la distribución de la diversidad humana utilizando un modelo más complejo que Lewontin y sus sucesores.
En resumen, estamos de acuerdo con la conclusión de Lewontin de que las clasificaciones raciales basadas en Occidente no tienen importancia taxonómica, y esperamos que esta investigación, que tiene en cuenta nuestra comprensión actual de la estructura de la diversidad humana, sitúe su hallazgo fundamental en base evolutiva.[55]
Los antropólogos (como C. Loring Brace),[56] filósofo Jonathan Kaplan y el genetista Joseph L. Graves[57] han argumentado que si bien es posible encontrar variaciones biológicas y genéticas que se correspondan aproximadamente con la raza, esto es cierto para casi todas las poblaciones geográficamente distinta: la estructura de grupos de datos genéticos depende de las hipótesis iniciales del investigador y de las poblaciones muestreadas. Cuando se toman muestras de grupos continentales, los racimos se vuelven continentales; con otros patrones de muestreo, los conglomerados serían diferentes. Weiss y Fullerton señalan que si se muestrearan solo islandeses, mayas y maoríes, se formarían tres grupos distintos; todas las demás poblaciones estarían compuestas por mezclas genéticas de material maorí, islandés y maya.[58] Por lo tanto, Kaplan concluye que, si bien las diferencias en frecuencias alélicas particulares se pueden usar para identificar poblaciones que se corresponden libremente con las categorías raciales comunes en el discurso social occidental, las diferencias no tienen más importancia biológica que las diferencias encontradas entre cualquier población humana (por ejemplo, el Español y portugués).[59]
Análisis histórico y geográfico
La estructura genética de la población actual no implica que los diferentes grupos o componentes indiquen solo un hogar ancestral por grupo; por ejemplo, un grupo genético en los Estados Unidos comprende hispanos con ascendencia europea, nativa americana y africana.[41]
Los análisis geográficos intentan identificar los lugares de origen, su importancia relativa y las posibles causas de variación genética en un área. Los resultados se pueden presentar como mapas que muestran la variación genética. Se ha argumentado que si se investigan las variaciones genéticas, a menudo corresponden a migraciones de población debido a nuevas fuentes de alimentos, transporte mejorado o cambios en el poder político. Por ejemplo, en Europa, la dirección más significativa de variación genética corresponde a la expansión de la agricultura desde el Medio Oriente a Europa entre hace 10,000 y 6,000 años.[60] Este análisis geográfico funciona mejor en ausencia de migraciones rápidas y a gran escala recientes.
Los análisis históricos utilizan las diferencias en la variación genética (medidas por distancia genética) como un reloj molecular que indica la relación evolutiva de especies o grupos, y se pueden utilizar para crear árboles evolutivos que reconstruyen las separaciones de poblaciones.[60]
Los resultados de la investigación de la ascendencia genética se apoyan si están de acuerdo con los resultados de la investigación de otros campos, como la lingüística o la arqueología.[60] También se ha argumentado que existe una correspondencia entre las familias de lenguas encontradas en la investigación lingüística y el árbol de población. Por lo general, existen distancias genéticas más cortas entre poblaciones que utilizan lenguas de la misma familia lingüística. También se encuentran excepciones a esta regla, por ejemplo Sami, que están genéticamente asociados con poblaciones que hablan idiomas de otras familias lingüísticas. Los Sami hablan una lengua urálica, pero genéticamente son principalmente europeos. Se argumenta que esto es el resultado de la migración (y el mestizaje) con los europeos mientras se conserva su idioma original. También existe un acuerdo entre las fechas de investigación en arqueología y las calculadas utilizando la distancia genética.[42]
Estudios de autoidentificación
Se ha encontrado que si bien los grupos de marcadores genéticos estaban correlacionados con algunos conceptos tradicionales de raza, las correlaciones eran imperfectas e imprecisas debido a la naturaleza continua y superpuesta de la variación genética, y señalaron que la ascendencia, que puede determinarse con precisión, no es equivalente al concepto de raza.[61]
Un estudio de 2005 utilizó 326 marcadores genéticos para determinar los grupos genéticos. Los 3.636 sujetos, de Estados Unidos y Taiwán, se autoidentificaron como pertenecientes a grupos étnicos blancos, afroamericanos, asiáticos orientales o hispanos. El estudio encontró "una correspondencia casi perfecta entre el grupo genético y SIRE para los principales grupos étnicos que viven en los Estados Unidos, con una tasa de discrepancia de solo el 0,14 por ciento".[41] Se encontró un acuerdo "esencialmente perfecto" entre 51 poblaciones de origen autoidentificadas y la estructura genética de la población, utilizando 650.000 marcadores genéticos. La selección de marcadores genéticos informativos permitió una reducción a menos de 650, manteniendo una precisión casi total.[62]
La correspondencia entre los grupos genéticos en una población (como la población actual de EE. UU.) y la raza o los grupos étnicos autoidentificados no significa que dicho grupo (o grupo) corresponda a un solo grupo étnico. Los afroamericanos tienen una mezcla genética europea estimada del 20 al 25 por ciento; los hispanos tienen ascendencia europea, nativa americana y africana.[41] En Brasil ha habido una gran mezcla entre europeos, amerindios y africanos. Como resultado, las diferencias en el color de la piel dentro de la población no son graduales y existen asociaciones relativamente débiles entre la raza autoinformada y la ascendencia africana.[63][64] La autoclasificación etnoracial en los brasileños ciertamente no es aleatoria con respecto a la ascendencia individual del genoma, pero la fuerza de la asociación entre el fenotipo y la proporción mediana de ascendencia africana varía en gran medida entre la población.[65]
Crítica de los estudios y conglomerados de la distancia genética
Las distancias genéticas generalmente aumentan continuamente con la distancia geográfica, lo que hace que una línea divisoria sea arbitraria. Cualesquiera dos asentamientos vecinos exhibirán alguna diferencia genética entre sí, lo que podría definirse como una raza. Por tanto, los intentos de clasificar las razas imponen una discontinuidad artificial a un fenómeno natural. Esto explica por qué los estudios sobre la estructura genética de la población arrojan resultados variables, según la metodología.
Rosenberg ha argumentado, basándose en el análisis de conglomerados de las 52 poblaciones en el Panel de diversidad genética humana, que las poblaciones no siempre varían continuamente y que la estructura genética de una población es consistente si se incluyen suficientes marcadores genéticos (y sujetos).
