Sterna hirundo
El charrán común[3] [lower-alpha 1] (Sterna hirundo) es una especie de ave caradriforme de la familia Laridae.[5] Es un ave marina y migratoria que pasa el invierno en las costas de países tropicales y subtropicales. Sus cuatro subespecies anidan en regiones templadas y subárticas de Europa, de Asia y del este y centro de América del Norte. Los individuos adultos tienen las partes superiores de color gris claro y el dorso de blanco a gris claro, el píleo negro, las patas anaranjadas y el pico de punta estrecha. Dependiendo de la subespecie, el pico es rojo con la punta negra o completamente negro. Existen varias especies similares, como el charrán ártico —parcialmente simpátrico—, que se diferencia por los detalles del plumaje, patas y color del pico, o por las vocalizaciones.
| Charrán común | ||
|---|---|---|
_in_Canary_Islands.jpg.webp) Ejemplar en las islas Canarias. | ||
| Estado de conservación | ||
 Preocupación menor (UICN 3.1)[1] | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Aves | |
| Orden: | Charadriiformes | |
| Familia: | Laridae | |
| Género: | Sterna | |
| Especie: |
S. hirundo Linneo, 1758 | |
| Distribución | ||
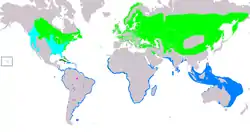 Distribución de Sterna hirundo Área de cría. Área de ocupación permanente. Área de invernada. Área de migración. Vagrant (estacionalidad incierta). | ||
| Subespecies | ||
Véase el texto | ||
| Sinonimia | ||
| ||
El hábitat reproductivo es más extenso y diverso que el de cualquiera de sus parientes. Construye su nido en cualquier sitio —desde superficies con escasa vegetación hasta cerca del agua, incluyendo playas e islas— y se adapta fácilmente a sustratos artificiales como balsas flotantes. El nido es un escarbado al descubierto en la arena o grava, pero también puede revestirlo por dentro o en los bordes con residuos que encuentre disponibles. La nidada consiste de tres huevos de color tenue y con patrones de manchas que proporcionan camuflaje en la playa abierta. La pareja incuba los huevos hasta la eclosión, unos veintiún a veintidós días después, pero este tiempo se prolongaría si la colonia es perturbada por depredadores. Los pichones abandonan el nido en un tiempo de veintidós a veintiocho días. Como la mayoría de charranes, esta especie bucea para cazar peces, ya sea en el mar o en agua dulce, pero los moluscos, crustáceos y otras presas invertebradas son una parte importante de su dieta en algunas zonas.
Los huevos y las crías son vulnerables a la depredación por mamíferos —como las ratas y el visón americano— y aves de mayor tamaño, como gaviotas, rapaces nocturnas y garzas. El charrán común es propenso a sufrir parasitismo por parte de piojos, helmintos, ácaros y hematozoos (este último en menor cantidad). La numerosa población y área extensa de reproducción permiten que esta especie sea clasificada como de «preocupación menor», pero las poblaciones norteamericanas han disminuido drásticamente en las últimas décadas. Pese a la legislación internacional que protege al charrán común, en algunas zonas las poblaciones están amenazadas por la destrucción del hábitat, la contaminación o la perturbación de las colonias de anidación.
Descripción

.jpg.webp)
La subespecie nominal del charrán común (S. h. hirundo) tiene 31-35 cm (centímetros) de longitud, 6.9 cm de la punta de la cola y 77-98 cm de envergadura. Su peso es de 110-141 g (gramos).[6][7] Las partes superiores de los adultos reproductores son de color gris tenue, mientras que las inferiores poseen una tonalidad gris muy clara; tienen un píleo negro, patas de color anaranjado rojizo y un pico con punta estrecha (la coloración de este último depende de la subespecie, ya que puede ser completamente negro o de color rojo con la punta negra).[8] Las plumas superiores son gris claro, pero a medida que llega el verano los cañones[lower-alpha 2] oscuros de las plumas de vuelo externas quedan expuestos al aire y aparece en las alas una porción grisácea. El obispillo y la cola son blancuzcos y si el ave está de pie la longitud de la cola no sobrepasa los extremos de las alas plegadas, a diferencia del charrán ártico y rosado, en los que la cola sobresale más allá de las alas. No se observan diferencias significativas entre ambos sexos.[9] En adultos no reproductores, la frente y las partes inferiores son blancas, el pico es negro en la punta y con una base de color rojizo —o completamente negro— y las patas son de color rojo oscuro o negro.[9] Las plumas superiores tienen una zona oscura en el borde delantero del ala, denominada banda carpiana. Las aves que se no han desarrollado apropiadamente pueden sufrir una muda del plumaje de adulto no reproductor en junio, pero en los individuos bien desarrollados ocurre a finales de julio, con una muda interrumpida durante la migración. Existen algunas variaciones geográficas: por ejemplo, los pájaros californianos suelen tener plumaje de adulto no reproductor durante la migración.[6]


Las plumas superiores de los jóvenes son de color gris claro con una banda carpiana oscura. El píleo y la nuca son marrones y la frente tiene un tono rojizo, que cambia a blanco en otoño. Las plumas de las partes superiores son rojizas con líneas marrones o blancas y la cola carece de las plumas largas de los adultos.[6] Normalmente, las aves con su primer plumaje posjuvenil permanecen en sus áreas de invernada y se parecen a los adultos no reproductores, pero tienen píleo más oscuro, banda carpiana oscura y plumaje más desgastado. Al segundo año de vida, la mayoría de los charranes jóvenes son prácticamente indistinguibles de los adultos o solo muestran diferencias menores, como un pico más oscuro o una frente blanca.[10]
El charrán común es un volador ágil, capaz de realizar giros y descensos rápidos en el cielo, planeos y despegue vertical. Cuando atrapa peces vuela cerca de la superficie acuática con un fuerte viento en contra, pero luego asciende 10-30 m (metros) sobre el agua con un viento favorable. Por lo general, vuela por debajo de los 100 m y a un promedio de 30 km/h (kilómetros por hora) en ausencia de viento favorable; estos valores no son los mismos durante la migración.[11] La velocidad promedio de vuelo durante el viaje migratorio diurno es 32-43 km/h[12] y en la noche es de 43-54 km/h[13] a una altura de 1000-3000 m.[11]
Muda

En octubre los jóvenes inician una muda a su primer plumaje adulto. Al siguiente año, durante del mes de febrero, el plumaje de la cabeza, la cola y la parte inferior del cuerpo son sustituidos por completo, seguidos de las alas. Las plumas primarias son remplazadas por etapas: primero las más internas, pero el proceso se interrumpe durante el invierno austral —a esta edad, las aves permanecen en sus áreas de invernada— y reinicia en el otoño austral. En el segundo año de vida (entre mayo y junio) comienza una nueva secuencia de muda, con una pausa de la muda principal en las aves que regresan al norte, pero no en aquellas que se quedan en las zonas de invernada. También se produce una muda importante al plumaje de adulto entre febrero y junio del siguiente año, en la que el 40-90 % de las plumas son reemplazadas.[6] Las primarias se desgastan hasta revelar las barbas negruzcas más próximas al raquis. El patrón de muda comienza en el plumaje más antiguo que está más cerca del centro del ala, por lo que a medida que llega el verano boreal aparece una cuña de tono oscuro en la parte superior del ala debido al envejecimiento de las plumas.[8]
La frecuencia de renovación de las plumas primarias es distinta a los otros charranes, pues las remplazan al menos dos o tres veces en un año. Las diferencias visibles según la edad del plumaje se acentúan por la mayor reflectancia ultravioleta de las nuevas plumas primarias, la que es utilizada por las hembras al momento de seleccionar a su pareja.[14] Las hembras más maduras tienden a buscar machos que exhiben una buena eficacia biológica a través la calidad de las plumas de las alas.[15] En raras ocasiones una muda muy precoz en la colonia nidificante está relacionada con una insuficiencia reproductiva, pues tanto el inicio de la muda como el comportamiento reproductivo están vinculados con una disminución de los niveles de la hormona prolactina.[16]
Vocalizaciones
El ave tiene un amplio repertorio de llamadas, así como un tono más grave que las de los charranes árticos. El sonido más distintivo es una llamada de alarma transcrita fonéticamente como kee-yah, que destaca la primera sílaba. Con el charrán ártico sucede lo contrario, pues este enfatiza la segunda sílaba en lugar de la primera.[17] La llamada de alarma funciona como una advertencia a los intrusos, aunque ante serias amenazas emite un kyar y el ave regresa rápidamente para tranquilizar a la colonia, habitualmente bulliciosa, mientras los charranes evalúan el peligro.[17] Puede pronunciar un keeur grave y borroso cuando un adulto se acerca al nido con un pez en el pico y es posible que lo utilice para la identificación individual (por ejemplo, cuando los polluelos salen de su escondite al escuchar a sus padres enunciando esta llamada).[17] Otra llamada común es un kip pronunciado durante el contacto social. Otras vocalizaciones son un kakakakaka en los ataques a intrusos y un entrecortado kek-kek-kek en las luchas que los machos sostienen entre sí.[17] Los padres y los polluelos pueden usar las llamadas para indicar su ubicación; los hermanos también se reconocen entre ellos mediante vocalizaciones alrededor de doce días después de la eclosión, lo que ayuda a mantener juntas a las crías.[18][19]
Taxonomía
_-_Sterna_hirundo_(106b).jpg.webp)

Los charranes o golondrinas de mar son aves marinas de tamaño pequeño a mediano estrechamente relacionadas con las gaviotas (grupo polifilético dentro de la familia Laridae), rayadores (Rynchops)[5] y págalos (Stercorariidae). En apariencia son similares a las gaviotas, pero a grandes rasgos tienen una estructura anatómica más ligera, alas largas y puntiagudas (que les proporcionan un vuelo rápido y ligero), cola con una bifurcación profunda, patas delgadas[20] y dedos con membranas interdigitales.[21] La mayoría de los charranes son grises por encima y blancos por debajo y tienen un píleo negro que se llena o motea de manchas blancas en la temporada no reproductiva.[20] Aparentemente los parientes más cercanos son los gaviotines antárticos (Sterna vittata),[11] seguidos de los charranes ártico y rosado. La evidencia genética sugiere que el charrán común pudo haberse separado de la línea ancestral antes que sus parientes.[22] No existen fósiles identificados en América del Norte y los encontrados en Europa son de edad y especie inciertas.[11]
