Transportador ABC
Los transportadores dependientes de ATP o transportadores ABC (del inglés ATP-binding cassette) son un grupo de proteínas transportadoras de membrana. Conforman una de las mayores y más antiguas superfamilias de dichas proteínas, con representantes en todos los filos existentes, desde los procariotas a los seres humanos.[1][2] Constan de varias subunidades: una o dos proteínas transmembranales y una o dos ATPasas asociadas a la membrana. Las ATPasas proporcionan la energía para la absorción o secreción de diversos sustratos a través de la membrana mediante la hidrólisis de ATP. La mayoría de los sistemas de absorción cuentan también con un receptor extracitoplasmático, una proteína a la que se liga la molécula transportada. Algunas proteínas ABC no participan en el transporte de moléculas, sino en la traducción y reparación del ADN.[3]
Los transportadores de ABC absorben numerosos nutrientes, precursores biosintéticos, oligoelementos y vitaminas, y secretan lípidos, esteroles y una amplia variedad de metabolitos primarios y secundarios. En los seres humanos, algunos de los sistemas secretores participan en los procesos de resistencia a los tumores, la fibrosis quística y otras enfermedades hereditarias. Una expresión elevada de los genes codificantes de ciertos transportatores secretores conduce al desarrollo de la resistencia a medicamentos y agentes quimioterapéuticos en organismos procariotas y eucariotas.
Se han hallado centenares de genes de transportadores ABC,[4] esenciales para multitud de funciones celulares. En los seres humanos se han encontrado 48 genes y mutaciones en ellos causan o contribuyen a diversas enfermedades,[5] como la fibrosis quística, la adrenoleucodistrofia, la enfermedad de Stargardt, los tumores resistentes a la quimioterapia, el síndrome de Dubin-Johnson, la colestasis intrahepática familiar progresiva, la anemia sideroblástica, la ataxia, y la hipoglucemia hiperinsuliménica.[4]
Función
Los transportadores ABC utilizan la energía proveniente de la hidrólisis del ATP para transportar diversas sustancias a través de la membrana celular. Se dividen en tres categorías funcionales principales. En los organismos procariotas se encuentran transportadores ABC implicados en la absorción de nutrientes. Entre los sustratos incorporados a la célula por estos transportadores se cuentan iones, aminoácidos, péptidos, azúcares y otras moléculas hidrofílicas. La porción transmembranal del transportador ABC aísla los sustratos de los lípidos presentes en la membrana mientras la atraviesan. Los organismos eucariotas, en cambio, no poseen transportadores ABC dedicados a la absorción. Un segundo tipo de transportadores dedicados a la secreción o excreción de sustancias están presentes tanto en los procariotas como los eucariotas. Estos transportadores bombean toxinas fuera de la célula. En las bacterias gramnegativas, los transportadores secretores llevan lípidos y algunos polisacáridos desde el citoplasma al periplasma. Existe un tercer subgrupo de proteínas ABC sin función transportadora, que participan en la traducción y los procesos de reparación del ADN.[3]
Procariotas
Los transportadores ABC bacterianos son esenciales para la supervivencia de la célula, la virulencia y la patogenicidad.[3] Por ejemplo, los sistemas de absorción del hierro son importantes para la virulencia;[6] los patógenos usan sideróforos, como la enterobactina, para capturar hierro ligado a proteínas con alta afinidad por el hierro o eritrocitos. Otro ejemplo es producto del gen chvE-gguAB en Agrobacterium tumefaciens, un transportador de glucosa y galactosa igualmente asociado con la virulencia.[7][8] Los transportadores ABC también preservan la integridad de la célula ante cambios que puedan afectarla. Por ejemplo, la activación de ciertos transportadores sensibles a la presión osmótica neutraliza los cambios potencialmente letales en la concentración de solutos en la célula mediande la absorción de iones.[9] Algunas proteínas de transporte ABC bacterianas también desempeñan un papel en la regulación de varios procesos fisiológicos.[3] Los transportadores secretores bacterianos eliminan componentes de la superficie de la célula bacteriana, como polisacáridos, lipopolisacáridos y ácido teicoico, así como proteínas involucradas en la patogénesis bacteriana, hemos, enzimas hidrolíticas, proteínas de la capa S, factor de competencia, toxinas, antibióticos, medicamentos y sideróforos.[10] También tienen roles en algunos procesos de biosíntesis.[11]
Eucariotas
