Sistema de tres dominios
El sistema de los tres dominios es una clasificación biológica propuesta por Carl Woese y colaboradores en 1977 que clasifica el árbol de la vida en tres grupos: Bacteria (en sentido restringido, excluyendo a las arqueas, a diferencia de la hasta en ese momento concepción del grupo), Arquea y Eukaryota. Woese se basó en las diferencias encontradas en la secuencia del ARN ribosomal de la subunidad menor, para concluir que estos grupos se desarrollaron por separado de un progenitor común llamado progenote.
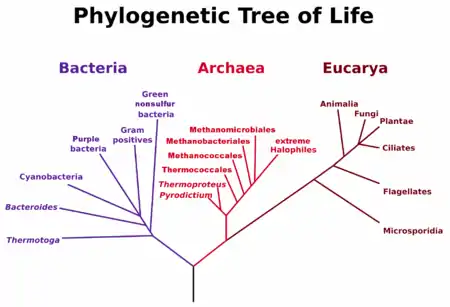
Este sistema llamado de tres dominios, reemplazó al anterior sistema de dos imperios formado por los grupos Eucariota y Procariota; sin embargo, a la luz de los postulados más recientes, es posible que se abandone este modelo ya que no corresponde con el árbol filogenético de la vida.[2]
Para describir estas tres grandes ramas, Woese se permitió tratarlas como dominios en 1990,[1] siempre promoviendo la separación de los dos grupos procariotas.
Aunque rápidamente la mayoría de los sistematistas moleculares aceptó el sistema de tres dominios, algunos biólogos como Mayr lo criticaron por dar demasiado énfasis a la singularidad de las arqueas y por ignorar las fuertes similitudes entre los grupos.
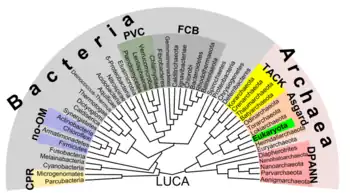
Argumentos a favor
Se ha confirmado que la membrana de las células de las arqueas tiene una composición inusual, así como la estructura de sus flagelos.[3] Otras diferencias significativas son los sistemas de las arqueas para la replicación del ADN y la transcripción, que son bastante diferentes a las de los eucariotas. Por ejemplo, las ARN polimerasa de las arqueas constan de hasta 14 subunidades, mientras que las ARN polimerasa de las bacterias solo tiene 4. Los análisis de dichas subunidades sugieren que están más estrechamente relacionadas con las encontradas en los eucariotas. Así también, las arqueas producen un número de proteínas de unión al ADN similar a las histonas de los eucariotas.[4]
Semejanzas y diferencias
La siguiente tabla comparativa relaciona las diferentes características eucariotas, bacterianas y arqueanas:
| Dominios | BACTERIA | ARCHAEA | EUCARYA |
|---|---|---|---|
| Estructura celular | procariota | procariota | eucariota |
| Cromosomas | uno circular y desnudo |
uno circular con cromatina | múltiples lineales con cromatina |
| Composición de la pared celular | peptidoglicano | glicopéptidos, proteínas o glúcidos |
glúcidos (celulosa, quitina) |
| Enlace lipídico de la membrana | éster | éter | éster |
| Ácidos grasos de la membrana | lineales | ramificados | lineales |
| Flagelo | bacteriano | arqueano | eucariota |
| Ribosoma | 70S | 70S | 80S, pero 70S (o 55S) en mitocondrias y cloroplastos |
| Intrones | No | en los genes de ARNt | en la mayoría de genes |
| Operones | Sí | Sí | No |
| Plásmidos | Sí | Sí | raros |