El charrán común fue descrito por Carlos Linneo en su Systema naturæ (1758) bajo su actual nombre científico, Sterna hirundo.[23] La palabra stearn se usaba en el inglés antiguo y un vocablo similar era utilizado por los frisones para designar a la golondrina de mar.[24] Una de las menciones más antiguas aparece en el poema The Seafarer (El navegante),[lower-alpha 3] escrito alrededor del año 1000. Linneo adoptó esta palabra para el nombre del género: Sterna. La palabra latina para golondrina es hirundo y, en este caso en particular, se aplica al charrán por su semejanza superficial con aquella ave no relacionada, la cual tiene una contextura similar y una cola larga bifurcada.[26] Esta semejanza también originó el nombre «golondrina de mar»,[27] documentado en la literatura inglesa desde al menos el siglo xvii.[26] Se cree también que los nombres escoceses picktarnie,[28] tarrock[29] y sus muchas variantes son de origen onomatopéyico, es decir, derivados de la llamada del ave.[26] A causa de la dificultad en diferenciar entre el charrán común y la golondrina, la mayoría de los nombres vulgares son compartidos con el charrán ártico.[30] Cuatro subespecies del charrán común son reconocidas por la mayoría de autores, aunque a veces S. h. minussensis se considera un intergrado entre S. h. hirundo y S. h. longipennis.[31][32]
| Subespecies | Zonas de anidación[33][34] | Características principales | |
|---|---|---|---|
| Nombre trinomial | Autoridad | ||
| S. h. hirundo | Linnaeus, 1758[23] | Europa, norte de África, desde Asia Oriental hasta Siberia occidental y Kazajistán, América del Norte.[6] | Las diferencias entre las poblaciones americana y eurasiáticas son mínimas. En promedio, las alas de las aves americanas tienen una longitud ligeramente más corta y la extensión de la punta oscura en la mandíbula superior tiende a ser menor que en las aves de los países escandinavos y del extremo este de Eurasia. La proporción de negro en el pico es mínima en el oeste de Europa, por lo que los adultos reproductores británicos son muy similares a las aves americanas a este aspecto.[6] |
| S. h. minussensis | Sushkin, 1925[35] | Zona oriental del lago Baikal hasta el norte de Mongolia y el sur de Tíbet.[36] | La parte superior del cuerpo y las alas son más pálidas que las de S. h. longipennis y tiene un pico carmesí con la punta negra. |
| S. h. longipennis | Nordmann, 1835[37] | Siberia central hasta China, también en Alaska.[6] | Tiene el plumaje con un tono gris más oscuro que la subespecie nominal, pico negro más corto, patas de color marrón rojizo oscuro y alas más largas.[6] |
| S. h. tibetana | Saunders, 1876[37] | Desde los Himalayas hasta el sur de Mongolia y China.[6] | Es similar a la subespecie nominal, pero el pico es más corto y con la punta negra y más amplia.[6] |
Especies similares
%252C_Ward_of_Clugan_-_geograph.org.uk_-_1357999.jpg.webp)
Varios miembros de Sterna poseen un tamaño y aspecto similares al charrán común.[38] Una especie habitualmente difícil de diferenciar es el charrán ártico (S. paradisaea) y, hasta que se definieron las características esenciales, ambas aves eran confundidas durante el vuelo o desde la distancia; también eran conocidas en forma conjunta como «charranes commic». Aunque son similares en tamaño,[39] ambos charranes difieren en estructura anatómica y vuelo. El charrán común tiene la cabeza más grande, un cuello más grueso, patas más largas y triangulares, alas más rígidas que su pariente y vuelo más potente y recto.[40] Por su lado, el charrán ártico tiene partes inferiores más grisáceas que el charrán común y sus mejillas blancas son más evidentes. El obispillo del charrán común puede ser grisáceo en el plumaje de adulto no reproductor, en comparación con el blanco puro de su pariente. El charrán común desarrolla una porción oscura en las alas cuando se acerca la temporada de reproducción, pero las alas del charrán ártico se tornan blancas durante todo el verano boreal. Desde tierra, las plumas de vuelo del charrán ártico se observan translúcidas con cielo brillante; sin embargo, solamente cuatro de las plumas más internas de las alas del charrán común poseen esta característica.[40][41] En el charrán ártico el borde posterior de las plumas de vuelo exteriores tiene una línea fina y negra, pero se observa más gruesa y menos definida en S. hirundo.[9] Un charrán común adulto tiene el pico rojizo anaranjado con una punta negra, excepto S. h. longipennis que tiene el pico negro y patas de color rojo brillante; en el charrán ártico las patas y el pico son de un rojo más oscuro y carece del extremo negro en el pico.[40]
En las zonas de anidación, el charrán rosado (S. dougallii) puede distinguirse por su plumaje claro, pico largo y negro y plumas largas en la cola.[41] El plumaje de adulto no reproductor es claro por encima y blanco por debajo —a veces con tonos rosáceos—, tiene el pico negro y se observan unas plumas oscuras bajo su cola blanca.[42] Durante el vuelo el charrán rosado se caracteriza por su cabeza y cuello grandes, pico largo y aleteos rápidos y rígidos.[43] Busca su alimento en el mar con mayor frecuencia que el charrán común.[42] En América del Norte, el charrán de Forster (S. forsteri) posee plumaje nupcial que lo hace aparentemente más grande que S. hirundo, también tiene las alas relativamente más cortas, cabeza considerablemente grande, pico grueso y patas largas y fuertes; en su plumaje de adulto no reproductor la cabeza es blanca con una mancha ocular oscura, algo que lo hace inconfundible de las demás aves americanas.[44] En las zonas de invernada también existe confusión en la diferenciación de las aves, con el charrán antártico (S. vittata) en los océanos australes, el charrán sudamericano (S. hirundinacea), el charrán maorí (S. striata) en Australasia y el charrán arábigo (S. repressa) en el océano Índico. Para la identificación se pueden tener en cuenta las diferencias del plumaje debido a las temporadas de reproducción «opuestas». El charrán antártico es más robusto que S. hirundo y con un pico más grueso. En el plumaje del adulto reproductor las partes inferiores son oscuras y el píleo es completamente negro y delineado con una línea blanca sobre las mejillas, pero el plumaje del adulto no reproductor carece, o está poco definido, de la banda carpiana y las aves jóvenes presentan franjas oscuras en las plumas terciarias que son visibles cuando el ala está cerrada o durante el vuelo.[45][46] El charrán sudamericano es más grande que S. hirundo, con un pico rojizo más curvo y grande y tiene un píleo negro más amplio y uniforme en el plumaje de adulto no reproductor.[47] Al igual que S. vittata, el charrán sudamericano carece de banda carpiana en el plumaje de adulto no reproductor y también comparte la mencionada característica de las plumas terciarias en las aves jóvenes.[48] La frente del charrán maorí es blanca en el plumaje de adulto reproductor y cuenta con un pico más grueso; el plumaje del adulto no reproductor es más claro en la parte inferior que S. hirundo y las partes inferiores de las alas son blancas.[49] El charrán arábigo es más pequeño, tiene partes superiores grises y lisas, el plumaje del adulto reproductor es más oscuro en la parte superior y las mejillas son más blancas.[50]
Los jóvenes charranes comunes se diferencian fácilmente de otras especies relacionadas de edad similar, pues presentan una coloración rojiza en la parte posterior de la cabeza y una base clara en el pico. En tanto, los charrares árticos jóvenes pueden desarrollar un dorso gris y un pico negro y los charranes rosados juveniles tienen una peculiar mancha en forma de silla de montar en tono sepia en el dorso.[9] Se han documentado hibridaciones entre charranes comunes y rosados —en particular, provenientes de los Estados Unidos— y el plumaje intermedio y las llamadas observados en estas aves son una dificultad potencial para la identificación. Estas aves pueden tener un pico casi completamente negro, pero la confirmación de una reproducción mixta puede depender de detalles específicos de las plumas de vuelo individuales.[6]






Distribución y hábitat

La mayoría de las poblaciones presenta una conducta migratoria, pues en el hemisferio norte viajan a regiones más al sur de sus zonas de anidación de clima templado y subártico para pasar el invierno. En su primer verano las aves jóvenes suelen permanecer en sus zonas de invernada, aunque algunas retornan a las colonias de anidación en algún momento después de la llegada de los adultos.[10] En América del Norte el charrán común anida a lo largo de la costa atlántica de Labrador hasta Carolina del Norte y hacia el interior atravesando gran parte de Canadá hasta el extremo oriental de las Rocallosas. En los Estados Unidos también pueden encontrarse algunas poblaciones nidificantes en los estados fronterizos de los Grandes Lagos y, de manera limitada, en la costa del golfo de México.[51] Las colonias del Caribe realizan migraciones pequeñas y parciales. Estas poblaciones se encuentran en las Bahamas, Cuba[52] y en los archipiélagos de Los Roques y Las Aves en las aguas venezolanas.[53] En el continente americano las aves pasan el invierno en las costas atlántica y pacífica de América Central y del Sur, en las costas de la Argentina y en el litoral norte de Chile.[10][51] Los registros provenientes de América del Sur y las Azores indican que algunas aves pueden cruzar el Atlántico en ambos sentidos en su ruta migratoria.[54][55]
El charrán común anida en la mayor parte de Europa y las poblaciones más grandes se encuentran en el norte y este del continente. Existen pequeñas poblaciones en la costa norte de África y en las Azores, Canarias y Madeira. La mayoría de los grupos mencionados pasa el invierno cerca del África Occidental o Meridional, si bien las aves provenientes del sur y oeste de Europa tienden a permanecer al norte del ecuador, otras poblaciones migran más hacia el sur.[56] La zona de anidación atraviesa las zonas templadas y la taiga en Asia, con asentamientos dispersos en el golfo Pérsico y la costa de Irán.[57] Unas poblaciones más pequeñas anidan en las islas cercanas a Sri Lanka[58][59] y en la región de Ladakh de la meseta del Tíbet.[60] Las aves de Asia occidental invernan en el norte del océano Índico;[10][61] la subespecie S. h. tibetana llega cerca de África Oriental durante el invierno boreal.[62][63] Las aves que habitan los extremos norte y este de Asia, como S. h. longipennis, migran a través de Japón, Tailandia y el Pacífico occidental hasta el sur de Australia.[10] Existen colonias pequeñas y erráticas en el África Occidental, específicamente en Nigeria y Guinea-Bisáu, pero son no son frecuentes porque están en una zona de invernada.[57] Solo se han observado unos cuantos charranes comunes en Nueva Zelanda[64] y la presencia de esta especie en la Polinesia no está completamente esclarecida.[65]
Un charrán anillado en su nido en Suecia fue encontrado muerto cinco meses más tarde en la isla Stewart después de haber volado aproximadamente 25 000 km(kilómetros).[66] Algunas veces, en las migraciones a larga distancia, el charrán común llega a sitios más allá de su distribución geográfica habitual. Se han encontrado aves perdidas en el interior de África (Zambia y Malaui), las Maldivas y las Comoras;[67] la subespecie nominal, por ejemplo, ha aparecido en Australia,[49] los Andes y en el interior de América del Sur.[47][68] También hay avistamientos recientes de S. h. longipennis (endémica de Asia) en Europa occidental.[69]