Aunque la mayoría de transportadores ABC eucariotas son secretores, algunos no transportan el sustrato directamente. Por ejemplo, en el regulador de la conductancia transmembrana de la fibrosis quística y en el receptor de la sulfonilurea (SUR), la hidrólisis del ATP conlleva la regulación de la apertura y cierre de canales iónicos asociados bien al transportador ABC mismo, bien a otras proteínas.[12] Las proteínas ABC se han detectado en la placenta, donde podrían proteger al feto contra sustancias xenobióticas.[13]
Los polimorfismos en los genes de los transportadores ABC humanos contribuyen a varias enfermedades, aun sin causar una pérdida de función completa de las proteínas ABC que codifican.[14] Entre estas enfermedades se cuentan la fibrosis quística, la adrenoleucodistrofia, la enfermedad de Stargardt, la enfermedad de Tangier, algunas inmunodeficiencia, el síndrome de Dubin-Johnson, la colestasis intrahepática familiar progresiva, el pseudoxantoma elástico, la anemia sideroblástica, la ataxia, la hipoglucemia hiperinsuliménica, la degeneración macular asociada a la edad, la hipoproteinemia familiar, la retinosis pigmentaria y la distrofia corneal entre otras.[12]
La familia de proteínas ABCB humanas es responsable de la resistencia antineoplásica. La proteína ABCB1 o glucoproteína-P tiene un papel en diversos procesos biológicos donde el transporte de lípidos es la función principal; es un mediador en la secreción de la aldosterona por las glándulas adrenales y su inhibición bloquea la migración de células dendríticas.[15] ABCB1 también regula el transporte de cortisol y dexametasona, pero no el de progesterona. Puede transportar colesterol, análogos de fosfatidilcolina (PC), fosfatidiletanolamina (PE), fosfatidilserina (PS), esfingomielina (SM), y glucosilceramida (GlcCer).[14]
Estructura
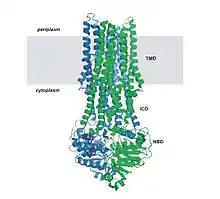
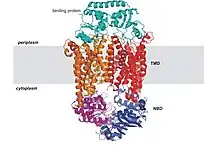
Todas las proteínas de transporte ABC comparten cuatro dominios básicos:[16] dos dominios transmembrana denominados comúnmente (TMD transmembrane domain o IMD integral membrane domain) y dos dominios citosólicos, llamados ABC o NBD —ATP-binding cassette domain o nucleotide-binding domain.
Los dominos transmembrana presentan una gran variedad en su estructura y secuencias, consistente con la diversidad de sustratos que pueden transportar. La secuencia de los dominios ABC, en cambio, está muy bien conservada, con muchas similitudes entre transportadores diferentes. En la mayoría de los ABC secretores o exportadores, los cuatro subdominios forman una sola cadena polipeptídica con una disposición TM-NB-TM-NB. Esta configuración se invierte en los ABC importadores, dispuestos como NB-TM-NB-TM.[3][12] Los cuatro dominios pueden presentarse también como dos o cuatro polipéptidos separados, como ocurre principalmente en las bacterias.[17]
Además de los cuatro dominios básicos, algunos transportadores ABC constan de elementos adicionales que contribuyen a la función regulatoria de este tipo de proteínas. Por ejemplo, los ABC importadores tienen una proteína de unión (binding protein o BP) de alta afinidad que liga al sustrato presente en el periplasma y lo traspasa al transportador. Los exportadores no tienen una BP, pero poseen un dominio intracelular o ICD (intracellular domain) que une las hélices del dominio transmembrana con el dominio ABC y que posiblemente participe en la comunicación entre ambos dominios.[18]
Dominio transmembrana
Los dominios TM constan típicamente de doce hélices alfa —seis hélices por monómero— que abarcan todo el espesor de la membrana plasmática, y que sirven como conducto de la sustancia transportada, pero el número puede también variar. Los dominios transmembranas se clasifican en tres tipos de plegamientos basados en una caracterización detallada de la secuencia de aminoácidos: importador ABC de tipo I, importador ABC de tipo II y secretor ABC o tipo III.[18] El plegamiento de tipo I se observó por vez primera en la subunidad transmembrana de un transportador de molibdato.[19] Este plagamiento se caracteriza por el emplazamiento de las hélices 2-5 a lo largo de la vía de transporte, perpendicularlmente a las superficies interior y exterior de la membrana, y de la hélice 1 que rodea la superficie exterior del haz formado por las otras hélicesy está en contacto con el resto de las hélices. El tipo II se ha encontrado en la proteína BtuCD[20] y en Hi1471, un transportador de Haemophilus influenzae.[21] La hélice 2 atraviesa el centro de la subunidad, rodeada por las otras hélices, y las hélices 5 y 10 se encuentran en la interfaz del dominio transmembrana. El tipo III, descrito inicialmente en el exportador Sav1866 se organiza en dos alas, cada una compuesta de las hélices 1 y 2 de un monómero y las hélices 3-6 del segundo monómero. Las hélices 1-3 y 4-6 están relacionadas por un eje de simetría axial perpendicular a la superficie de la membrana.[18]
Dominio NB
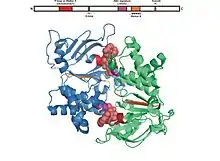