| Cromatina con nucleosomas e histonas | No | Sí | Sí |
| Iniciador de ARNt | formil-metionina | metionina | metionina |
| Factores de elongación (EF) | EF-Tu, -Ts, -G y -P | aEF1A, 1B, 2 y 5A | eEF1A, 1B, 2, 5A y mitocondriales: mtEFTu, Ts y G1 |
| ARN polimerasas | una | una | varias |
| Subunidades del ARNP | 4-5 | 8-14 | 12-14 |
| Estreptomicina y cloranfenicol | sensible | resistente | resistente |
| Toxina diftérica | resistente | sensible | sensible |
Las principales características que llevaron a reconocer tres diferentes dominios fueron los siguientes análisis del ARN ribosomal de la subunidad menor: en Bacteria hay una secuencia particular entre las posiciones 500-545 del ARNr-16S, en Archaea hay una estructura única entre las posiciones 180-197 o 405-498 del mismo ARNr-16S y en Eukarya el ARNr de la subunidad menor es 18S y difiere de los procariontes entre las posiciones 585-656.[5]
Filogenia
Existen varios postulados diferentes sobre la relación filogenética entre los tres dominios, los cuales pueden resumirse cronológicamente del siguiente modo:
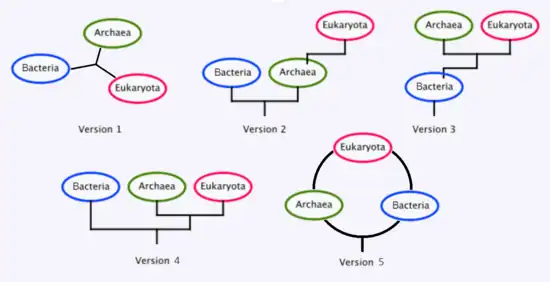
2. Lake et al. 1984, hipótesis del eocito, origen de Eukaryota a partir de Archaea.
3. Cavalier-Smith 1987, hipótesis neomurana del origen de Archaea y Eukaryota a partir de una Actinobacteria.
4. Woese et al. 1990, hipótesis de los tres dominios.
5. Rivera & Lake 2004, anillo de la vida, origen eucariota por simbiogénesis entre una arquea y una bacteria.
Historia
Situación previa
Durante los siglos XIX y XX se hicieron notables avances en el conocimiento microbiológico. Sin embargo esto no significó avances en filogenia y clasificaciones naturales en procariontes. La clasificación de plantas y animales se basaba en anatomía comparada y embriología, en cambio las bacterias carecen de complejidad morfológica y tienen enorme diversidad fisiológica.
El manual de Bergey desde los años 60 a los 80, optó por dar clasificaciones no naturales, pero razonables, en lugar de especular filogenias que cambiasen continuamente (ver clasificación de Monera). Muchos especialistas (Stanier, van Niel, Winogradsky) se resignaron a aceptar que una clasificación filogenética procariota era imposible, a pesar de la aceptación en general de que son un grupo coherente que está relacionado con el origen monofilético de la vida.
Descubrimiento
El gran paso en biología molecular se da cuando un equipo de la Universidad de Illinois, realiza análisis genéticos en los años 70 usando el método de secuencia molecular del ARN ribosomal procariota 16S. Además los ribosomas son abundantes en cada célula y fáciles de extraer, haciendo de este organelo el favorito para la investigación. El resultado fue toda una revolución en la elaboración de árboles filogenéticos; así se vio renacer la taxonomía microbiana que hasta entonces parecía imposible y aparece un nuevo campo dentro del amplio estudio procariota.