Esta ave reside en una mayor cantidad de hábitats que cualquiera de sus parientes, pues anida desde la taiga asiática hasta las costas tropicales[70] y en altitudes de 2000 m s. n. m. (metros sobre el nivel del mar) en Armenia y hasta 4800 m s. n. m. en Asia.[56] Evita los lugares frecuentemente expuestos al exceso de lluvia o viento y los cuerpos de agua congelada, por lo que no anida tan al norte como el charrán ártico. Prefiere anidar cerca de lagos de agua dulce o en el mar, en casi cualquier hábitat de terreno llano y abierto, como playas de arena o guijarros, zonas de dunas firmes, marismas salinas o, más comúnmente, islas. Los brezales o pastizales llanos, o incluso grandes rocas planas, pueden resultar adecuados en un entorno isleño.[70] En colonias mixtas tolera vegetación de mayor altura durante más tiempo que el charrán ártico, pero de menor altura que la tolerable por el charrán rosado; en este caso, el factor relevante es la longitud diferente de las patas de las tres especies mencionadas.[71] El charrán común se adapta fácilmente a las balsas flotantes artificiales e incluso puede anidar en el tejado plano de las fábricas.[70] Puede asentar su nido en lugares poco regulares, como fardos de heno, tocones que sobresalen 0.6 m (metros) sobre el agua y troncos o vegetación flotantes. Existe el caso documentado de un charrán común que se adueñó del nido de un playero manchado y puso sus huevos junto a los del ave limícola.[72] Fuera de la temporada de reproducción todo lo que necesita en cuanto al hábitat es un acceso a sitios para pescar y algún lugar para aterrizar. Aparte de las playas y rocas naturales, utiliza muchas veces los barcos, boyas y muelles como posaderos y perchas nocturnas.[70]
Comportamiento
Territorialidad


.jpg.webp)
El charrán común anida en colonias que normalmente no superan las 2000 parejas,[56] pero en ocasiones pueden contar más de 20 000.[73] Las colonias tienden a ser más pequeñas tierra adentro que en la costa. Casi siempre anida junto a otras especies costeras, como los charranes ártico,[74] rosado y patinegro (Thalasseus sandvicensis); la gaviota reidora (Chroicocephalus ridibundus)[75][76] y el rayador americano (Rynchops niger).[77] Por alguna razón desconocida, la mayor parte o la totalidad de los charranes vuela silenciosa y rápidamente hacia el mar. A este fenómeno se denomina «pavor» y ocurre sobre todo en la primera etapa de la temporada de reproducción.[56]
A su llegada a las zonas de cría, puede merodear durante unos días antes de establecerse en un territorio[78] y el verdadero inicio de la nidada puede estar relacionado con una alta disponibilidad de presas.[79] Únicamente defienden una zona pequeña y las distancias entre nidos pueden ser de solo 50 cm (centímetros), aunque lo habitual es que se hallen separados de 150 a 350 cm entre sí. Al igual que muchas aves, el sitio es reutilizado cada año; por ejemplo, una pareja regresó al mismo lugar en diecisiete temporadas reproductivas sucesivas. Alrededor del 90 % de las aves experimentadas reutilizaron su antiguo territorio, mientras que las aves jóvenes anidaron en la periferia, encontraron un compañero desconsolado o se trasladaron a otra colonia.[78] El macho selecciona un territorio de anidación pocos días después de su llegada en la primavera y se une a su hembra anterior, excepto si ella llega con más de cinco días de retraso, en cuyo caso se desentiende de la pareja.[80]

El macho se ocupa principalmente de la defensa del territorio y repele a otros charranes intrusos. Cuando se acerca algún extraño, emite una llamada de alarma, abre sus alas, levanta la cola e inclina la cabeza para mostrar su píleo negro. Si el intruso persiste, el macho interrumpe la llamada y ataca usando su pico hasta que el intruso levanta la cabeza exponiendo su garganta en señal de sometimiento. Los intrusos aéreos simplemente son atacados, a veces con un ascenso en espiral.[78] Si bien se observa agresión a pájaros adultos, los polluelos errantes suelen ser tolerados, aunque en una colonia de gaviotas estos serían atacados y asesinados. El nido es defendido hasta que los polluelos han emplumado y todos los adultos de la colonia ahuyentarán colectivamente a depredadores potenciales.[81]
Reproducción
Las parejas se consolidan luego de un cortejo de exhibiciones aéreas en las que el macho y la hembra vuelan en amplios círculos a una altura de aproximadamente 200 m (metros) y con cantos a cada rato, antes de que ambos desciendan en planeos en zigzag. Si el macho lleva un pez en el pico, puede llamar la atención de otros machos también. Si están en tierra, el macho corteja a la hembra caminando alrededor de ella con su cuello y cola alzados, la cabeza apuntando hacia abajo y las alas parcialmente abiertas. Si ella le corresponde, ambos adoptan una postura con la cabeza señalando hacia el cielo. El macho puede provocar a la hembra con un pez, que no le da hasta que ella le ha suplicado lo suficiente.[82] Una vez que ha terminado el cortejo, el macho cava un agujero poco profundo en la arena y la hembra escarba en el mismo sitio. Tras varios intentos, la pareja tendrá un sitio preparado para el nido.[82] La hembra pone sus huevos sobre la arena desnuda, grava o tierra, pero puede usar un revestimiento de residuos o vegetación si está disponible[56] o forrar el nido con algas, piedras o conchas. El agujero en forma de plato tiene aproximadamente 4 cm (centímetros) de profundidad y 10 cm de diámetro, pero se puede extender hasta 24 cm, incluyendo el material decorativo circundante.[83] El éxito reproductivo en las regiones propensas a inundaciones ha mejorado con la colocación de esteras artificiales hechas de crin marina que fomentan el anidamiento de charranes en áreas más elevadas y menos vulnerables, pues muchos prefieren estas esteras a la arena desnuda para asentar su nido.[84] El charrán común usa más material en los nidos que los charranes rosados o árticos, aunque muchas veces el charrán rosado anida en zonas con mayor vegetación.[85][86] El ave es experta en localizar su nido en una colonia extensa. Los estudios demuestran que el charrán común es capaz de encontrar sus huevos cuando están enterrados, incluso si se quita material del nido o los cubre la arena. Pueden localizar su nido a 5 m de su sitio original —incluso si se traslada en varias etapas— y aceptan los huevos si son remendados con plastilina o coloreados con amarillo, pero no con rojo o azul. Esta capacidad de localizar sus huevos podría ser una adaptación a la vida en un entorno inestable, al viento y las mareas.[71]

El apogeo de la producción de huevos se da a principios de mayo, pero en algunas aves, particularmente los adultos en su primera vez, ocurre a finales de dicho mes o en junio.[72][83] Normalmente el tamaño de la puesta es de tres huevos y es probable que las nidadas de mayor tamaño sean el resultado de dos hembras que ponen en el mismo nido.[83] El tamaño promedio de cada huevo es 41 × 31 mm (milímetros), aunque cada huevo sucesivo de una puesta es ligeramente más pequeño que el primero; el peso promedio es de 20.2 g (gramos), de los cuales el 5 % corresponde al cascarón.[87] El peso del huevo depende de lo bien alimentada que esté la hembra, así como de su postura en la puesta. Los huevos son de color crema, ocre pardo o marrón claro, con rayas, puntos o manchas negras, marrones o grises que ayudan a camuflarlos.[83] La pareja realiza la incubación —aunque la hembra se hace cargo de ella por más tiempo— y tiene una duración de aproximadamente de 21 a 22 días, extiendiéndose a 25 días si hay distubios frecuentes en la colonia que obliguen a los adultos a dejar los huevos sin vigilancia;[83] la depredación nocturna puede conducir a que la incubación dure hasta 34 días.[72] En los días calurosos el padre sale en busca de agua para humedecer las plumas de su vientre y regresa de nuevo a la incubación, lo que ofrezce a los huevos cierto enfriamiento.[11] A menos que la colonia sufra algún desastre, el 90 % de los huevos consigue eclosionar.[88] El polluelo es precoz, presenta un fino plumón amarillento con manchas negras o marrones[83] y, así como los huevos, sus etapas de crecimiento son similares a las de los charranes árticos.[89] Abandonan el nido a los 22-28 días,[87] generalmente entre los días 25 y 26.[56] Los polluelos siguen recibiendo alimento en el nido durante unos cinco días más y después acompañan a los adultos en expediciones de pesca; también pueden recibir alimentos suplementarios de los padres hasta el final de la temporada de reproducción o más. Se tiene constancia de que el charrán común alimenta a sus crías durante la migración y en las zonas de invernada, al menos hasta que los adultos se trasladan más hacia el sur durante el mes de diciembre.[11][90]
.jpg.webp)
Como muchos miembros del género Sterna, esta especie protege su nido y polluelos y ahuyenta a humanos, perros, ratas almizcleras y una gran variedad de aves diurnas, pero a diferencia del más agresivo charrán ártico, rara vez agrede al intruso y suele desviarse bruscamente en el último momento. Las aves adultas son capaces de distinguir a cada humano y atacan con mayor intensidad a las personas con las que están más familiarizadas que a los extraños.[91] Los depredadores nocturnos no sufren agresiones similares;[92] las colonias pueden ser devastadas por las ratas y los adultos abandonan el lugar durante casi ocho horas si hay búhos cornudos en las cercanías.[93]
Por lo general, la pareja se reproduce una vez al año y posiblemente tengan una segunda puesta si se pierde la primera. En raras ocasiones, la pareja puede incubar una segunda puesta al mismo tiempo que alimenta algunos polluelos de la primera.[94] Normalmente el primer intento reproductivo ocurre a los cuatro años de edad, a veces a los tres. A nivel mundial, el promedio de jóvenes por pareja que sobrevive hasta el emplumecimiento puede variar de cero —en caso de que la colonia se inunde— a más de 2.5 en un buen año. En América del Norte, la media está entre 1.0 y 2.0 en las islas, pero es inferior a 1.0 en los litorales y tierra adentro. Con la edad las aves adquieren mayor eficiencia en la crianza de sus polluelos, esto se observa durante toda la etapa reproductiva y alcanza su máximo en los primeros cinco años.[11][89] La edad máxima documentada en la naturaleza es de 23 años en América del Norte[95][96] y 33 en Europa,[97][98] si bien la edad más común es de 12 años.[87]
Alimentación