El examen de la relación entre la distancia genética y geográfica respalda una visión en la que los conglomerados surgen no como un artefacto del esquema de muestreo, sino a partir de pequeños saltos discontinuos en la distancia genética para la mayoría de los pares de poblaciones en lados opuestos de las barreras geográficas, en comparación con la distancia genética. para parejas del mismo lado. Por lo tanto, el análisis del conjunto de datos de 993 locus corrobora nuestros resultados anteriores: si se usan suficientes marcadores con una muestra mundial lo suficientemente grande, los individuos pueden dividirse en grupos genéticos que coincidan con las principales subdivisiones geográficas del mundo, y algunos individuos de ubicaciones geográficas intermedias tienen membresía mixta en los clústeres que corresponden a regiones vecinas.
También escribieron, con respecto a un modelo con cinco grupos correspondientes a África, Eurasia (Europa, Medio Oriente y Asia Central /Meridional), Asia Oriental, Oceanía y América:
Para los pares de poblaciones del mismo conglomerado, a medida que aumenta la distancia geográfica, la distancia genética aumenta de manera lineal, consistente con una estructura poblacional clinal. Sin embargo, para pares de diferentes grupos, la distancia genética es generalmente mayor que la que existe entre pares dentro del grupo que tienen la misma distancia geográfica. Por ejemplo, las distancias genéticas para los pares de poblaciones con una población en Eurasia y la otra en el este de Asia son mayores que las de los pares a una distancia geográfica equivalente dentro de Eurasia o en el este de Asia. En términos generales, son estos pequeños saltos discontinuos en la distancia genética —a través de océanos, el Himalaya y el Sahara— los que proporcionan la base para la capacidad de la ESTRUCTURA para identificar agrupaciones que corresponden a regiones geográficas.[45]
Esto se aplica a las poblaciones en sus hogares ancestrales cuando las migraciones y el flujo de genes fueron lentos; las migraciones grandes y rápidas exhiben características diferentes. Afirmando,
...solo detectamos una diferenciación genética modesta entre diferentes lugares geográficos actuales dentro de cada grupo racial/étnico. Por lo tanto, la ascendencia geográfica antigua, que está altamente correlacionada con la raza/etnia autoidentificada, a diferencia de la residencia actual, es el principal determinante de la estructura genética en la población de Estados Unidos.[41]

El análisis de conglomerados ha sido criticado porque el número de conglomerados a buscar se decide de antemano, con diferentes valores posibles (aunque con diferentes grados de probabilidad).[66] El análisis de componentes principales no decide de antemano cuántos componentes buscar.[67]
El estudio de 2002 de Noah Rosenberg ejemplifica por qué los significados de estos agrupamientos son discutibles. El estudio muestra que en el análisis de conglomerados K = 5, los conglomerados genéticos se mapean aproximadamente en cada una de las cinco regiones geográficas principales. En 2005 se obtuvieron resultados similares en estudios adicionales[68]
Sin embargo, además de los cinco conglomerados supuestamente geográficos principales, un sexto grupo, los Kalash, un grupo étnico minoritario en Pakistán, comenzó a aparecer a partir de K = 6. El naturalista racial Nicholas Wade considera que los resultados "no tienen sentido genético o geográfico". Por lo tanto, se omiten en su libro A Troublesome Inheritance a favor del análisis de conglomerados K = 5.
Este sesgo, sin embargo, refleja cómo la investigación es inherentemente defectuosa. La población de muestra se elige teniendo en cuenta la representación geográfica y los conceptos populares de raza, en lugar de tener en cuenta la diversidad genética dentro de las diferentes regiones geográficas. Los Kalash no encajaban en el patrón general porque había sido una población genéticamente aislada que se reflejó en este estudio. Potencialmente numerosos grupos genéticamente modificados, como los Sentineleses no contactados, no están representados en el estudio.
Crítica de los marcadores informativos de ascendencia
Los marcadores informativos de ascendencia (AIM) son una tecnología de rastreo de genealogía que ha sido objeto de muchas críticas debido a su dependencia de las poblaciones de referencia. Se ha descrito cómo la tecnología contemporánea permite rastrear el linaje ancestral, pero solo a lo largo de las líneas de una línea materna y una línea paterna. Es decir, de 64 tatarabuelos en total, solo se identifica uno de cada padre, lo que implica que los otros 62 antepasados se ignoran en los esfuerzos de rastreo.[69] Además, las "poblaciones de referencia" utilizadas como marcadores para la pertenencia a un grupo particular se designan de forma arbitraria y contemporánea. El uso de poblaciones que actualmente residen en lugares determinados como referencias para ciertas razas y grupos étnicos no es confiable debido a los cambios demográficos que se han producido durante muchos siglos en esos lugares. Además, los marcadores informativos de ascendencia se comparten ampliamente entre toda la población humana, lo que se prueba es su frecuencia, no su mera ausencia/presencia. Por lo tanto, debe establecerse un umbral de frecuencia relativa. Los criterios para establecer dichos umbrales son un secreto comercial de las empresas que comercializan las pruebas. Por tanto, no se puede decir nada concluyente sobre si son apropiados. Los resultados de los AIM son extremadamente sensibles a dónde se establece esta barra.[70] Dado que muchos rasgos genéticos se encuentran muy similares entre muchas poblaciones diferentes, la tasa de frecuencia que se considera suficiente para formar parte de una población de referencia es muy importante. Esto también puede conducir a errores, dado que muchas poblaciones pueden compartir los mismos patrones, si no exactamente los mismos genes.
Esto significa que alguien de Bulgaria cuyos antepasados se remontan al siglo XV podría (y en algún momento lo hace) mapear como en parte 'nativo americano'.
Esto sucede porque los AIM se basan en una suposición de '100% de pureza' de las poblaciones de referencia. Es decir, asumen que un patrón de rasgos sería idealmente condición necesaria y suficiente para asignar un individuo a una población ancestral de referencia.