En 1977, se identificó un grupo sui generis de bacterias metanógenas que figuraban en la prueba de ARNr-16S alejadas de las demás bacterias. Se concluyó de este modo que la mayor divergencia entre organismos procariotas se da entre los metanógenos (arqueas) y las demás bacterias.[6]
Ese mismo año, Carl Woese y G. Fox, observan que los metanógenos son tan distantes de las bacterias como de los eucariontes, declarando en consecuencia el descubrimiento de “una tercera forma de vida”. Asumiendo que las condiciones en la Tierra durante el inicio de sus tiempos fueron anaerobias, se concluyó que los metanógenos serían un linaje muy antiguo y se les llamó arqueobacterias (Archaebacteria). De este modo se determinó que la más alta categoría taxonómica no estaba en la dicotomía procariota-eucariota, sino en tres reinos primarios o urkingdoms: "eubacteria, archaebacteria y urkaryotes".[7]
En 1990, Woese y colaboradores, usan el mismo análisis para la comparación con otros organismos. Se observó que los metanógenos y un grupo importante de extremófilos (conocidos como eocitos) figuraban como un grupo coherente con la prueba de ARNr-16S, postulando el sistema de tres dominios y renombrando a los tres grupos: Archaea, Bacteria y Eucarya, los cuales tienen diferencias distintivas a nivel de la subunidad menor del ARN ribosomal y desestiman a Procaryotae o Monera como taxón válido. Los tres dominios tendrían una antigüedad de 3 a 4 mil millones de años descendiendo desde un ancestro común[1] denominado progenote o protobionte. Este mismo estudio postula que las arqueas (Archaea) pueden dividirse en dos reinos: el de los euriotas (Euryarchaeota) que son metanógenos y el de los crenotas (Crenarchaeota) que son extremófilos.
Análisis proteico
El análisis de alineamiento de secuencias de proteínas, ha producido árboles filogenéticos que avalan las investigaciones previas del ARNr, respaldando fuertemente la separación natural en los tres dominios descubiertos (Brown 2001).[8]
Posición a favor del sistema
Ya que en el origen eucariota (eucariogénesis) está involucrado un evento único de fusión biológica entre una arquea y una bacteria, el dominio eucariota resulta fundamentalmente diferente de los otros dominios de varias formas cruciales, lo que justifica mantener el sistema de tres dominios como un sistema natural de organismos.[9] Esto significa que podemos hablar de dos dominios primarios (Archaea y Bacteria) y uno secundario (Eukaryota) que viene a ser menos antiguo.
Descubrimiento de un posible 4 dominio
En 2012 en fuentes hidrotermales de Japón se llevó a cabo el descubrimiento de Parakaryon myojinensis un organismo unicelular que presenta características que no encajan con las células de los otros tres dominios y probablemente constituya su propio dominio Parakaryota. Parakaryon myojinensis al igual que los eucariotas tiene núcleo y otros endosimbiontes en su célula, sin embargo su envoltura nuclear es de una sola capa, no hecha de dos membranas concéntricas como en cualquier eucariota y el material genético está almacenado como en las bacterias, en filamentos y no en cromosomas lineales. Además no presenta retículo endoplásmico, aparato de Golgi, citoesqueleto, mitocondrias, poros nucleares y carece totalmente de flagelo. Los ribosomas se encuentran no solo en el citoplasma sino también en el núcleo.[10] Posee una pared celular compuesta por peptidoglucanos al igual que las bacterias y su modo de nutrición sería osmótrofa.[11]
|
Es difícil precisar su relación filogenética con los otros seres vivos porque solo se ha encontrado un espécimen y su genoma no ha podido ser secuenciado. Según algunos autores sería un organismo intermediario entre los procariotas y los eucariotas.[12]
Críticas contra el sistema
Hipótesis de 2 dominios
Muchos autores, en especial los biólogos evolutivos, no aceptan el sistema de tres dominios; aunque comúnmente se incorpora a Archaea y Bacteria (o Eubacteria) como procariotas (por ejemplo AHG Balows y otros).[13]