Como los demás charranes, S. hirundo captura a sus presas zambulléndose en picado desde una altura de 1 a 6 m (metros), tanto en el mar como en lagos y ríos. La inmersión dura alrededor de un segundo, pero nunca desciende más de 50 cm (centímetros) bajo el agua.[99] Cuando busca peces vuela mirando hacia abajo con su pico apuntando verticalmente a la superficie del agua; también puede volar en círculos o cernirse brevemente antes de zambullirse en el agua. Por su parte, el charrán ártico emplea una técnica conocida como «planeo escalonado» (stepped-hover)[100] [lower-alpha 4] y el charrán rosado se sumerge rápidamente desde una mayor altura y durante más tiempo.[101] El charrán común puede buscar alimento hasta 5.10 km (kilómetros) de distancia de la colonia de anidación, en ocasiones hasta 15 km.[102] Perseguirá a los bancos de peces y su ruta de migración en África occidental se ve afectada por el movimiento de grandes cardúmenes de sardinas frente a las costas de Ghana; también seguirá los grupos de peces depredadores[lower-alpha 5] o delfines y esperará que su presa sea conducida hasta la superficie del mar.[102][103] Habitualmente se alimenta en bandadas —en especial si la comida es abundante— y el resultado habitual de la pesca colectiva es aproximadamente un tercio mayor que el de la pesca individual.[99]
En el interior de los conos de la retina de sus ojos posee unas gotitas de aceite rojizo. Estas mejoran el contraste y agudizan la visión a distancia, sobre todo en condiciones brumosas.[104] Las aves que necesitan ver a través del plano de contacto aire-agua, como las gaviotas y los charranes, tienen gotitas de aceite coloreadas con pigmentos carotenoides en mayor intensidad que otras especies.[105] Esta vista mejorada ayuda a los charranes a localizar cardúmenes, aunque no se sabe si pueden percibir el fitoplancton del que se alimentan los peces u observan a otros charranes buceando en busca de alimento.[106] La mayoría de miembros de las familias Sternidae y Laridae no cuenta con ojos particularmente sensibles a los rayos ultravioleta, una adaptación más adecuada para aves que se alimentan en tierra como las gaviotas.[107]

El tamaño más frecuente de las peces que captura va desde los 5 a los 15 cm (centímetros) de longitud.[72][99] Las especies capturadas varían según lo que encuentre disponible en el mar, pero cuando es posible elegir los charranes con nidadas más numerosas capturan presas de mayor tamaño que los que tienen nidadas más pequeñas.[108] La proporción de polluelos alimentados con peces puede llegar al 95 % en algunas zonas, pero los invertebrados pueden constituir una parte importante de la dieta, en la que se incluyen: gusanos, sanguijuelas y moluscos, como calamares pequeños y crustáceos (gambas, quisquillas y cangrejos topo). En zonas de agua dulce pueden capturar insectos grandes como escarabajos, gusanos blancos y polillas; también atrapan insectos adultos en el aire y recogen larvas del suelo o sobre la superficie del agua. La presa queda atrapada en el pico y la traga de cabeza o se la lleva a los polluelos. En raras ocasiones transporta dos o más peces pequeños en el mismo viaje.[99] Cuando los adultos regresan con alimento al nido reconocen a sus crías por las llamadas en lugar de por la identificación visual.[19]
Cuando conviven juntos, el charrán común roba los peces capturados por el charrán ártico,[109] así como sufrir cleptoparasitismo por págalos,[110] gaviotas reidoras americanas,[111] charranes rosados[112] o por otras aves de su misma especie mientras regresa con peces a su nido.[109][113] En una investigación, dos machos cuyas parejas habían muerto lograron sacar adelante su nidada robando comida de los nidos vecinos.[114] S. hirundo es capaz de beber mientras vuela y habitualmente toma agua de mar en lugar de agua dulce, si ambas están disponibles.[11] Los polluelos no beben antes del emplumecimiento, sino que reabsorben agua y, al igual que los adultos, excretan el exceso de sal en una solución concentrada por una glándula nasal especializada.[115][116] Los huesos de peces y los exoesqueletos duros de crustáceos o insectos son regurgitados en forma de egagrópilas. Los adultos vuelan del nido para defecar, inclusive los pequeños polluelos salen para depositar sus heces en un agujero escarbado a poca distancia del nido. Cuando los adultos atacan a animales o seres humanos suelen defecar en el momento que descienden en picado y casi siempre tienen éxito para ensuciar al intruso.[11]
Depredadores y parásitos

Existe una amplia variedad de animales terrestres implicados en ataques a individuos adultos y sus colonias. Por ejemplo, en América del Norte se ha reportado que las ratas roban huevos y los acaparan en grandes cantidades en escondites;[117] y el visón americano (especie introducida en Escocia)[88] y el zorro rojo[118] son importantes depredadores de polluelos indefensos.[119] En diferentes zonas geográficas, otros animales terrestres que en mayor o menor medida atacan las colonias son los mapaches boreales, mofetas rayadas, comadrejas andinas, ardillas arborícolas, perros domésticos, gatos salvajes, reptiles y hormigas.[119] En África Occidental se han reportado humanos consumiendo los huevos y destruyendo parte de las colonias buscándolos.[120] Debido a que los charranes comunes suelen anidar en islas, los depredadores más comunes suelen ser otras aves en lugar de mamíferos. Por ejemplo, el vuelvepiedras común depreda los nidos sin vigilancia;[121][122] las gaviotas se llevan a los polluelos;[123][124] los búhos cornudos y campestres matan a los adultos y polluelos; el martinete común se alimenta de los pequeños polluelos;[11][125] los esmerejones y halcones peregrinos agreden a los individuos adultos mientras vuelan. Aparentemente, los charranes logran confundir a los depredadores que vuelan rápido si se congregan en bandadas.[81]
Entre los parásitos están los piojos de las plumas, que son muy diferentes de los que se encuentran en los charranes árticos pese a la estrecha similitud con S. hirundo.[126] También algunos helmintos —como los miembros del género Diphyllobothrium, Ligula intestinalis (un parásito encontrado principalmente en patos), Schistocephalus (descubierto inicialmente en los peces) y cestodos de la familia Cyclophyllidea— han infestado a los charranes comunes. El ácaro Reighardia sternae apareció en charranes comunes de Italia, América del Norte y China.[127] Un estudio de 75 charranes comunes en etapa reproductiva encontró que ninguno portaba parásitos en la sangre.[128] Se han observado las colonias afectadas por el cólera aviar y ornitosis[11] y es posible que el charrán común esté amenazado en el futuro por brotes de gripe aviaria a los que ya es susceptible.[102] En 1961, fue la primera especie de aves silvestre identificada como infectada por la gripe aviar y se encontró la variante H5N3 en un brote de aves sudafricanas.[129]
Estado de conservación
A nivel mundial está catalogada como una especie bajo preocupación menor en la Lista Roja de la UICN.[1] En 2006 contaba con una población aproximada de 1 600 000-4 600 000 individuos adultos y su zona de anidación abarca una superficie de unos 29 200 000 km² (kilómetros cuadrados); se han estimado 250 000-500 000 parejas reproductoras, la mayoría en Asia.[1] Unas 140 000 parejas anidan en Europa (estimación del año 2000)[130] y casi 80 000 lo hacen en América del Norte (1983), la mayoría en la costa noreste del Atlántico[131] y región de los Grandes Lagos, donde se asienta una población de casi 10 000 parejas en franco declive (2003).[132] En el siglo xix el uso de las plumas y alas de los charranes en los sombreros fue la causa fundamental de la reducción en las poblaciones europeas y norteamericanas, sobre todo en las costas del Atlántico y hacia el interior. A veces se empleaban charranes enteros disecados para hacer sombreros. Las poblaciones se recuperaron a principios del siglo xx, debido principalmente a las nuevas leyes y a la labor de las organizaciones de conservación ambiental.[11][118] Aunque las poblaciones euroasiáticas son estables, las norteamericanas se han reducido en más del 70 % en los últimos 40 años y existe una tendencia negativa en las estimaciones mundiales de esta especie.[102]
Las principales amenazas son la pérdida de hábitat por la construcción de viviendas costeras, el crecimiento de la vegetación o la perturbación de las aves en cortejo por seres humanos, vehículos, barcos o perros. Las inundaciones pueden conducir a pérdidas de nidos y algunas colonias son vulnerables a la depredación por ratas y gaviotas, estas últimas también compiten con los charranes por los lugares de anidación. En el Caribe algunas aves son cazadas para su venta comercial como alimento.[102] El éxito reproductivo puede mejorar si se usan balsas flotantes, islas artificiales u otros nidos artificiales y al impedir la perturbación humana. Otras intervenciones realizadas por el hombre consisten en la quema de la vegetación para despejar el terreno y el exterminio de las gaviotas con el fin de desalentar sus incursiones.[102] El uso de policlorobifenilos (PCB) dio lugar a niveles altos de feminización[lower-alpha 6] en embriones masculinos —que parecía desaparecer antes de emplumecer—, pero no afecta la productividad de la colonia;[135] sin embargo, el diclorodifenildicloroetileno (DDE), que resulta de la degradación del DDT, condujo a niveles muy bajos en el éxito reproductivo en algunos lugares en los Estados Unidos.[11]
La especie se ve particularmente afectada por el cambio climático. Un equipo de investigación de la Agencia de Medio Ambiente del Reino Unido y de la Real Sociedad para la Protección de las Aves estudió el futuro desarrollo de la distribución europea de las aves de cría sobre la base de los modelos climáticos y concluyó que para finales del siglo xxi el área del charrán común se encogerá considerablemente y se trasladará hacia el norte. Ponen en evidencia que el área de distribución en Europa Oriental se fragmentó de manera significativa y que han aparecido nuevas zonas de cría en Islandia y en partes de Nueva Zembla, pero no pueden compensar el territorio geográfico perdido.[136] El charrán común es una de las especies protegidas en el Acuerdo sobre la Conservación de las Aves Acuáticas Migratorias Afrourasiáticas (AEWA) y el Tratado de Aves Migratorias de 1918.[137][138] Los países participantes del AEWA están obligados a cumplir con las estrategias de conservación descritas en el plan de acción. Este tiene por objetivo abordar cuestiones como la conservación de la especie y sus hábitats y la gestión, investigación, educación e implementación de las actividades humanas.[139] Las legislaciones norteamericanas son similares, pero hace mayor énfasis en la protección.[140]
Fuentes
Notas
- Charrán común es el nombre recomendado por la SEO para España (también es conocido así en Costa Rica, México, Nicaragua y la República Dominicana), si bien en países americanos tiene otros nombres: gaviotín pico grueso (en Argentina), gaviotín golondrina (Argentina y Uruguay), gaviotín boreal (Chile), gaviotín común (Colombia y Perú), charrán piquinegro (Costa Rica), gaviota común (Cuba y la República Dominicana), gaviota golondrina común (Honduras), charrán pico grueso, golondrina-marina común (México), gaviotín (Paraguay), golondrina de mar (Uruguay) y tirra medio cuchillo (Venezuela).[4]
- Un cañón o cálamo es la parte inferior hueca de la pluma de un ave que está insertada en la piel.