Raza, genética y medicina
Existen ciertas diferencias estadísticas entre grupos raciales en cuanto a la susceptibilidad a ciertas enfermedades.[71] Los genes cambian en respuesta a enfermedades locales; por ejemplo, las personas que son negativas tienden a tener una mayor resistencia a la malaria. El fenotipo negativo es muy frecuente en África central y la frecuencia disminuye con la distancia a África central, con frecuencias más altas en poblaciones globales con altos grados de inmigración africana reciente. Esto sugiere que el genotipo negativo evolucionó en África subsahariana y posteriormente fue seleccionado positivamente para la zona endémica de malaria. Varias condiciones genéticas prevalentes en áreas endémicas de malaria pueden proporcionar resistencia genética a la malaria, incluida la anemia de células falciformes, talasemias y glucosa-6-fosfato deshidrogenasa. La fibrosis quística es la enfermedad autosómica recesiva limitante de la vida más común entre las personas de ascendencia europea; se ha cuestionado una hipotética ventaja heterocigótica, que proporciona resistencia a enfermedades que antes eran comunes en Europa.[72] Los científicos Michael Yudell, Dorothy Roberts, Rob DeSalle y Sarah Tishkoff argumentaron que el uso de estas asociaciones en la práctica de la medicina ha llevado a los médicos a pasar por alto o identificar erróneamente la enfermedad.
Por ejemplo, las hemoglobinopatías pueden diagnosticarse erróneamente debido a la identificación de la anemia drepanocítica como una enfermedad 'negra' y talasemia como enfermedad 'mediterránea'. La fibrosis quística está infradiagnosticada en poblaciones de ascendencia africana, porque se la considera una enfermedad 'blanca'.[73]
La información sobre la población de origen de una persona puede ayudar en el diagnóstico y las respuestas adversas a los medicamentos pueden variar según el grupo.[42] Debido a la correlación entre la raza autoidentificada y los grupos genéticos, los tratamientos médicos influenciados por la genética tienen diferentes tasas de éxito entre los grupos raciales autodefinidos.[74] Por esta razón, algunos médicos consideran la raza del paciente al elegir el tratamiento más eficaz,[75] y algunos medicamentos se comercializan con instrucciones específicas de la raza.[76] Se ha argumentado que debido a la variación genética dentro de los grupos raciales, cuando
...finalmente sea factible y esté disponible, la evaluación genética individual de genes relevantes probablemente resultará más útil que la raza en la toma de decisiones médicas.
Sin embargo, la raza sigue siendo un factor al examinar grupos (como la investigación epidemiológica).[61] Algunos médicos y científicos, como el genetista Neil Risch, argumentan que el uso de la raza autoidentificada como un sustituto de la ascendencia es necesario para poder obtener una muestra suficientemente amplia de diferentes poblaciones ancestrales y, a su vez, para poder brindar una atención médica personalizada a las necesidades de los grupos minoritarios.[77]
Uso en revistas científicas
Algunas revistas científicas han abordado errores metodológicos anteriores al exigir un escrutinio más riguroso de las variables de población. Desde 2000, Nature Genetics exige a sus autores que "expliquen por qué hacen uso de grupos étnicos o poblaciones particulares y cómo se logró la clasificación". Los editores de Nature Genetics dicen que "[ellos] esperan que esto genere conciencia e inspire diseños más rigurosos de estudios genéticos y epidemiológicos".[78]
Interacciones gen-ambiente
Se ha argumentado que la raza autoidentificada es de mayor uso en medicina, ya que se correlaciona fuertemente con exposomas relacionados con el riesgo que son potencialmente heredables cuando se incorporan al epigenoma. Resumen la evidencia del vínculo entre la discriminación racial y los resultados de salud debido a la peor calidad de los alimentos, el acceso a la atención médica, las condiciones de vivienda, la educación, el acceso a la información, la exposición a agentes infecciosos y sustancias tóxicas y la escasez de materiales. También citan evidencia de que este proceso puede funcionar positivamente; por ejemplo, la ventaja psicológica de percibirse a sí mismo en la cima de una jerarquía social está relacionada con una mejor salud. Sin embargo, advierten que los efectos de la discriminación no ofrecen una explicación completa de las tasas diferenciales de enfermedad y factores de riesgo entre grupos raciales, y el empleo de personas de raza autoidentificada tiene el potencial de reforzar las desigualdades raciales.[79]
Objeciones al naturalismo racial
El naturalismo racial es la opinión de que las clasificaciones raciales se basan en patrones objetivos de similitudes y diferencias genéticas. Los defensores de este punto de vista lo han justificado utilizando la evidencia científica descrita anteriormente. Sin embargo, este punto de vista es controvertido y los filósofos de la raza le han presentado cuatro objeciones principales.[80]
Las objeciones semánticas, como la objeción de la discreción, argumentan que las poblaciones humanas seleccionadas en la investigación genética de poblaciones no son razas y no corresponden a lo que significa "raza" en los Estados Unidos.
La objeción de la discreción no requiere que no haya una mezcla genética en la especie humana para que haya 'grupos raciales' de EE. UU. (...) más bien (...) lo que afirma la objeción es que la pertenencia a grupos raciales de EE. UU. es diferente de la membresía en poblaciones continentales. (...) Por lo tanto, estrictamente hablando, los negros no son idénticos a los africanos, los blancos no son idénticos a los euroasiáticos, los asiáticos no son idénticos a los asiáticos orientales, etc.[81]
Por lo tanto, se podría argumentar que la investigación científica no se trata realmente de raza. Las dos siguientes objeciones son objeciones metafísicas que argumentan que incluso si las objeciones semánticas fallan, los resultados del agrupamiento genético humano no apoyan la realidad biológica de la raza. La "objeción muy importante" estipula que las razas en la definición estadounidense no son importantes para la biología, en el sentido de que las poblaciones continentales no forman subespecies biológicas. La 'objeción objetivamente real' establece que
...los grupos raciales estadounidenses no son biológicamente reales porque no son objetivamente reales en el sentido de existir independientemente del interés humano, las creencias o algún otro estado mental de los humanos.[82]
Los naturalistas raciales han respondido a cada una de estas objeciones con argumentos en contra. También hay críticos metodológicos que rechazan el naturalismo racial debido a preocupaciones relacionadas con el diseño experimental, la ejecución o la interpretación de la investigación genética de poblaciones relevante.[83] Otra objeción semántica es la objeción de visibilidad que refuta la afirmación de que existen grupos raciales estadounidenses en las estructuras de la población humana. Filósofos como Joshua Glasgow y Naomi Zack creen que los grupos raciales estadounidenses no pueden definirse por rasgos visibles, como el color de la piel y los atributos físicos:
El material de rastreo genético ancestral no tiene efecto sobre los fenotipos o rasgos biológicos de los organismos, que incluirían rasgos considerados raciales, debido a que el material genético de rastreo ancestral no juega ningún papel en la producción de proteínas, no es el tipo de material que 'codifica' para la producción de proteínas.[84]
Una objeción diferente establece que los grupos raciales estadounidenses no son biológicamente reales porque no son objetivamente reales en el sentido de existir independientemente de algún estado mental de los seres humanos.[84][85] Se sostiene que una entidad puede ser tanto biológicamente real como socialmente construida y para capturar con precisión las entidades biológicas reales, también se deben considerar los factores sociales. Se ha argumentado que el conocimiento de la raza de una persona tiene un valor limitado, ya que las personas de la misma raza varían entre sí.[61] David J. Witherspoon y sus colegas han argumentado que cuando se asignan individuos a grupos de población, dos individuos elegidos al azar de diferentes poblaciones pueden parecerse más entre sí que un miembro elegido al azar de su propio grupo. Descubrieron que se tenían que usar muchos miles de marcadores genéticos para la respuesta a "¿Con qué frecuencia un par de individuos de una población es genéticamente más diferente que dos individuos elegidos de dos poblaciones diferentes?" ser "nunca". Esto asumió tres grupos de población, separados por grandes distancias geográficas (Europa, África y Asia Oriental). La población humana mundial es más compleja y estudiar una gran cantidad de grupos requeriría un mayor número de marcadores para la misma respuesta. Llegan a la conclusión de que "se debe tener precaución al utilizar la ascendencia geográfica o genética para hacer inferencias sobre fenotipos individuales".[86]
El hecho de que, con suficientes datos genéticos, los individuos puedan asignarse correctamente a sus poblaciones de origen es compatible con la observación de que la mayor parte de la variación genética humana se encuentra dentro de las poblaciones, no entre ellas. También es compatible con nuestro hallazgo de que, incluso cuando se consideran las poblaciones más distintas y se utilizan cientos de loci, los individuos suelen ser más similares a los miembros de otras poblaciones que a los miembros de su propia población.[87]
Esto es similar a la conclusión a la que llegó el antropólogo Norman Sauer en un artículo de 1992 sobre la capacidad de los antropólogos forenses para asignar "raza" a un esqueleto, basándose en las características craneofaciales y la morfología de las extremidades.
...la asignación exitosa de raza a un espécimen esquelético no es una reivindicación del concepto de raza, sino más bien una predicción de que un individuo, mientras estaba vivo, fue asignado a una categoría 'racial' particular construida socialmente. Un espécimen puede mostrar características que apuntan a la ascendencia africana. En este país, es probable que esa persona haya sido etiquetada como negra independientemente de si tal raza existe o no en la naturaleza.[88]
Críticas a los medicamentos basados en la raza
Troy Duster señala que la genética a menudo no es el determinante predominante de la susceptibilidad a las enfermedades, aunque podrían correlacionarse con categorías específicas definidas socialmente. Esto se debe a que esta investigación a menudo carece de control sobre una multiplicidad de factores socioeconómicos. Cita datos recopilados que indican cómo las diferencias dietéticas desempeñan un papel importante en la explicación de las variaciones de la prevalencia de la diabetes entre las poblaciones.
Duster elabora presentando el ejemplo de los pima de Arizona, una población que padece tasas desproporcionadamente altas de diabetes mellitus. La razón de esto, argumenta, no fue necesariamente el resultado de la prevalencia del gen FABP2, que está asociado con la resistencia a la insulina. Más bien, sostiene que los científicos a menudo descartan las implicaciones del estilo de vida en contextos sociohistóricos específicos. Por ejemplo, a finales del siglo XIX, la economía pima se basaba predominantemente en la agricultura. Sin embargo, a medida que la población estadounidense de origen europeo se instala en territorio tradicionalmente pima, los estilos de vida de los pima se occidentalizaron en gran medida. En tres décadas, la incidencia de diabetes aumentó en múltiples pliegues. La provisión gubernamental de alimentos gratuitos para aliviar la prevalencia de la pobreza en la población, que tenía un contenido relativamente alto de grasas, se señala como una explicación de este fenómeno.[89]
Se ha argumentado en contra de la suposición de que "la raza autoidentificada es un buen indicador de una ascendencia genética específica"[79] sobre la base de que la raza autoidentificada es compleja: depende de una variedad de factores psicológicos, culturales y sociales, y, por lo tanto, "no es un proxy sólido de la ascendencia genética".[90] Además, explican que la raza autoidentificada de un individuo se compone de otros factores colectivamente arbitrarios: opiniones personales sobre qué es la raza y hasta qué punto debe tenerse en cuenta en la vida cotidiana. Además, los individuos que comparten una ascendencia genética pueden diferir en su autoidentificación racial en contextos históricos o socioeconómicos. A partir de esto, se concluye que la precisión en la predicción de la ascendencia genética sobre la base de la autoidentificación es baja, específicamente en poblaciones mezcladas racialmente nacidas de historias ancestrales complejas.
Véase también
Referencias
- Goodman, Alan H. (2020). Race : are we so different?. Yolanda T. Moses, Joseph L. Jones (Second edición). Hoboken, NJ. ISBN 978-1-119-47247-6. OCLC 1121420797.
- Jablonski, Nina G. (2006). Skin : a natural history. Berkeley: University of California Press. ISBN 0-520-24281-5. OCLC 64592114.
- Rosenberg, Noah A.; Pritchard, Jonathan K.; Weber, James L.; Cann, Howard M.; Kidd, Kenneth K.; Zhivotovsky, Lev A.; Feldman, Marcus W. (20 de diciembre de 2002). «Genetic structure of human populations». Science (New York, N.Y.) 298 (5602): 2381-2385. ISSN 1095-9203. PMID 12493913. doi:10.1126/science.1078311.
- Lorusso, Ludovica; Bacchini, Fabio (2015-08). «A reconsideration of the role of self-identified races in epidemiology and biomedical research». Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences (en inglés) 52: 56-64. doi:10.1016/j.shpsc.2015.02.004.
- Saini, Angela (2019). Superior : the return of race science. Boston. ISBN 978-0-8070-7691-0. OCLC 1091260230.
- Marks, Jonathan (2017). Is science racist?. Malden, MA. ISBN 978-0-7456-8921-0. OCLC 961801723.
- Bamshad, Michael; Wooding, Stephen; Salisbury, Benjamin A.; Stephens, J. Claiborne (2004-08). «Deconstructing the relationship between genetics and race». Nature Reviews Genetics (en inglés) 5 (8): 598-609. ISSN 1471-0064. doi:10.1038/nrg1401. Consultado el 25 de mayo de 2021.