Lynn Margulis rechaza el sistema de 3 dominios y apoya la división en dos grandes grupos, definiendo a los procariontes por su simplicidad y a los eucariontes por su evolución endosimbiótica. Sostiene que la mayor discontinuidad existente entre todas las formas de vida presentes en la Tierra, es la que hay entre procariotas y eucariotas, pues la especiación, en su mayor sentido, no era posible antes del origen y evolución de los eucariontes por simbiogénesis. Son dos los metabolismos eucariotas, heterótrofo (animales, hongos) que depende de las mitocondrias y autótrofo (plantas, algas) que depende de los cloroplastos. En cambio hay más de 20 metabolismos procariotas. No es comprensible la biología eucariota sin reconocer el origen bacteriano de mitocondrias y cloroplastos. El sistema en 3 dominios se basa en algunos aspectos genéticos, pero ignora los aspectos de la historia de la vida, estructura genética, ecología, relaciones simbióticas, morfología y desarrollo evolutivo.[14]
Ernst Mayr, en 1990 y 1998 defiende la unidad del imperio Prokaryota y sostiene que la carencia o no-posesión de una característica es tan importante en cualquier clasificación como su posesión, salvo en los casos en que la carencia está demostrada por una pérdida secundaria. Critica al sistema de tres dominios pues piensa que la evolución no es solo una cuestión de fenotipos, y resalta la complejidad y tamaño de los eucariontes.[15]
R. Gupta considera que la transición de procariontes a eucariontes es la mayor discontinuidad evolutiva de la historia de la Tierra; las diferencias ente ellos son enormes y la transición muy brusca.[16]
Tom Cavalier-Smith sostiene que el origen de los eucariontes fue resultado de los cambios más radicales en estructura celular y mecanismos de división, en la historia de la vida. Eukaryota es el principal ejemplo de la complejidad del proceso evolutivo y, sobre todo, de la masiva creación de nuevos genes y proteínas. El sistema de 3 dominios ha ignorado los datos fósiles, los cuales dan 900 Ma (millones de años) a los eucariontes y 3500 Ma a los procariontes.[17] Estando los eucariontes más cercanos evolutivamente a las arqueas, se deduce que estas últimas no son muy antiguas a pesar del nombre, su origen estaría relacionado con la adaptación a condiciones anaerobias termoacidófilas, y los metanógenos fueron probablemente responsables de las superglaciaciones del Proterozoico por alteración del balance atmosférico. Según Cavalier-Smith, se ha exagerado la diferencia entre los ARNr procariotas; y en las demás características son mayores las homologías.
Un estudio de 2008 que analizó 53 genes de los tres dominios y que incluye a componentes esenciales de la replicación, transcripción y sistemas de traducción de los ácidos nucleicos eucariotas, utilizó modelos filogenéticos que permiten cambiar las composiciones de nucleótidos o aminoácidos sobre los árboles elaborados; dando como resultado el apoyo a la hipótesis del eocito de James Lake que postula el origen arqueano de los eucariontes, rechazando tanto la monofilia del grupo Archaea (que resulta parafilético respecto a Eucarya), como el árbol de los tres dominios de la vida.[18] Filogenéticamente, la diversidad profunda de arqueas y bacterias eclipsa por completo a la diversidad eucariota.[19] En general se puede decir que la gran mayoría de teorías relacionadas con el origen eucariota, tales como la endosimbiosis seriada, hipótesis del hidrógeno, hipótesis fagotrófica y otras, son incompatibles con el modelo de tres dominios, el cual habría caído en error principalmente por el factor de la atracción de ramas largas.
La rivalidad entre los sistemas de tres dominios y el de dos imperios llegó a tener un debate encendido entre biólogos, en donde Woese fue atacado incluso con epítetos como "loco chiflado" y por el otro lado sus defensores calificaban a los tradicionalistas de "dogmáticos" y cerrados a los nuevos descubrimientos.[20] Sin embargo es relevante destacar que el gran mérito de Woese y sus colaboradores, fue el descubrimiento del análisis genético ribosomal, el cual permitió profundizar en la filogenia procariota, hasta entonces esquiva; un método que se usa ahora extensamente, incluso hasta por los críticos del -por ahora vigente- sistema de tres dominios.