- Título usado por Borges y María Kodama es su Breve antología anglosajona.[25]
- Consiste en descender poco a poco y planear de nuevo antes de la zambullida final.
- Los peces depredadores son aquellos que se aprovechan de otros peces o animales; incluyen a la perca, el muskallonga, los lucios, la lucioperca y los salmones.
- En biología y medicina, la feminización es el desarrollo de características físicas que suelen ser exclusivas de la hembra de una especie. Esto puede ser parte del proceso normal de desarrollo, lo que contribuye a la diferenciación sexual. También puede ser inducida por factores ambientales y este fenómeno se ha observado en varias especies animales.[133][134]
Referencias
- BirdLife International (2016). «Sterna hirundo». Lista Roja de especies amenazadas de la UICN 2016.3 (en inglés). ISSN 2307-8235. Consultado el 27 de diciembre de 2016.
- Murray, James A. (2005) [1890]. «Laridæ: Sterna». The avifauna of the island of Ceylon: a systematic account with descriptions of all the known species of birds inhabiting the island, also observations on their habits, nidification, &c, and tables of their geographical distribution throughout India (en inglés). Nueva Delhi: Asian Educational Services. p. 328. ISBN 978-8-120-61974-6. OCLC 70133217.
- Bernis, F; De Juana, E; Del Hoyo, J; Fernández-Cruz, M; Ferrer, X; Sáez-Royuela, R; Sargatal, J (1996). «Nombres en castellano de las aves del mundo recomendados por la Sociedad Española de Ornitología (Tercera parte: Opisthocomiformes, Gruiformes y Charadriiformes)». Ardeola. Handbook of the Birds of the World (Madrid: SEO/BirdLife) 43 (2): 231-238. ISSN 0570-7358. Consultado el 14 de octubre de 2015.
- Charrán Común Sterna hirundo (Linnaeus, 1758) en Avibase.
- Gill, F; Donsker, D, eds. (2017). «Coursers, noddies, gulls, terns, auks & sandgrouse Archivado el 6 de mayo de 2014 en Wayback Machine.» IOC World Bird List (v.7.2) por el Congreso Ornitológico Internacional (en inglés). Consultado el 21 de abril de 2017.
- Olsen y Larsson, 1995, pp. 77-89.
- Firouz, Eskandar (2005). The Complete Fauna of Iran (en inglés). Nueva York: I. B. Tauris. p. 149. ISBN 978-1-85043-946-2. OCLC 57484234.
- Hume, 1993, pp. 21-29.
- Vinicombe, Keith; Tucker, Laurel; Harris, Alan (1990). The Macmillan Field Guide to Bird Identification (en inglés). Londres: Macmillan. pp. 133-138. ISBN 0-333-42773-4. OCLC 717204727.
- Harrison, Peter (1988). Seabirds (en inglés). Londres: Christopher Helm. pp. 370-374. ISBN 0-7470-1410-8. OCLC 19456667.
- Nisbet, I. C. T. «Common Tern (Sterna hirundo)». En Poole, Alan, ed. The Birds of North America Online. Cornell Lab of Ornithology 2002 (en inglés). Ithaca: Universidad Cornell. Consultado el 25 de enero de 2012. (requiere suscripción).
- Campbell, Bruce; Lack, Elizabeth (1985). A dictionary of birds (en inglés). Vermillion: British Ornithologists' Union/Buteo Books. p. 226. ISBN 978-0-931-13012-0. OCLC 12073954.
- Alerstam, T. (1985). «Strategies of migratory flight, illustrated by Arctic and common terns, Sterna paradisaea and Sterna hirundo». En Rankin, M. A., ed. Migration: mechanisms and adaptive significance. Contributions to Marine Science (en inglés) 27 (supl.). Port Aransas: Marine Science Institute, the University of Texas at Austin. pp. 580-603. ISSN 0082-3449. OCLC 14238066.
- Bridge, Eli S.; Eaton, Muir D. (enero de 2005). «Does ultraviolet reflectance accentuate a sexually selected signal in terns?». Journal of Avian Biology (en inglés) (Oxford: Nordic Society Oikos/Blackwell Publishers) 36 (1): 18-21. JSTOR 3677536. doi:10.1111/j.0908-8857.2005.03470.x.
- Bridge, Eli S.; Nisbet, Ian C. T. (mayo de 2004). «Wing molt and assortative mating in Common Terns: a test of the molt-signaling hypothesis». The Condor (en inglés) (Washington D. C.: Cooper Ornithological Society) 106 (2): 336-343. ISSN 0010-5422. JSTOR 1370640. OCLC 4630492786. doi:10.1650/7381.
- Braasch, Alexander; García, Germán O. (marzo de 2012). «A case of aberrant post-breeding moult coinciding with nest desertion in a female Common Tern». British Birds (en inglés) (Londres: MacMillan Jounals) 105: 154-159. ISSN 0007-0335. OCLC 783242642.
- Hume, 1993, pp. 68-75.
- Burger, Joanna; Gochfeld, Michael; Boarman, William I. (enero de 1988). «Experimental evidence for sibling recognition in Common Terns (Sterna hirundo)». The Auk (en inglés) (Washington D. C.: American Ornithologists' Union) 105 (1): 142-148. ISSN 0004-8038. JSTOR 4087337. OCLC 5550616871.
- Stevenson, J. G.; Hutchison, R. E.; Hutchison, J. B.; Bertram, B. C. R.; Thorpe, W. H. (mayo de 1970). «Individual recognition by auditory cues in the Common Tern (Sterna hirundo)». Nature (en inglés) (Londres: Macmillan Journals, Ltd.) 226 (5245): 562-563. ISSN 0028-0836. OCLC 4652363177. PMID 16057385. doi:10.1038/226562a0.
- Snow y Perrins, 1998, p. 764.
- Wassink, Jan L.; Ort, Kathleen (1995). Birds of the Pacific Northwest Mountains: The Cascade Range, the Olympic Mountains, Vancouver Island, and the Coast Mountains (en inglés). Missoula: Mountain Press. p. 78. ISBN 0-87842-308-7. OCLC 31243049.
- Bridge, Eli S.; Jones, Andrew W.; Baker, Allan J. (2005). «A phylogenetic framework for the terns (Sternini) inferred from mtDNA sequences: implications for taxonomy and plumage evolution». Molecular Phylogenetics and Evolution (en inglés) (Orlando: Academic Press) 35 (2): 459-469. ISSN 1055-7903. OCLC 4663268721. PMID 15804415. doi:10.1016/j.ympev.2004.12.010. Archivado desde el original el 19 de abril de 2011.
- Linnæus, Carolus (1758). «Aves». Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis (en latín) I (Décima edición). Estocolmo: Holmiae (Laurentii Salvii). p. 137. ISBN 978-0-66-553008-1. OCLC 49076342. doi:10.5962/bhl.title.542. LCC 03022973.
S. cauda forficata: rectricibus duabus extimis albo nigroque dimitiatis.
- «Sterna». Oxford English Dictionary (en inglés) (Tercera edición). Oxford: Oxford University Press. septiembre de 2005. (requiere suscripción).
- Toswell, M. J. (2014). «Borges the Scholar and Writer». Borges the unacknowledged medievalist: Old English and Old Norse in his life and work. New Middle Ages (en inglés). Nueva York: Palgrave Macmillan. p. 54. ISBN 978-1-137-45129-3. OCLC 890435535. doi:10.1057/9781137444479_4.
- Hume, 1993, pp. 12-13.
- Baerg, William J. (1951). Birds of Arkansas (en inglés) (258). Fayetteville: Agricultural Experiment Station, University of Arkansas. p. 72. OCLC 942279.
- Marcuzzi, G. (1978). «Ecological approach to the study of bird names on the Faeroe Islands». En Bardii, G., ed. Rivista di biologia (en inglés) (Perugia: Università di Perugia) 71 (1-4): 157-272. ISSN 0035-6050. OCLC 114374801. PMID 757887.
- «tarrock». Dictionary of the Scots Language/Dictionar o the Scots Leid (en inglés). Dundee: University of Dundee. Consultado el 16 de octubre de 2015.
- Cocker, Mark; Mabey, Richard (2005). Birds Britannica (en inglés). Londres: Chatto & Windus. pp. 246-247. ISBN 0-7011-6907-9. OCLC 60512849.