- Livingstone, Frank (Summer 1962). «On the Non-Existence of Human Races». Chicago Journals. Archivado desde el original el 25 de mayo de 2021. Consultado el 25 de mayo de 2021.
- Honnay, O. (2013), «Genetic Drift», Brenner's Encyclopedia of Genetics (en inglés) (Elsevier): 251-253, ISBN 978-0-08-096156-9, doi:10.1016/b978-0-12-374984-0.00616-1.
- Martin, Cyrus; Zhang, Yi (June 2007). «Mechanisms of epigenetic inheritance». Current Opinion in Cell Biology (en inglés) 19 (3): 266-272. doi:10.1016/j.ceb.2007.04.002.
- Jorde, Lynn B; Wooding, Stephen P (November 2004). «Genetic variation, classification and 'race'». Nature Genetics (en inglés) 36 (S11): S28-S33. ISSN 1061-4036. doi:10.1038/ng1435.
- «Implications of biogeography of human populations for 'race' and medicine». Nature Genetics 36 (11 Suppl): S21-7. November 2004. PMID 15507999. doi:10.1038/ng1438.
- «A global reference for human genetic variation». Nature 526 (7571): 68-74. October 2015. Bibcode:2015Natur.526...68T. PMC 4750478. PMID 26432245. doi:10.1038/nature15393.
- Cavalli-Sforza, Luigi Luca; Menozzi, Paolo; Piazza, Alberto (1994). The History and Geography of Human Genes. Princeton: Princeton University Press. ISBN 978-0-691-08750-4. Resumen divulgativo (1 December 2013).
- McDonald, J; Lehman, DC (Spring 2012). «Forensic DNA analysis». Clinical Laboratory Science 25 (2): 109-113. PMID 22693781. doi:10.29074/ascls.25.2.109.
- Butler, John. «Genetics and Genomics of Core STR Loci Used in Human Identity Testing*».
- Redd, A. J.; Chamberlain, VF; Kearney, V. F.; Stover, D; Karafet, T.; Calderon, K.; Walsh, B.; Hammer, M. F. (May 2006). «Genetic structure among 38 populations from the United States based on 11 U.S. core Y chromosome STRs.». Journal of Forensic Sciences 51 (3): 580-585. PMID 16696705. doi:10.1111/j.1556-4029.2006.00113.x.
- Hammer, M. F.; Chamberlain, V. F.; Kearney, V. F.; Stover, D.; Zhang, G; Karafet, T.; Walsh, B.; Redd, A. J. (1 de diciembre de 2006). «Population structure of Y chromosome SNP haplogroups in the United States and forensic implications for constructing Y chromosome STR databases.». Forensic Science International 164 (1): 45-55. PMID 16337103. doi:10.1016/j.forsciint.2005.11.013.
- Sims, L. M.; Ballantyne, J. (March 2008). «The golden gene (SLC24A5) differentiates US sub-populations within the ethnically admixed Y-SNP haplogroups.». Legal Medicine (Tokyo, Japan) 10 (2): 72-7. PMID 17720606. doi:10.1016/j.legalmed.2007.06.004.
- Andrea Orsucci, ""Ariani, indogermani, stirpi mediterranee: aspetti del dibattito sulle razze europee (1870–1914)" Archivado el 10 de marzo de 2002 en Wayback Machine., Cromohs, 1998 (en italiano)
- «Do Races Differ? Not Really, DNA Shows». 22 de agosto de 2000.
- Owens, Kelly; King, Mary-Claire (15 de octubre de 1999). «Genomic Views of Human History». Science 286 (5439): 451-453. ISSN 0036-8075. PMID 10521333. doi:10.1126/science.286.5439.451. «Variation in other traits popularly used to identify 'races' is likely to be due to similarly straightforward mechanisms, involving limited numbers of genes with very specific physiological effects. »
- «Multiple evidence strands suggest that there may be as few as 19,000 human protein-coding genes». Human Molecular Genetics 23 (22): 5866-5878. November 2014. PMC 4204768. PMID 24939910. doi:10.1093/hmg/ddu309.
- Sturm, Richard A.; Duffy, David L. (2012). «Human pigmentation genes under environmental selection». Genome Biology 13 (9): 248. ISSN 1474-760X. PMC 3491390. PMID 23110848. doi:10.1186/gb-2012-13-9-248.
- White, Désirée; Rabago-Smith, Montserrat (January 2011). «Genotype-phenotype associations and human eye color». Journal of Human Genetics 56 (1): 5-7. PMID 20944644. doi:10.1038/jhg.2010.126.
- Sykes, Bryan (2001). «From Blood Groups to Genes». The seven daughters of Eve. New York: Norton. pp. 32–51. ISBN 978-0-393-02018-2.
- Blancher, Antoine; Klein, Jan; Socha, Wladyslaw W. (2012). Molecular biology and evolution of blood group and MHC antigens in primates. Springer Science & Business Media. ISBN 978-3-642-59086-3.
- Ségurel, Laure; Thompson, Emma E.; Flutre, Timothée; Lovstad, Jessica; Venkat, Aarti; Margulis, Susan W.; Moyse, Jill; Ross, Steve et al. (6 de noviembre de 2012). «The ABO blood group is a trans-species polymorphism in primates». Proceedings of the National Academy of Sciences 109 (45): 18493-18498. Bibcode:2012PNAS..10918493S. ISSN 0027-8424. PMC 3494955. PMID 23091028. arXiv:1208.4613. doi:10.1073/pnas.1210603109.
- Lewontin, Richard (1972). «The Apportionment of Human Diversity». En Theodosius Dobzhansky; Max K. Hecht; William C. Steere, eds. Evolutionary Biology 6. pp. 381-398. ISBN 978-1-4684-9065-7. doi:10.1007/978-1-4684-9063-3_14.
- Risch, Neil; Burchard, Esteban; Ziv, Elad; Tang, Hua (2002). «Categorization of humans in biomedical research: genes, race and disease». Genome Biology 3 (7): comment2007.1. ISSN 1465-6906. PMC 139378. PMID 12184798. doi:10.1186/gb-2002-3-7-comment2007.
- Templeton, Alan R. (2003). «Human Races in the Context of Recent Human Evolution: A Molecular Genetic Perspective». En Goodman, Alan H.; Heath, Deborah; Lindee, eds. Genetic nature/culture: anthropology and science beyond the two-culture divide. Berkeley: University of California Press. pp. 234-257. ISBN 978-0-520-23792-6. Consultado el 23 de septiembre de 2014.