Así menciona Cavalier-Smith (1998)[21] el tema:
"La caracterización de Archaebacteria y de sus sustanciales diferencias con respecto a Eubacteria (Woese y Fox 1977a,[22] Woese 1987[23]) han sido de profunda importancia para nuestra comprensión de la diversidad de bacterias. También lo han sido los métodos para determinar la filogenia basados en ARNr cuyo pionero fue Woese (1987,[23] ver también Olsen y Woese 1993[24]) para el progreso del conocimiento filogenético de bacterias y de eucariotas. No es mi deseo minimizar la gran importancia de estas muy positivas contribuciones a la ciencia, pero debo enfatizar, como ya lo he hecho tibiamente en una publicación anterior (Cavalier-Smith 1986c[25]), que la importancia sistemática de la distinción entre arquebacterias y eubacterias ha sido enormemente exagerada, una visión fuertemente sostenida por Mayr (1990[26]). (...) Una larga charla con George Fox (nota: uno de los proponentes de la clasificación en 3 dominios) hace 10 años hizo claro para mí que cuando Woese empezó a criticar el concepto de procariota (Woese y Fox 1977b[27]) no tenían las ideas de Stanier (1961) y Van Niel (1962) (nota: de las profundas diferencias entre procariotas y eucariotas) en mente en absoluto, sino que tenían en mente la visión naïve (ingenua) entonces popular entre los biólogos moleculares de que la bacteria Escherichia coli era un típico procariota y que estudiando su biología molecular toda la biología de las bacterias sería revelada. Esta visión naïve claramente merecía ser criticada. Lo más desafortunado fue la forma en que la crítica fue realizada, ya que fue hecha sobre qué es lo que constituye un procariota, más que sobre las visiones naïve de los biólogos moleculares sobre la materia. Los biólogos moleculares, siendo orientados en química más que en morfología, no se enteraron demasiado de los conceptos de Stanier (1961, 1970,[28] Stanier y Van Niel 1962), que son principalmente de la biología de la célula. En lugar de eso ellos se figuraban un procariota en términos de características genéticas como la estructura del operón y la organización del promotor en los cuales se conocía que E. coli difería de los eucariotas. Woese y Fox (1977b[27]) estuvieron correctos al criticar el supuesto de que todas las bacterias tenían que tener necesariamente estos caracteres, pero fallaron en darse cuenta de que estos detalles no tenían un rol en la definición de bacteria en términos de su organismo básico y la biología de su célula. Exagerar la distinción biológica de Archaebacteria fue, sin embargo, útil en sentido histórico ya que promovió que una horda de bacteriólogos muy competentes y biólogos moleculares las estudiaran esperando que ellas muestren algunas diferencias verdaderamente radicales en su organización genética con respecto a E. coli y a los eucariotas.""Lo que esta enorme empresa ha mostrado, sin embargo, es que Archaebacteria no son en realidad tan diferentes de E. coli en la organización de sus genes (por ejemplo en los operones) y genomas, ni en su maquinaria de replicación, transcripción y traducción, como han aceptado claramente Keeling, Charlebois y Doolittle (1994[29]). Las primeras secuencias de todo el genoma de eubacterias y arquebacterias reforzó la visión de que la organización genética de todas las bacterias [aquí sinónimo de procariotas] es fundamentalmente la misma, y radicalmente diferente de los eucariotas (Edgell et al. 1996). Se admitieron algunas diferencias, pero éstas, excepto por algunos aspectos de la transcripción, son relativamente pequeñas, principalmente de interés para el biólogo molecular especializado más que para el bacteriólogo general, y estas propiedades especiales de las arquebacterias son en general compartidas con los eucariotas, más que indicativas de una tercer forma de vida. Estas diferencias empalidecen en significancia en comparación con las diferencias inmensamente más profundas entre la organización genética de los eucariotas y los procariotas, de lo cual Stanier (1961, 1970[28]) tenía muy poca noticia, pero que ya he discutido en detalle en otros trabajos (Cavalier-Smith 1981b,[30] 1987c[31] ,[32] 1991a, b, 1993b[33]). Esta conclusión claramente refuta la idea de que Archaebacteria y Eubacteria surgieron independientemente de un progenota precelular crudo con mecanismos genéticos pobremente desarrollados (Woese y Fox 1977b[27]). Esa idea fue basada en las grandes diferencias entre los dos taxones reveladas por el catálogo de ARNr 16S. La forma en la cual esta técnica exagera severamente las diferencias cuantitativas fue severamente criticada por Hori, Itoh y Osawa (1982[34]), y la validez de su crítica ha sido completamente confirmada cuando se secuenció el ARN total. Sin embargo, aun en la época en que fue propuesto, probablemente era obvio para cualquier biólogo celular, pero curiosamente no lo era para ningún biólogo molecular, que la idea de que eubacterias, arquebacterias y eucariotas hayan evolucionado independientemente no hubiera podido ser correcta jamás. Hacía ya mucho tiempo que se sabía que los eucariotas y las eubacterias compartían tantas características en la organización de la célula y el metabolismo, que su ancestro común debía haber sido una célula altamente desarrollada con muchos miles de genes, y con una maquinaria de replicación de ADN altamente desarrollada (Edgell y Doolittle 1997[35]), por lo que no podía haber sido un progenota con unas crudamente desarrolladas maquinarias de replicación, reparación de ADN, transcripción y traducción como sugirieron Woese y Fox (1977b[27]). Muchos autores se han dado cuenta de esto (por ejemplo Ouzonis y Kyrpides 1996[36])."