- Hume, 1993, pp. 88-89.
- «Sterna hirundo Linnaeus (1758)». Integrated Taxonomic Information System (en inglés). Washington D. C.: ITIS. Consultado el 23 de enero de 2012.
- Gill, F; Donsker, D, eds. (2015). «Coursers, noddies, gulls, terns, auks & sandgrouse Archivado el 6 de mayo de 2014 en Wayback Machine.» IOC World Bird List (v.5.3) por el Congreso Ornitológico Internacional (en inglés). doi 10.14344/IOC.ML.5.0 Consultado el 16 de octubre de 2015.
- Clements, J. F.; Schulenberg, T. S.; Iliff, M. J.; Roberson, D.; Fredericks, T. A.; Sullivan, B. L.; Wood, C. L. (2015). «The eBird/Clements checklist of birds of the world: v2015». The Cornell Lab of Ornithology. Ithaca: Universidad Cornell. Consultado el 16 de octubre de 2015.
- Sushkin, P. P. (1925). «Зоологические области Средней Сибири и ближайших частей нагорной Азии и опыт истории современной фауны палеарктической Азии» [Observaciones de la región zoológica de Siberia central y las zonas montañosas asiáticas circundantes, y antecedentes históricos de la fauna paleártica moderna de Asia]. Boletín de la Sociedad de Naturalistas de Moscú, División Biología (en ruso) 34: 7-86. ISSN 0027-1403.
- Brazil, Mark (2008). Birds of East Asia (en inglés). Londres: A & C Black. p. 220. ISBN 0-7136-7040-1. OCLC 768731565.
- Stuart Baker, E. C. (1929). «Family Sternidæ». Birds. The fauna of British India, including Ceylon and Burma (en inglés) VI (Segunda edición). Londres: Taylor and Francis. pp. 130-132. OCLC 3365406. doi:10.5962/bhl.title.14195. LCC 23002228.
- Firouz, Eskandar (2005). The Complete Fauna of Iran (en inglés). Londres: I. B. Tauris. p. 149. ISBN 978-1-85043-946-2. OCLC 57484234.
- Shirihai, Hadoram (2002). A complete guide to Antarctic wildlife: the birds and marine mammals of the Antarctic continent and the Southern Ocean (en inglés). Degerby: Alula Press. p. 231. ISBN 951-98947-0-5. OCLC 51081322.
- Hume, Rob A. (1993). «Common, Arctic and Roseate Terns: an identification review». British Birds (en inglés) (Londres: MacMillan Jounals) 86: 210-217. ISSN 0007-0335.
- van Duivendijk, Nils (2011). Advanced Bird ID Handbook: The Western Palearctic (en inglés). Londres: New Holland Publishers. pp. 200-202. ISBN 1-78009-022-6. OCLC 752789141.
- Olsen y Larsson, 1995, pp. 69-76.
- Blomdahl, Anders; Breife, Bertil; Holmstrom, Niklas (2007). Flight Identification of European Seabirds (en inglés). Londres: Christopher Helm. p. 340. ISBN 0-7136-8616-2. OCLC 123374738.
- Olsen y Larsson, 1995, pp. 103-110.
- Enticott y Tipling, 2002, p. 196.
- Sinclair, Ian; Hockey, Phil; Tarboton, Warwick (2002). SASOL: Vöels van Suider-Afrika [SASOL: Birds of Southern Africa] (en afrikáans) (Tercera edición). Ciudad del Cabo: Struik. p. 212. ISBN 1-86872-721-1. OCLC 52377304.
- Schulenberg, Thomas S.; Stotz, Douglas F.; Lane, Daniel F.; O'Neill, John P.; Parker, Theodore A. (2010). «Species accounts: Medium-sized terns». Birds of Peru (en inglés). Princeton: Princeton University Press. p. 154. ISBN 0-691-13023-X. OCLC 705944514.
- Enticott y Tipling, 2002, p. 192.
- Simpson, Ken; Day, Nicolas (2010). Field Guide to the Birds of Australia (en inglés) (Octava edición). Camberwell: Penguin Books. p. 110. ISBN 0-670-07231-1. OCLC 471354730.
- Grimmett, Richard; Inskipp, Carol; Inskipp, Tim (2002). Pocket Guide to Birds of the Indian Subcontinent (en inglés). Londres: Christopher Helm. pp. 140-141. ISBN 0-7136-6304-9. OCLC 48883104.
- Cuthbert, Wires y Timmerman, 2003, p. 4.
- Raffaele, Herbert A.; Raffaele, Janis I.; Wiley, James; Garrido, Orlando H.; Keith, Allan R. (2003). Field Guide to the Birds of the West Indies (en inglés). Londres: Christopher Helm. p. 292. ISBN 0-7136-5419-8. OCLC 439837918.
- Hilty, Steven L. (2002). Birds of Venezuela (en inglés). Londres: Christopher Helm. p. 310. ISBN 0-7136-6418-5. OCLC 51031554.
- Lima, Pedro (2006). Aves do litoral norte da Bahia (en portugués e inglés). Bahía: Atualidades Ornitológicas. p. 132. Archivado desde el original el 23 de septiembre de 2015. Consultado el 18 de octubre de 2015.
- Neves, Verónica C.; Bremer, R. Esteban; Hays, Helen W. (2002). «Recovery in Punta Rasa, Argentina of Common Terns banded in the Azores archipelago, North Atlantic». Waterbirds (en inglés) (Washington D. C.: Waterbird Society) 25 (4): 459-461. ISSN 1524-4695. OCLC 4632001918. doi:10.1675/1524-4695(2002)025[0459:RIPRAO]2.0.CO;2.
- Snow y Perrins, 1998, pp. 779-782.
- Hume, 1993, pp. 39-41.
- Hoffmann, Thilo W. (1990). «Breeding of the Common Tern Sterna hirundo in Sri Lanka». Journal of the Bombay Natural History Society (en inglés) (Bombay: Bombay Natural History Society) 87 (1): 68-72. ISSN 0006-6982.
- Hoffmann, Thilo W. (1992). «Confirmation of the breeding of the Common Tern Sterna hirundo Linn. in Sri Lanka». Journal of the Bombay Natural History Society (en inglés) (Bombay: Bombay Natural History Society) 89 (2): 251-252. ISSN 0006-6982.
- Rasmussen, Pamela C.; Anderton, John C. (2005). Birds of South Asia: The Ripley Guide (en inglés) II. Barcelona: Lynx Edicions y el Instituto Smithsoniano. pp. 194-195. ISBN 84-87334-67-9. OCLC 60359701.
- Khan, Asif N. (abril de 2015). «Record of Common Tern Sterna hirundo from Andaman & Nicobar Islands, India». Journal of Bombay Natural History Society (en inglés) (Bombay: Bombay Natural History Society) 112 (1): 30-30. ISSN 0006-6982. OCLC 6755074562. doi:10.17087/jbnhs/2015/v112i1/92329. Archivado desde el original el 2 de junio de 2018. Consultado el 21 de abril de 2017.
- Zimmerman, Dale A.; Pearson, David J.; Turner, Donald A. (2010). Birds of Kenya and Northern Tanzania (en inglés). Londres: Christopher Helm. p. 354. ISBN 0-7136-7550-0. OCLC 62890751.
- Bauer, Hans-Günther; Bezzel, Einhard; Fiedler, Wolfgang, eds. (2005). Nonpasseriformes – Nichtsperlingsvögel. Das Kompendium der Vögel Mitteleuropas: Alles über Biologie, Gefährdung und Schutz (en alemán) I. Wiesbaden: Aula-Verlag Wiebelsheim. p. 647. ISBN 3-89104-647-2. OCLC 611586310.
- Robertson, Hugh; Heather, Barrie (2005). The Field Guide to the Birds of New Zealand (en inglés). Auckland: Penguin Group. p. 126. ISBN 0-14-302040-4. OCLC 63259713.
- Watling, Dick (2003). A Guide to the Birds of Fiji and Western Polynesia (en inglés). Suva: Environmental Consultants. pp. 204-205. ISBN 982-9030-04-0. OCLC 60814638.
- Newton, Ian (2010). Bird Migration. Collins New Naturalist Library (en inglés) 113. Londre: Collins. pp. 150-151. ISBN 0-00-730731-4. OCLC 458729497.
- «Sterna hirundo, additional information». BirdLife International Species factsheet (en inglés). Cambridge: BirdLife International. Archivado desde el original el 4 de septiembre de 2011. Consultado el 26 de enero de 2012.
- DiCostanzo, Joseph (julio de 1978). «Occurrences of the Common Tern in the interior of South America». Bird-Banding (en inglés) (Nueva York: Association of Field Ornithologists/Northeastern Bird-Banding Association) 49 (3): 248-251. ISSN 0006-3630. OCLC 5550373240. doi:10.2307/4512366.
- Darby, Chris (2011). «Eastern Common Terns in Suffolk and Belgium». Birding World (en inglés) (Cley next the Sea: Bird Information Service) 24 (12): 511-512. ISSN 0969-6024.
- Hume, 1993, pp. 30-37.
- Fisher, James; Lockley, R. M. (1989). Sea-birds: an introduction to the natural history of the sea-birds of the North Atlantic. Collins New Naturalist series (en inglés). Londres: Bloomsbury Books. pp. 252-260. ISBN 1-870630-88-2. OCLC 53094179.
- Sandilands, Allan P. (2005). Nonpasserines: waterfowl through cranes. Birds of Ontario: habitat requirements, limiting factors, and status (en inglés) I. Vancouver: University of British Columbia Press. pp. 157-160. ISBN 0-7748-1066-1. OCLC 648347618.
- de Wolf, P. (1984). «BioIndicators and the Quality of the Wadden Sea». En Best, E. P. H.; Haeck, J., eds. Ecological Indicators for the Assessment of the Quality of Air, Water, Soil and Ecosystems: Symposium Papers at a Symposium held in Utrecht, October 1982. Environmental Monitoring & Assessment (en inglés). Dordrecht: D Reidel. p. 362. ISBN 90-277-1708-7. OCLC 840306072.
- Robinson, James A.; Chivers, Lorraine S.; Hamer, Keith C. (2001). «A comparison of Arctic Tern Sterna paradisaea and Common Tern S. hirundo nest-site characteristics on Coquet Island, north-east England». Atlantic Seabirds (en inglés) (Oudeschild: Nederlandse Zeevogelgroep) 3 (2): 49-58. ISSN 1388-2511. Archivado desde el original el 4 de marzo de 2016.