El dominio asociado al nucleótido consiste a su vez de dos subdominios: el núcleo catalítico, similar a las proteínas RecA, y un subdominio de menor tamaño compuesto de hélices alfa que solo aparece en los transportadores ABC y presenta una marcada diversidad estructural. El subdominio mayor consta de dos láminas beta y seis hélices alfa, y cuenta con un bucle P (motivo de Walker A) y un motivo de Walker B —constituido por cuatro aminoácidos hidrofóbicos y un aspartato. El subdominio menor contiene tres o cuatro hélices y el llamado motivo característico ABC, motivo LSGGQ, péptido de enlace o motivo C. El dominio NB también tiene una glutamina en un bucle flexible conocido como bucle Q, tapadera o conmutador de fosfato-gamma, que conecta con el dominio transmembrana; se supone que el bucle Q participa en la interacción entre los dominios NB y TM y tiene que ver con la relación entre la hidrólisis del nucleótido y los cambios en conformación que experimenta el dominio transmembrana durante la traslocación del sustrato. Una última zona, conocida como el motivo H or región de conmutación contiene una histidina muy conservada que desempeña un papel importante en la interacción del dominio NB con el ATP. La denominación del dominio asociado al nucleótido como ABC o ATP-binding cassette proviene de la forma de caja o casete que adoptan los motivos que lo integran cuando se produce la hidrólisis del ATP.[3][10][18]
Unión e hidrólisis del ATP
La dimerización del dominio ABC o NB de los transportadores requiere la asociación con ATP.[22] Cuando el ATP se encuentra unido, las interacciones entre los dos dominios ABC tienen lugar sobre una amplia superficie, pero en la ausencia del nucleótido aumenta la distancia entre los dos dominios.[18] Se han elucidado varias estructuras del dominio ABC de varior importadores, como el HisP,[23] GlcV,[24] MJ1267,[25] E. coli MalK,[26] T. litoralis MalK,[27] y de secretores como TAP,[28] HlyB,[29] MJ0796,[30][31] Sav1866,[32] y MsbA.[33] En todos estos transportadores, dos moléculas de ATP se unen a los dominios NB en la interfaz formada por el motivo Walker A de una subunidad y el motivo LSGGQ de la otra.[18] Esta conformación es consistente con resultados de experimentos bioquímicos que demuestran que el ATP se encuentra próximo a los aminoácidos del bucle P y el motivo LSGGQ durante el proceso de catálisis.[34]
Las siguientes interacciones son importantes para la estabilidad del compuesto proteína+nucleótido: 1) interacción entre un residuo aromático que precede al bucle P y el anillo de la adenosina del ATP;[35][36] 2) enlace de hidrógeno entre una lisina en el bucle P y dos átomos de oxígeno en los fosfatos del ATP y coordinación de estos fosfatos y varios aminoácidos del bucle P con un ion Mg2+, que también está asociado al aspartato del motivo Walker D vía la molécula de agua que reaccionará con el ATP;[24][28] 3) coordinación de un fosfato con la serina y los grupos amida de las glicinas en el motivo LSGGQ.[37] La histidina en el bucle H también presenta contactos con ambas subunidades.[26][31][38] No hay un consenso firme sobre el mecanismo preciso de catálisis del ATP;[3] Tanto el glutamato adyacente al motivo Walker B, la glutamina en el bucle P o la histidina en el motivo H podrían estar implicados.[31]
Mecanismo de transporte
Los transportadores ABC son transportadores activos, es decir, necesitan energía en forma de ATP para trasladar los sustratos a través de la membrana celular. La hidrólisis del ATP en el dominio ABC — dominio de unión al nucleótido— causa un cambio conformacional en los dominios transmembrana, que alternan entre una orientación hacia adentro y otra hacia afuera.[17] Este cambio conformacional empuja al sustrato desde un extremo de la membrana hacia el otro.[39] Tanto los importadores como los secretores utilizan el mismo mecanismo, siendo la afinidad por el sustrato en cada una de las dos conformaciones lo que determina la dirección de transporte: en los importadores, el transportador se une más fácilmente al sutrato en la orientación hacia fuera, al contrario de lo que ocurre con los exportadores.[18]
En todos los transportadores ABC el estado de reposo se caracteriza por encontrarse el dominio ABC en un estado abierto, con baja afinidad hacia el ATP y que deja una oquedad abierta al interior del transportador. Al asociarse el dominio TM al sustrato, se producen cambios en el domino ABC que facilitan la unión a dos moléculas de ATP y la adopción del estado cerrado por el dímero ABC. En esta conformación el dominio ABC fuerza la apertura del dominio TM por el lado opuesto al de la unión del sustrato, que se traslada a lo largo del TM. Al haber en esta posición menor afinidad entre el transportador y el sustrato, este queda libre. Tras la hidrólisis del ATP, el ADP restaura la conformación de reposo.