- «Race and genetics: controversies in biomedical, behavioral, and forensic sciences». The American Psychologist 60 (1): 115-128. January 2005. PMID 15641926. doi:10.1037/0003-066X.60.1.115.
- Lewontin, R. C. (2005). "Confusions About Human Races". Race and Genomics, Social Sciences Research Council. Retrieved 28 December 2006.
- Long, Jeffrey C.; Kittles, Rick A. (2009). «Human Genetic Diversity and the Nonexistence of Biological Races». Human Biology 81 (5): 777-798. ISSN 1534-6617. PMID 20504196. doi:10.3378/027.081.0621. Consultado el 13 de enero de 2016.
- Edwards AW (August 2003). «Human genetic diversity: Lewontin's fallacy». BioEssays 25 (8): 798-801. PMID 12879450. doi:10.1002/bies.10315.
- Winther, Rasmus Grønfeldt (2018). Phylogenetic Inference, Selection Theory, and History of Science: Selected Papers of A. W. F. Edwards with Commentaries. Cambridge, U.K.: Cambridge University Press. ISBN 9781107111721.
- Edwards, AWF (2003). Human genetic diversity: Lewontin's fallacy, BioEssays. pp. 798-801.
- Sesardic, N. (2010). Race: a social destruction of a biological concept. Biology and Philosophy. pp. 143-162.
- Winther, R.G. (2018). «The Genetic Reification of "Race"? A Story of Two Mathematical Methods». En R.G. Winther, ed. Phylogenetic Inference, Selection Theory, and History of Science: Selected Papers of AWF Edwards with Commentaries. pp. 489, 488-508. ISBN 9781107111721.
- Witherspoon, D. J.; Wooding, S.; Rogers, A. R.; Marchani, E. E.; Watkins, W. S.; Batzer, M. A.; Jorde, L. B. (2007). «Genetic Similarities Within and Between Human Populations». Genetics 176 (1): 351-359. ISSN 0016-6731. PMC 1893020. PMID 17339205. doi:10.1534/genetics.106.067355.
- «Genetic Structure, Self-Identified Race/Ethnicity, and Confounding in Case-Control Association Studies». American Journal of Human Genetics 76 (2): 268-75. February 2005. PMC 1196372. PMID 15625622. doi:10.1086/427888.
- Rosenberg NA; Pritchard JK; Weber JL (December 2002). «Genetic structure of human populations». Science 298 (5602): 2381-5. Bibcode:2002Sci...298.2381R. PMID 12493913. doi:10.1126/science.1078311.
- Lewontin, R. C.. «Confusions About Human Races».
- «Race, ancestry, and genes: implications for defining disease risk». Annual Review of Genomics and Human Genetics 4: 33-67. 2003. PMID 14527296. doi:10.1146/annurev.genom.4.070802.110356.
- Rosenberg NA; Mahajan S; Ramachandran S; Zhao C; Pritchard JK; Feldman MW (December 2005). «Clines, Clusters, and the Effect of Study Design on the Inference of Human Population Structure». PLOS Genetics 1 (6): e70. PMC 1310579. PMID 16355252. doi:10.1371/journal.pgen.0010070.
- Li, J. Z.; Absher, D. M.; Tang, H.; Southwick, A. M.; Casto, A. M.; Ramachandran, S.; Cann, H. M.; Barsh, G. S. et al. (2008). «Worldwide Human Relationships Inferred from Genome-Wide Patterns of Variation». Science 319 (5866): 1100-1104. Bibcode:2008Sci...319.1100L. PMID 18292342. doi:10.1126/science.1153717.
- Jakobsson, M.; Scholz, S. W.; Scheet, P.; Gibbs, J. R.; Vanliere, J. M.; Fung, H. C.; Szpiech, Z. A.; Degnan, J. H. et al. (2008). «Genotype, haplotype and copy-number variation in worldwide human populations». Nature 451 (7181): 998-1003. Bibcode:2008Natur.451..998J. PMID 18288195. doi:10.1038/nature06742.
- Xing, J.; Watkins, W. S.; Witherspoon, D. J.; Zhang, Y.; Guthery, S. L.; Thara, R.; Mowry, B. J.; Bulayeva, K. et al. (2009). «Fine-scaled human genetic structure revealed by SNP microarrays». Genome Research 19 (5): 815-825. PMC 2675970. PMID 19411602. doi:10.1101/gr.085589.108.
- López Herráez, D.; Bauchet, M.; Tang, K.; Theunert, C.; Pugach, I.; Li, J.; Nandineni, M. R.; Gross, A. et al. (2009). «Genetic Variation and Recent Positive Selection in Worldwide Human Populations: Evidence from Nearly 1 Million SNPs». En Hawks, John, ed. PLOS ONE 4 (11): e7888. Bibcode:2009PLoSO...4.7888L. PMC 2775638. PMID 19924308. doi:10.1371/journal.pone.0007888.
- Ramachandran, S; Deshpande, O; Roseman, CC; Rosenberg, NA; Feldman, MW; Cavalli-Sforza, LL (November 2005). «Support from the relationship of genetic and geographic distance in human populations for a serial founder effect originating in Africa». Proceedings of the National Academy of Sciences 102 (44): 15942-7. Bibcode:2005PNAS..10215942R. PMC 1276087. PMID 16243969. doi:10.1073/pnas.0507611102.
- Harpending, Henry (1 de noviembre de 2002). «Kinship and Population Subdivision». Population & Environment 24 (2): 141-147. doi:10.1023/A:1020815420693.
- Long, J. C.; Kittles, R. A. (2009). «Human genetic diversity and the nonexistence of biological races». Human Biology 81 (5/6): 777-798. PMID 20504196. doi:10.3378/027.081.0621.
- Mountain, J. L.; Risch, N. (2004). «Assessing genetic contributions to phenotypic differences among 'racial' and 'ethnic' groups». Nature Genetics 36 (11 Suppl): S48-S53. PMID 15508003. doi:10.1038/ng1456.
- Pearse, D. E.; Crandall, K. A. (2004). «Beyond FST: analysis of population genetic data for conservation». Conservation Genetics 5 (5): 585-602. doi:10.1007/s10592-003-1863-4.
- Hunley, Keith L.; Cabana, Graciela S.; Long, Jeffrey C. (1 de diciembre de 2015). «The apportionment of human diversity revisited». American Journal of Physical Anthropology 160 (4): 561-569. ISSN 1096-8644. PMID 26619959. doi:10.1002/ajpa.22899.