- Ramos, Jaime A.; Adrian, J. (julio de 1995). «Nest-site selection by Roseate Terns and Common Terns in the Azores». The Auk (en inglés) (Washington D. C.: American Ornithologists' Union) 112 (3): 580-589. ISSN 0004-8038. JSTOR 4088675. OCLC 5550933835.
- Fuchs, Eduard (junio de 1977). «Predation and anti-predator behaviour in a mixed colony of terns Sterna sp. and Black-Headed Gulls Larus ridibundus with special reference to the Sandwich Tern Sterna sandvicensis». Ornis Scandinavica (en inglés) (Oslo: Scandinavian Ornithologists' Union/Universitetsforlaget) 8 (1): 17-32. ISSN 0030-5693. OCLC 5553053683. doi:10.2307/3675984.
- Erwin, Michael R. (octubre de 1977). «Black Skimmer breeding ecology and behavior». The Auk (en inglés) (Washington D. C.: American Ornithologists' Union) 94 (4): 709-717. ISSN 0004-8038. OCLC 5554061038. doi:10.2307/4085267.
- Hume, 1993, pp. 86-90.
- Safina, Carl; Burger, Joanna (octubre de 1988). «Prey dynamics and the breeding phenology of Common Terns (Sterna hirundo)». The Auk (en inglés) (Washington D. C.: American Ornithologists' Union) 105 (4): 720-726. ISSN 0004-8038. JSTOR 4087385. OCLC 5553975293.
- González-Solís, J.; Becker, P. H.; Wendeln, H. (noviembre de 1999). «Divorce and asynchronous arrival in Common Terns (Sterna hirundo)». Animal Behaviour (en inglés) (Londres: Baillière Tindall/Association for the Study of Animal Behaviour) 58 (5): 1123-1129. ISSN 0003-3472. OCLC 4922183526. PMID 10564616. doi:10.1006/anbe.1999.1235.
- Hume, 1993, pp. 79-85.
- Hume, 1993, pp. 91-99.
- Hume, 1993, pp. 100-111.
- Palestis, Brian G. (2009). «Use of artificial eelgrass mats by saltmarsh-nesting Common Terns (Sterna hirundo)». In Vivo (en inglés) 30 (3): 11-16.
- Lloyd, Clare; Tasker, Mark L.; Partridge, Ken (2010). The Status of Seabirds in Britain and Ireland (en inglés). Londres: Poyser. p. 207. ISBN 1-4081-3800-X. OCLC 747400583.
- Bent, Arthur Cleveland (1921). Arctic tern. «Life histories of North American gulls and terns: order Longipennes». Bulletin of the United States National Museum (en inglés) (113) (Washington D. C.: Smithsonian Institute, United States National Museum/U. S. Government Printing Office). p. 252. OCLC 988652143.
- «Common Tern Sterna hirundo (Linnaeus, 1758)». Bird Facts (en inglés). Thetford: British Trust for Ornithology. Consultado el 9 de febrero de 2012.
- Hume, 1993, pp. 112-119.
- Hume y Pearson, 1993, pp. 121-124.
- Hume, 1993, pp. 120-123.
- Burger, Joanna; Shealer, D. A.; Gochfeld, Michael (1993). «Defensive aggression in terns: discrimination and response to individual researchers». Aggressive Behavior (en inglés) (Nueva York: Wiley-Liss, Inc.) 19 (4): 303-311. ISSN 0096-140X. OCLC 4654346541. doi:10.1002/1098-2337(1993)19:4<303::AID-AB2480190406>3.0.CO;2-P.
- Hunter, Rodger A.; Morris, Ralph D. (julio de 1976). «Nocturnal predation by a Black-Crowned Night Heron at a Common Tern colony». The Auk (en inglés) (Washington D. C.: American Ornithologists' Union) 93 (3): 629-633. ISSN 0004-8038. JSTOR 4084965. OCLC 5550623821.
- Nisbet, Ian C. T.; Welton, M. (febrero de 1984). «Seasonal variations in breeding success of Common Terns: consequences of predation». The Condor (en inglés) (Washington D. C.: Cooper Ornithological Society) 86 (1): 53-60. ISSN 0010-5422. JSTOR 1367345. OCLC 5550257668. doi:10.2307/1367345.
- Hays, H. (abril de 1984). «Common Terns raise young from successive broods». The Auk (en inglés) (Washington D. C.: American Ornithologists' Union) 101 (2): 274-280. ISSN 0004-8038. JSTOR 4086364. OCLC 5550623603.
- Nisbet, Ian C. T.; Cam, Emmanuelle (enero de 2002). «Test for age-specificity in survival of the Common Tern». Journal of Applied Statistics (en inglés) (Sheffield: Sheffield City Polytechnic/Routledge) 29 (1-4): 65-83. ISSN 0266-4763. OCLC 4636828235. doi:10.1080/02664760120108467.
- Austin, Oliver L., Sr. (1953). «A Common Tern at least 23 years old». Bird-Banding (en inglés) (Nueva York: Association of Field Ornithologists/Northeastern Bird-Banding Association) 24 (1): 20. ISSN 0006-3630.
- «Longevity records for Britain & Ireland in 2010». Online ringing report (en inglés). Thetford: British Trust for Ornithology. Archivado desde el original el 28 de febrero de 2012. Consultado el 11 de febrero de 2012.
- «European Longevity Records». Longevity (en inglés). Wilhelmshaven: European Union for Bird Ringing. Consultado el 11 de febrero de 2012.
- Hume, 1993, pp. 55-67.
- Beaman, Mark; Madge, Steve; Burn, Hilary; Zetterstrom, Dan (1998). The Handbook of Bird Identification: for Europe and the western Palearctic (en inglés). Londres: Christopher Helm. p. 440. ISBN 0-7136-3960-1. OCLC 754713791.
- Kirkham, Ian R.; Nisbet, Ian C. T. (1987). «Feeding techniques and field identification of Arctic, Common and Roseate Terns». British Birds (en inglés) (Londres: MacMillan Jounals) 80 (2): 41-47. ISSN 0007-0335.
- «Sterna hirundo». BirdLife International Species factsheet (en inglés). Cambridge: BirdLife International. Consultado el 26 de enero de 2012.
- Bugoni, Leandro; Vooren, Carolus Maria (julio de 2004). «Feeding ecology of the Common Tern Sterna hirundo in a wintering area in southern Brazil». The Ibis (en inglés) (Tring: British Ornithologists' Union) 146 (3): 438-453. ISSN 0019-1019. OCLC 5157025128. doi:10.1111/j.1474-919X.2004.00277.x.
- Sinclair, Sandra (1985). How Animals See: Other Visions of Our World (en inglés). Beckenham: Croom Helm. pp. 93-95. ISBN 0-7099-3336-3. OCLC 12501466.
- Varela, F. J.; Palacios, A. G.; Goldsmith, T. M. (1993). «Color vision of birds». En Zeigler, Harris Philip; Bischof, Hans-Joachim, eds. Vision, brain, and behavior in birds: a comparative review (en inglés). Cambridge: MIT Press. pp. 77-94. ISBN 0-262-24036-X. OCLC 27727176.
- Lythgoe, J. N. (1979). The Ecology of Vision (en inglés). Oxford: Clarendon Press. pp. 180-183. ISBN 0-19-854529-0. OCLC 4804801.
- Håstad, Olle; Ernstdotter, Emma; Ödeen, Anders (septiembre de 2005). «Ultraviolet vision and foraging in dip and plunge diving birds». Biology Letters (en inglés) (Londres: The Royal Society) 1 (3): 306-309. ISSN 1744-9561. OCLC 4662132955. PMC 1617148. PMID 17148194. doi:10.1098/rsbl.2005.0320.
- Stephens, David W.; Brown, Joel Steven; Ydenberg, Ronald C. (2007). Foraging: behavior and ecology (en inglés). Chicago: University of Chicago Press. p. 295. ISBN 0-226-77264-0. OCLC 310969610.
- Hopkins, C. D.; Wiley, R. H. (julio de 1972). «Food parasitism and competition in two terns». The Auk (en inglés) (Washington D. C.: American Ornithologists' Union) 89 (3): 583-594. ISSN 0004-8038. JSTOR 4084258. OCLC 5550600912.
- Bélisle, M. (septiembre de 1998). «Foraging group size: models and a test with jaegers kleptoparasitizing terns». Ecology (en inglés) (Washington D. C.: Ecological Society of America) 79: 1922-1938. ISSN 0012-9658. OCLC 5165737562. doi:10.2307/176699.
- Hatch, J. J. (julio de 1975). «Piracy by laughing gulls Larus atricilla: an example of the selfish group». The Ibis (en inglés) (Tring: British Ornithologists' Union) 117 (3): 357-365. ISSN 0019-1019. OCLC 5156974874. doi:10.1111/j.1474-919X.1975.tb04222.x.
- Dunn, E. K. (julio de 1973). «Robbing behavior of Roseate Terns». The Auk (en inglés) (Washington D. C.: American Ornithologists' Union) 90 (3): 641-651. ISSN 0004-8038. JSTOR 4084163. OCLC 5550931455. doi:10.2307/4084163.
- Kaufman, K. (1996). Lives of North American birds (en inglés). Boston: Houghton Mifflin. p. 260. ISBN 0-395-77017-3. OCLC 59591478.
- Nisbet, Ian C. T.; Wilson, Karen J.; Broad, William A. (abril de 1978). «Common Terns raise young after death of their mates». The Condor (en inglés) (Washington D. C.: Cooper Ornithological Society) 80 (1): 106-109. ISSN 0010-5422. OCLC 5550390087. doi:10.2307/1367802.
- Hughes, M. R. (abril de 1968). «Renal and extrarenal sodium excretion in the Common Tern Sterna hirundo». Physiological Zoology (en inglés) (Chicago: University of Chicago Press) 41: 210-219. ISSN 0031-935X. JSTOR 30155452. OCLC 5554660478.
- Karleskint, George; Turner, Richard; Small, James (2009). Introduction to Marine Biology (en inglés). Florence: Brooks/Cole. p. 317. ISBN 0-495-56197-5. OCLC 316433444.