"(...) Me sentí obligado a contar esta historia en algún detalle porque las ideas tempranas de Woese y Fox (1977a,[22] b[27]) acerca de la divergencia independiente y tempranamente primaria de arquebacterias, eubacterias y eucariotas a partir de un progenota (si bien hoy en día no es aceptada ni siquiera por sus mismos autores) y su crítica equivocada del concepto de bacteria y procariotas, han tenido un rol importante en la determinación de las visiones actuales de muchos biólogos acerca del estatus de las arquebacterias. (...)"
"Como ya conté en una publicación previa (Cavalier-Smith 1986c[25]) no hay buenas razones para igualar el rango de los tres taxones, eucariotas, archaebacterias y eubacterias, ni como dominios (Woese, Kandler y Wheelis 1990[37]) ni tampoco como reinos. (...) Igualar el rango de estos taxones produce un sistema grotescamente desbalanceado para los organismos vivientes como conjunto."Cavalier-Smith 1998[21]
Como se ha mostrado (Cavalier-Smith 2002a.[38] 2006a, c), ignorar la estructura de los organismos, la biología de la célula y la paleontología llevó a una ahora generalizada interpretación fundamental, pero equivocada, de la historia de la vida, en el sistema de tres dominios, que enraiza el árbol entre neomuras y eubacterias (Woese et al. 1990[37]). Este serio error se basa no solo en no integrar la evidencia de secuencias con otros datos, sino también en no tener en cuenta la naturaleza muchas veces extremadamente no-reloj de la evolución de las secuencias, y de errores sistemáticos que guiaron fuertemente otras conclusiones, en los árboles de secuencias para moléculas que no evolucionan de acuerdo con preconcepciones estadísticas inocentes[38] Los análisis de transición usando complejos caracteres tridimensionales están menos dispuestos a artefactos filogenéticos que las secuencias, y proveen poderosa evidencia de que la topología del árbol es muy diferente(Valas y Bourne 2009[39]). La paleontología provee evidencia igualmente convincente de que las cianobacterias son sustancialmente más antiguas que los eucariotas.
Solo dos dominios primarios
Un aspecto que postula que el árbol de tres dominios puede ser incorrecto, está en que una arquea participó del origen eucariota al fundar el linaje del huésped para el endosimbionte mitocondrial; por lo que solo hay respaldo para dos dominios primarios de la vida: Archaea y Bacteria.[40]
Críticas a la redefinición de Bacteria
En la clasificación cladista en supergrupos de Adl et al. (2005) reconocen que el término parafilético "bacterias" es el más conveniente para referirse a los procariotas, y reservan al grupo que excluye a las arqueas el nombre Eubacteria: "Reconocemos a los procariotas como incluyendo a Archaea y Eubacteria, con (el nombre) bacteria siendo un nombre vulgar conveniente para los procariotas" (We recognize prokaryotes to include the Archaea and the Eubacteria, with bacteria being a convenient common term for prokaryotes (Cavalier-Smith 2002; Walsh and Doolittle 2005).)"