- Brace, C. Loring (2005). "Race" is a Four-letter Word: The Genesis of the Concept. Oxford: Oxford University Press. ISBN 978-0-19-517351-2. Resumen divulgativo (27 November 2010).
- Graves, Joseph L (2001). The Emperor's New Clothes: Biological Theories of Race at the Millennium. Rutgers University Press.
- Weiss, Kenneth M.; Fullerton, Stephanie M. (2005). «Racing around, getting nowhere». Evolutionary Anthropology: Issues, News, and Reviews 14 (5): 165-169. ISSN 1060-1538. doi:10.1002/evan.20079.
- Kaplan, Jonathan Michael (17 de enero de 2011). «'Race': What Biology Can Tell Us about a Social Construct». Encyclopedia of Life Sciences (ELS). ISBN 978-0470016176. doi:10.1002/9780470015902.a0005857. Consultado el 23 de septiembre de 2014.
- Luigi Luca Cavalli-Sforza, "Genes, peoples, and languages", Proceedings of the National Academy of Sciences, 1997, vol.94, pp.7719–7724, doi 10.1073/pnas.94.15.7719
- Jordge, Lynn B.; Wooding, Stephen P. (2004). «Genetic Variation, classification and 'race'». Nature Genetics 36 (11 Suppl): S28-33. PMID 15508000. doi:10.1038/ng1435.
- Paschou, Peristera; Lewis, Jamey; Javed, Asif; Drineas, Petros (2010). «Ancestry informative markers for fine-scale individual assignment to worldwide populations». J Med Genet 47 (12): 835-847. PMID 20921023. doi:10.1136/jmg.2010.078212.
- Pena, Sérgio D. J.; Di Pietro, Giuliano; Fuchshuber-Moraes, Mateus; Genro, Julia Pasqualini; Hutz, Mara H.; Kehdy, Fernanda de Souza Gomes; Kohlrausch, Fabiana; Magno, Luiz Alexandre Viana et al. (2011). «The Genomic Ancestry of Individuals from Different Geographical Regions of Brazil is More Uniform Than Expected». En Harpending, Henry, ed. PLoS ONE 6 (2): e17063. Bibcode:2011PLoSO...617063P. PMC 3040205. PMID 21359226. doi:10.1371/journal.pone.0017063.
- Parra, F. C. (2002). «Color and genomic ancestry in Brazilians». Proceedings of the National Academy of Sciences 100 (1): 177-182. Bibcode:2002PNAS..100..177P. PMC 140919. PMID 12509516. doi:10.1073/pnas.0126614100.
- Lima-Costa, M. Fernanda; Rodrigues, Laura C.; Barreto, Maurício L.; Gouveia, Mateus; Horta, Bernardo L.; Mambrini, Juliana; Kehdy, Fernanda S. G.; Pereira, Alexandre et al. (2015). «Genomic ancestry and ethnoracial self-classification based on 5,871 community-dwelling Brazilians (The Epigen Initiative)». Scientific Reports 5: 9812. Bibcode:2015NatSR...5E9812.. PMC 5386196. PMID 25913126. doi:10.1038/srep09812.
- Bolnick, Deborah A. (2008). «Individual Ancestry Inference and the Reification of Race as a Biological Phenomenon». En Koenig, Barbara A.; Richardson, Sarah S.; Lee, eds. Revisiting race in a genomic age. Rutgers University Press. ISBN 978-0-8135-4324-6.
- Patterson, Nick; Price, Alkes L.; Reich, David (2006). «Population Structure and Eigenanalysis». PLOS Genetics 2 (12): e190. PMC 1713260. PMID 17194218. doi:10.1371/journal.pgen.0020190.
- Rosenberg, N. A.; Mahajan, S.; Ramachandran, S.; Zhao, C.; Pritchard, J. K. (2005). «Clines, Clusters, and the Effect of Study Design on the Inference of Human Population Structure». PLOS Genet 1 (6): e70. PMC 1310579. PMID 16355252. doi:10.1371/journal.pgen.0010070.
- Duster, Troy (March 2015). «A post-genomic surprise. The molecular reinscription of race in science, law and medicine». The British Journal of Sociology 66 (1): 1-27. ISSN 0007-1315. PMID 25789799. doi:10.1111/1468-4446.12118.
- Fullwiley, D. (2008). "The Biologistical Construction of Race: 'Admixture' Technology and the New Genetic Medicine". Social Studies of Science, 38(5), 695–735. doi 10.1177/0306312708090796
- Risch N (July 2005). «The whole side of it--an interview with Neil Risch by Jane Gitschier». PLOS Genetics 1 (1): e14. PMC 1183530. PMID 17411332. doi:10.1371/journal.pgen.0010014.
- «Active intestinal chloride secretion in human carriers of cystic fibrosis mutations: an evaluation of the hypothesis that heterozygotes have subnormal active intestinal chloride secretion». Am. J. Hum. Genet. 67 (6): 1422-1427. December 2000. PMC 1287919. PMID 11055897. doi:10.1086/316911.
- Yudell, Michael; Roberts, Dorothy; DeSalle, Rob; Tishkoff, Sarah (2016). «Taking Race out of Human Genetics». Science 351 (6273): 564-65. PMID 26912690. doi:10.1126/science.aac4951.
- Schwartz, Robert S. (2001). «Racial Differences in the Response to Drugs — Pointers to Genetic Differences». New England Journal of Medicine 344 (18): 1393-1396. PMID 11333999. doi:10.1056/NEJM200105033441810.
- Bloche, Gregg M. (2004). «Race-Based Therapeutics». New England Journal of Medicine 351 (20): 2035-2037. PMID 15533852. doi:10.1056/nejmp048271.
- Drug information for the drug Crestor. Warnings for this drug state, "People of Asian descent may absorb rosuvastatin at a higher rate than other people. Make sure your doctor knows if you are Asian. You may need a lower than normal starting dose."
- Risch, N.; Burchard, E.; Ziv, E.; Tang, H. (2002). «Categorization of humans in biomedical research: genes, race and disease». Genome Biol 3 (7): 1-12. PMC 139378. PMID 12184798. doi:10.1186/gb-2002-3-7-comment2007.
- «Census, Race and Science». Nature Genetics 24 (2): 97-98. 2000. PMID 10655044. doi:10.1038/72884.