- Austin, O. L. (abril de 1948). «Predation by the common rat (Rattus norvegicus) in the Cape Cod colonies of nesting terns». Bird-Banding (en inglés) (Nueva York: Association of Field Ornithologists/Northeastern Bird-Banding Association) 19 (2): 60-65. ISSN 0006-3630. OCLC 5553098558. doi:10.2307/4510014.
- «Common Tern Sterna hirundo». Latest population trends (en inglés). Londres: Joint Nature Conservation Committee. Consultado el 25 de enero de 2012.
- Burger, J.; Gochfeld, M. (1991). «Predators in common terns colonies». The Common Tern: its breeding biology and social behavior (en inglés). Nueva York: Columbia University Press. pp. 169-177. ISBN 978-1-583-48110-3. OCLC 156744971.
- del Hoyo, Josep; Elliott, Andrew; Sargatal, Jordi (1996). Hoatzin to auks. Handbook of the Birds of the World (en inglés) III. Barcelona: Lynx Edicions. p. 653. ISBN 84-87334-20-2. OCLC 230184105.
- Parkes, K. C.; Poole, A.; Lapham, H. (1971). «The Ruddy Turnstone as an egg predator». Wilson Bulletin (en inglés) (Lawrence: Wilson Ornithological Society) 83 (3): 306-307. ISSN 0043-5643. JSTOR 4160107. OCLC 5550935045.
- Farraway, A.; Thomas, K.; Blokpoel, H. (noviembre de 1986). «Common Tern egg predation by Ruddy Turnstones». The Condor (en inglés) (Washington D. C.: Cooper Ornithological Society) 88 (4): 521-522. ISSN 0010-5422. OCLC 5550257305. doi:10.2307/1368282.
- Houde, P. (1977). «Gull-tern interactions on Hicks Island». Proceedings of the Linnean Society of New York (en inglés) (Nueva York: Linnean Society of New York) 73: 58-64. ISSN 0075-9694.
- Whittam, R. M.; Leonard, M. L. (mayo de 2000). «Characteristics of predators and offspring influence on nest defense by Arctic and Common Terns». The Condor (en inglés) (Washington D. C.: Cooper Ornithological Society) 102 (2): 301-306. ISSN 0010-5422. JSTOR 1369641. OCLC 5550395593. doi:10.1650/0010-5422(2000)102[0301:COPAOI]2.0.CO;2.
- Morris, R. D.; Wiggins, D. A. (marzo de 1986). «Ruddy Turnstones, Great Horned Owls, and egg loss from Common Tern clutches». Wilson Bulletin (en inglés) (Lawrence: Wilson Ornithological Society) 98: 101-109. ISSN 0043-5643. JSTOR 4162188. OCLC 5550644639.
- Rothschild y Clay, 1953, p. 135.
- Rothschild y Clay, 1953, pp. 194-197.
- Fiorello, Christine V.; Nisbet, Ian C. T.; Hatch, Jeremy J.; Corsiglia, Carolyn; Pokras, Mark A. (2009). «Hematology and absence of hemoparasites in breeding Common Terns (Sterna hirundo) from Cape Cod, Massachusetts». Journal of Zoo and Wildlife -Medicine (en inglés) (Lawrence: American Association of Zoo Veterinarians) 40 (3): 409-413. ISSN 1042-7260. OCLC 4631528914. PMID 19746853. doi:10.1638/2006-0067.1.
- Olsen, Björn; Munster, Vincent J.; Wallensten, Anders; Waldenström, Jonas; Osterhaus, D. M. E.; Fouchier, Ron A. M. (abril de 2006). «Global patterns of influenza A virus in wild birds». Science (en inglés) (Washington D. C.: American Association for the Advancement of Science) 312 (5772): 384-388. ISSN 0036-8075. OCLC 109143568. PMID 16627734. doi:10.1126/science.1122438.
- Enticott y Tipling, 2002, p. 194.
- Kress, Stephen W.; Weinstein, Evelyn H.; Nisbet, Ian C. T.; Shugart, Gary W.; Scharf, William C.; Blokpoel, Hans; Smith, Gerald A.; Karwowski, Kenneth; Maxwell, George R.; Chapdelaine, Gilles; Montevecchi, William A.; Lock, Anthony R.; Smith, Carol F.; Miller, Eileen; Spendelow, Jeffrey A.; Gochfeld, Michael; Burger, Joanna; Erwin, R. Michael (enero de 1983). «The status of tern populations in northeastern United States and adjacent Canada». Colonial Waterbirds (en inglés) (Washington D. C.: Waterbird Society) 6: 84-106. ISSN 0738-6028. OCLC 5552956379. doi:10.2307/1520976.
- Cuthbert, 2003, p. 1.
- Fry, D. M.; Toone, C. K. (agosto de 1981). «DDT-induced feminization of gull embryos». Science (en inglés) (Washington D. C.: American Association for the Advancement of Science) 213 (4510): 922-924. ISSN 0036-8075. OCLC 113028725. PMID 7256288. doi:10.1126/science.7256288.
- Gimeno, S.; Gerritsen, A.; Bowmer, T.; Komen, H. (noviembre de 1996). «Feminization of male carp». Nature (en inglés) (Londres: Macmillan Journals, Ltd.) 384 (6606): 221-222. ISSN 0028-0836. OCLC 4653850419. PMID 8918871. doi:10.1038/384221a0.
- Hart, Constance A.; Nisbet, Ian C. T.; Kennedy, Sean W.; Hahn, Mark E. (2003). «Gonadal feminization and halogenated environmental contaminants in Common Terns (Sterna hirundo): evidence that ovotestes in male embryos do not persist to the prefledgling stage». Ecotoxicology (en inglés) (Londres: Chapman & Hall) 12 (1-4): 125-140. ISSN 0963-9292. OCLC 5649151959. PMID 12739862. doi:10.1023/A:1022505424074.
- Huntley, Brian; Green, Rhys E.; Collingham, Yvonne C.; Willis, Stephen G. (2007). A climatic atlas of European breeding birds (en inglés). Barcelona: Durham University, The RSPB and Lynx Editions. p. 227. ISBN 978-84-96553-14-9. OCLC 254207990.
- «Annex 2: Waterbird species to which the Agreement applies». African-Eurasian Waterbird Agreement (en inglés). Bonn: UNEP/AEWA Secretariat. Archivado desde el original el 26 de agosto de 2012. Consultado el 25 de enero de 2012.
- «List of Migratory Birds». Birds protected by the Migratory Bird Treaty Act (en inglés). Washington D. C.: U. S. Fish and Wildlife Service. Consultado el 11 de junio de 2017.
- «Introduction». African-Eurasian Waterbird Agreement (en inglés). Bonn: UNEP/AEWA Secretariat. Archivado desde el original el 26 de agosto de 2012. Consultado el 25 de enero de 2012.
- «Migratory Bird Treaty Act of 1918». Digest of Federal Resource Laws of Interest to the U. S. Fish and Wildlife Service (en inglés). Washington D. C.: U. S. Fish and Wildlife Service. Consultado el 25 de enero de 2012.
Bibliografía consultada
- Cuthbert, Francesca J.; Wires, Linda R.; Timmerman, Kristina (2003). Status Assessment and Conservation Recommendations for the Common Tern (Sterna hirundo) in the Great Lakes Region (en inglés). Fort Snelling: U. S. Department of the Interior, Fish and Wildlife Service. Archivado desde el original el 27 de noviembre de 2014.
- Enticott, Jim; Tipling, David (2002). Seabirds of the World (en inglés). Londres: New Holland Publishers. ISBN 1-84330-327-2. OCLC 54397121.
- Hume, Rob (1993). The Common Tern (en inglés). Londres: Hamlyn. ISBN 0-540-01266-1. OCLC 28963398.
- Olsen, Klaus Malling; Larsson, Hans (1995). Terns of Europe and North America (en inglés). Londres: Christopher Helm. ISBN 0-7136-4056-1. OCLC 31970361.
- Rothschild, Miriam; Clay, Theresa (1953). Fleas, Flukes and Cuckoos. A Study of Bird Parasites (en inglés). Londres: Collins. OCLC 2889018. doi:10.5962/bhl.title.6413.
- Snow, David; Perrins, Christopher M. (1998). The Birds of the Western Palearctic (BWP), concise edition (2 volumes) (en inglés). Oxford: Oxford University Press. ISBN 0-19-854099-X. OCLC 37180316.
Enlaces externos
 Wikispecies tiene un artículo sobre Sterna hirundo.
Wikispecies tiene un artículo sobre Sterna hirundo. Wikimedia Commons alberga una galería multimedia sobre Sterna hirundo.
Wikimedia Commons alberga una galería multimedia sobre Sterna hirundo.- Gill, F.; Donsker, D., eds. (2015). «Coursers, noddies, gulls, terns, auks & sandgrouse Archivado el 6 de mayo de 2014 en Wayback Machine.» IOC World Bird List (v.5.4) por el Congreso Ornitológico Internacional (en inglés).
- Ficha de la UICN (en inglés).
- «Sterna hirundo (TSN 176888)». Sistema Integrado de Información Taxonómica (en inglés).
- Sterna hirundo (Linnaeus, 1758) Archivado el 13 de marzo de 2016 en Wayback Machine. en Catalogue of Life (en inglés).
- Ficha en Animal Diversity Web (en inglés).
- CITES: taxon Sterna hirundo (Linnaeus, 1758) (en francés).
- Ficha en Fauna Europaea (en inglés).
- Ficha del Centro Nacional para la Información Biotecnológica (en inglés).
- Peterson, Alan P. «Sterna hirundo (Linnaeus, 1758) en el orden Charadriiformes» (en inglés).
- Videos, fotos y grabaciones sonoras en Internet Bird Collection (en inglés).
- «Charrán Común (Sterna hirundo) • (Linnaeus, 1758)». xeno-canto. Leiden: Naturalis Biodiversity Center. Consultado el 23 de octubre de 2015.
- Common Tern - Sterna hirundo VIREO (Universidad Drexel) (en inglés).
- Ficha en el Cornell Lab of Ornithology (en inglés).
- USGS Patuxent Bird Identification InfoCenter: Identificación anatómica (en inglés).
| Control de autoridades |
|
|---|
 Datos: Q18875
Datos: Q18875- Multimedia: Sterna hirundo / Q18875
- Especies: Sterna hirundo