Actualmente, existe amplia aceptación en la redefinición de los términos Archaea y Bacteria, aunque pocos siguen prefiriendo utilizar Eubacteria.
Referencias
- C. R. Woese, O. Kandler & M. L. Wheelis 1990. Towards a natural system of organisms: Proposal of the domains Arquea, Bacteria and Eucariota. Proc. Natl. Acad. Sci. USA
- Hug, L. A., Baker, B. J., Anantharaman, K., Brown, C. T., Probst, A. J., Castelle, C. J., ... & Suzuki, Y. (2016). A new view of the tree of life. Nature Microbiology, 1, 16048.
- Nikhil A Thomas€t al 2001, The archaeal flagellum: a different kind of prokaryotic motility structure FEMS Microbiology Reviews. Volume 25, Issue 2, pages 147–174, April 2001
- Malcolm F. White and Stephen D. Bell 2002, Holding it together: chromatin in the Archaea. TRENDS in Genetics Vol.18 No.12
- Helena Curtis, Sue Barnes, Adriana Schnek 2008, Biologia Cap.24 Bacteria y Archaea. Cuadro 24-1, p.459
- Balch WE, Magrum LJ, Fox GE, Wolfe RS, Woese CR. 1977, An ancient divergence among the bacteria. J Mol Evol. 1977 Aug 5;9(4):305-11.
- C R Woese and G E Fox 1977, Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proc Natl Acad Sci U S A. 1977 November; 74(11): 5088–5090.
- Brown JR et al 2001. Universal trees based on large combined protein sequence data sets. Nat Genet. 2001 Jul;28(3):281-5.
- P.T.S. van der Gulik, W.D. Hoff & D. Speijer 2017, In defence of the three-domains of life paradigm. BMC Evolutionary Biology (2017) 17:218 DOI 10.1186/s12862-017-1059-z
- Nick Lane (2015). «Epilogue: From the Deep». The Vital Question: Energy, Evolution, and the Origins of Complex Life. W.W.Norton and Company. pp. 281–290. ISBN 978-0-393-08881-6. Parámetro desconocido
|title-link=ignorado (ayuda) - Evolution of complex life on Earth, take 2
- Yamaguchi M, Mori Y, Kozuka Y, Okada H, Uematsu K, Tame A, Furukawa H, Maruyama T, Worman CO, Yokoyama K. (2012). «Prokaryote or eukaryote? A unique microorganism from the deep sea.». J Electron Microsc (Tokyo) 61 (6): 423-431. doi:10.1093/jmicro/dfs062.
- Balows, A. H. G. Trüper, M. Dworkin, W. Harder, and K. H. Schleifer (ed.) 1992. The prokaryotes, 2nd ed., vol. 1:, p. vii. Springer-Verlag, New York, N.Y.
- Lynn Margulis & Michael J Chapman, 1982-1998-2009, "Kingdoms and Domains: An Illustrated Guide to the Phyla of Life on Earth." p53
- Aharon Oren & R. Thane Papke 2010. Molecular Phylogeny of Microorganisms. Caister Academic Press. p 12
- El País 2001, 'Los eucariotas son la mayor discontinuidad de la historia' ENTREVISTA: Radhey Gupta. Evolucionista molecular
- Thomas Cavalier-Smith 2006, Cell evolution and Earth history: stasis and revolution. Phil. Trans. R. Soc. B 29 June 2006 vol. 361 no. 1470 969-1006
- Cox, Cymon J. et al 2008, The archaebacterial origin of eukaryotes Proc Natl Acad Sci U S A. 2008 December 23; 105(51): 20356–20361. doi: 10.1073/pnas.0810647105
- Cindy J. Castelle & Jillian F. Banfield 2018, Major New Microbial Groups Expand Diversity and Alter our Understanding of the Tree of Life. PERSPECTIVE, VOL 172, Issue 6, P1181-1197, 2018, DOI:https://doi.org/10.1016/j.cell.2018.02.016
- Jeffrey Marlow 2013, Carl Woese, (R)evolutionary Biologist The extremo files.