- Lorusso, Ludovica; Bacchini, Fabio (2015). «A reconsideration of the role of self-identified races in epidemiology and biomedical research». Studies in History and Philosophy of Biological and Biomedical Sciences 52: 56-64. PMID 25791919. doi:10.1016/j.shpsc.2015.02.004.
- «Revelations», Shut Up and Listen (Palgrave Macmillan), ISBN 978-0-230-36298-7, consultado el 28 de enero de 2021.
- Spencer, Quayshawn (2015). «Philosophy of race meets population genetics». Studies in History and Philosophy of Biological and Biomedical Sciences 52: 49. PMID 25963045. doi:10.1016/j.shpsc.2015.04.003.
- Spencer, Quayshawn (2015). «Philosophy of race meets population genetics». Studies in History and Philosophy of Biological and Biomedical Sciences 52: 51. PMID 25963045. doi:10.1016/j.shpsc.2015.04.003.
- Spencer, Quayshawn (2015). «Philosophy of race meets population genetics». Studies in History and Philosophy of Biological and Biomedical Sciences 52: 46-47. PMID 25963045. doi:10.1016/j.shpsc.2015.04.003.
- Zach, Naomi (2002). «Philosophy of Science and Race». The International Journal of African Historical Studies 36 (3): 656-658. doi:10.2307/3559445.
- Sundstrom, R (2002). «Race as a human kind». Philosophy & Social Criticism 28: 91-115. doi:10.1177/0191453702028001592.
- «Genetic Similarities Within and Between Human Populations». Genetics 176 (1): 351-9. May 2007. PMC 1893020. PMID 17339205. doi:10.1534/genetics.106.067355.
- «Genetic Similarities Within and Between Human Populations». Genetics 176 (1): 358. May 2007. PMC 1893020. PMID 17339205. doi:10.1534/genetics.106.067355.
- Sauer, N. J. (January 1992). «Forensic anthropology and the concept of race: if races don't exist, why are forensic anthropologists so good at identifying them?». Social Science & Medicine 34 (2): 107-111. PMID 1738862. doi:10.1016/0277-9536(92)90086-6.
- Duster, Troy (2015). «A post-genomic surprise. The molecular reinscription of race in science, law and medicine». The British Journal of Sociology 66 (1): 1-27. PMID 25789799. doi:10.1111/1468-4446.12118.
- Hunt, L. M.; Megyesi, M.S. (Fall 2007). «The ambiguous meanings of the racial/ethnic categories routinely used in human genetics research». Social Science and Medicine 66 (2): 349-361. PMC 2213883. PMID 17959289. doi:10.1016/j.socscimed.2007.08.034.
Otras lecturas
- Helms JE, Jernigan M, Mascher J (January 2005). «The meaning of race in psychology and how to change it: a methodological perspective». The American Psychologist 60 (1): 27-36. PMID 15641919. doi:10.1037/0003-066X.60.1.27. Archivado desde el original el 26 de febrero de 2019. Consultado el 25 de mayo de 2021.
- Keita SO, Kittles RA, Royal CD, etal (November 2004). «Conceptualizing human variation». Nature Genetics 36 (11 Suppl): S17-20. PMID 15507998. doi:10.1038/ng1455.
- Koenig, Barbara A.; Lee, Sandra Soo-jin; Richardson, Sarah S., eds. (2008). Revisiting Race in a Genomic Age. New Brunswick (NJ): Rutgers University Press. ISBN 978-0-8135-4324-6. This review of current research includes chapters by Jonathan Marks, John Dupré, Sally Haslanger, Deborah A. Bolnick, Marcus W. Feldman, Richard C. Lewontin, Sarah K. Tate, David B. Goldstein, Jonathan Kahn, Duana Fullwiley, Molly J. Dingel, Barbara A. Koenig, Mark D. Shriver, Rick A. Kittles, Henry T. Greely, Kimberly Tallbear, Alondra Nelson, Pamela Sankar, Sally Lehrman, Jenny Reardon, Jacqueline Stevens, and Sandra Soo-Jin Lee.
- Lieberman, Leonard; Kirk, Rodney C.; Corcoran, Michael (2003). «The Decline of Race in American Physical Anthropology». Przegląd Antropologiczny – Anthropological Review 66: 3-21. ISSN 0033-2003. Archivado desde el original el 8 de junio de 2011. Consultado el 12 de septiembre de 2010.
- Long JC, Kittles RA (August 2003). «Human genetic diversity and the nonexistence of biological races». Human Biology 75 (4): 449-71. PMID 14655871. S2CID 26108602. doi:10.1353/hub.2003.0058.
- Miththapala, Sriyanie; Seidensticker, John; O'Brien, Stephen J. (1996). «Phylogeographic Subspecies Recognition in Leopards (Panthera pardus): Molecular Genetic Variation». Conservation Biology 10 (4): 1115-1132. doi:10.1046/j.1523-1739.1996.10041115.x.
- Ossorio P, Duster T (January 2005). «Race and genetics: controversies in biomedical, behavioral, and forensic sciences». The American Psychologist 60 (1): 115-28. PMID 15641926. doi:10.1037/0003-066X.60.1.115.
- Parra EJ, Kittles RA, Shriver MD (November 2004). «Implications of correlations between skin color and genetic ancestry for biomedical research». Nature Genetics 36 (11 Suppl): S54-60. PMID 15508005. doi:10.1038/ng1440.
- Sawyer SL, Mukherjee N, Pakstis AJ, etal (May 2005). «Linkage disequilibrium patterns vary substantially among populations». European Journal of Human Genetics 13 (5): 677-86. PMID 15657612. doi:10.1038/sj.ejhg.5201368.
- Rohde DL, Olson S, Chang JT (September 2004). «Modelling the recent common ancestry of all living humans». Nature 431 (7008): 562-6. Bibcode:2004Natur.431..562R. PMID 15457259. S2CID 3563900. doi:10.1038/nature02842.
- Serre D, Pääbo S (September 2004). «Evidence for Gradients of Human Genetic Diversity Within and Among Continents». Genome Research 14 (9): 1679-85. PMC 515312. PMID 15342553. doi:10.1101/gr.2529604.
- Smedley A, Smedley BD (January 2005). «Race as biology is fiction, racism as a social problem is real: Anthropological and historical perspectives on the social construction of race». The American Psychologist 60 (1): 16-26. PMID 15641918. doi:10.1037/0003-066X.60.1.16.
| Control de autoridades |
|
|---|
 Datos: Q7278940
Datos: Q7278940 Multimedia: Race and genetics / Q7278940
Multimedia: Race and genetics / Q7278940