- Cavalier-Smith, T. (1998). A revised six-kingdom system of life., Medicine Biology - Published in Biological reviews of the Cambridge Philosophical Society, Biol. Rev. 73: 203-266. DOI:10.1111/j.1469-185X.1998.tb00030.x
- Woese, C. R. & Fox, G. E. (1977a). Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proceedings of the National Academy of Sciences, USA 74 , 5088–5090.
- Woese, C. R. (1987). Bacterial evolution. Microbiological Reviews 51, 221–271.
- Olsen, G. J. & Woese, C. R. (1993). Ribosomal RNA, a key to phylogeny. FASEB Journal 7 , 113–123.
- Cavalier-Smith, T. (l986c). The kingdoms of organisms. Nature 324 , 4l6–4l7
- Mayr, E. (1990). A natural system of organisms. Nature 348, 491.
- Woese, C. R. & Fox, G. E. (1977b). The concept of cellular evolution. Journal of Molecular Evolution 10 , 1–6.
- Stanier, R. (1970). Some aspects of the biology of cells and their possible evolutionary significance. Symposium for the Society of Genernal Microbiology 20, 1–38.
- Keeling, P., Charlebois, R. L. & Doolittle, W. F. (1994). Archaebacterial genomes: eubacterial form, eukaryotic content. Current Opinion in Genetics and Development 4, 816–822.
- Cavalier-Smith, T. (1981b). The origin and early evolution of the eukaryotic cell. En Molecular and Cellular Aspects of Microbial Evolution. Society for General Microbiology Symposium 32 (ed. M. J. Carlile, J. F. Collins and B. E. B. Moseley), pp. 33–84. Cambridge University Press.
- Cavalier-Smith, T. (1987c). The origin of eukaryote and archaebacterial cells. Annals of the New York Academy of Sciences 503, 17–54.
- Cavalier-Smith, T. (1989a). Systems of kingdoms. En McGraw Hill Yearbook of Science and Technology, pp. 175–179.
- Cavalier-Smith, T. (1993b). Evolution of the eukaryotic genome. En The Eukaryotic Genome, Organization and Regulation (ed. P. M. A. Broda, S. G. Oliver & P. F. G. Sims ), pp. 333–385. Cambridge University Press.
- Hori, H. T., Itoh, T. & Osawa, S. (1982). The phylogenetic structure of the metabacteria. Zentralblatt für Bakteriologie und Hygiene [C] 3, 18–30.
- Edgell, D. R. & Doolittle, W. F. (1997). Archaea and the origin(s) of DNA replication proteins. Cell 89 , 995–998.
- Ouzonis, C. A. y Kyrpides, N. K. (1996). Hypothesis: the emergence of major cellular processes in evolution. FEBS Letters 399, 119–123.
- Woese, C. R., Kandler, O. y Wheelis, M. L. (1990). Towards a natural system of organisms, proposal for the domains Archaea, Bacteria, and Eucarya. Proceedings of the National Academy of Sciences, USA 87 , 4576–4579.
- Cavalier-Smith T. 2002a The neomuran origin of archaebacteria, the negibacterial root of the universal tree and bacterial megaclassification. Int. J. Syst. Evol. Microbiol. 52, 7–76
- Valas R. E., Bourne P. E. 2009 Structural analysis of polarizing indels: an emerging consensus on the root of the tree of life. Biol. Direct. 4, 30
- Williams, T., Foster, P., Cox, C. et al. An archaeal origin of eukaryotes supports only two primary domains of life. Nature 504, 231–236 (2013).
Enlaces externos
- Woese et al 1990. "Towards a natural system of organisms: Proposal of the domains Archaea, Bacteria and Eucarya" En el sitio web del PNAS
| Control de autoridades |
|
|---|
 Datos: Q2430501
Datos: Q2430501