No hay acuerdo sobre qué parte del ciclo proporciona la energía inicial para la función de los transportadores ABC. Generalmente se supone que la hidrólisis del ATP es el motor principal del proceso,[40] pero los resultados de análisis estructurales y bioquímicos apuntan a la unión con el ATP, o la dimerización del dominio ABC resultante como el paso que recarga la energía del ciclo.[39] Análisis bioquímicos también sugieren que la unión de las dos moléculas de ATP es un proceso cooperativo, es decir, que el ATP debe asociarse a ambos sitios activos en los dominios ABC antes de que estos puedan dimerizarse y adoptar la configuración cerrada y catalíticamente activa.[39]
Importadores ABC
Los importadores ABC transportan nutrientes desde el exterior hasta el interior de la célula. En la mayoría de las bacterias, las molécules se unen primero a una proteína de unión con alta afinidad por el soluto, que puede estar ubicada en el espacio periplasmático entre las membranas exterior e interior de las bacterias gramnegativas, o, en las bacterias grampositivas, unida a la cara externa de la membrana citoplásmica o incluso fusionada con el transportador mismo.[3] La primera estructura conocida de un importador completo fue la del transportador de molibdeno ModBC A de Archaeoglobus fulgidus,[19] a la que siguieron otras. Las estructuras muestran las dos distintas configuraciones del transportador, con aperturas en los dos extremos opuestos. Otra característica común es que el dominio ABC y el dominio transmembrana están unidos por una hélice corta, la «hélice de acoplamiento», que adopta una orientación paralela a la de la bicapa lipídica.[41]
Mecanismo de transporte en los importadores
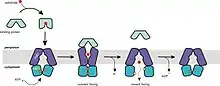
En el estado de reposo —en orientación hacia adentro—, el dominio transmembrana (TM) mantiene abierta la interfaz del dímero del dominio de unión al nucleótido (ABC), ocluyendo el acceso al citoplasma. Al unirse la proteína de unión cargada con el sustrato al dominio TM, el ATP se asocia al dominio ABC y el dímero se cierra. El dominio transmembrana adopta la orientación hacia afuera para recibir el sustrato. Tras la hidrólisis del ATP, el dímero ABC se abre y el sustrato pasa al citoplasma. Al liberarse el ADP y fosfato el transportador adopta de nuevo la configuración inicial de reposo.[3][10][18][39][41]
Exportadores ABC
Los secretores o exportadores ABC procariotas abundan mucho y cuentan con homólogos cercanos en los organismos eucariotas. Los secretores se clasifican según el tipo de sustrato que transporten en dos clases principales. Una clase se especializa en la secreción de proteínas, como toxinas, hidrolasas, proteínas de la capa S, lantibióticos, bacteriocinas y factores de competencia; la otra clase comprende los exportadores de moléculas orgánicas de menor tamaño. Los secretores ABC han despertado un gran interés porque contribuyen a la resistencia de las células a los antibióticos y los medicamentos contra el cáncer, al expulsar estas moléculas del interior de la célula.[3] Entre los exportadores responsables de la resistencia a los medicamentos, la P-glicoproteína es la más estudiada.[14]
En las bacterias gram-negativas, los transportadores ABC secretan los sustratos a través de las membranas interior y exterior sin contacto con el periplasma. A este mecanismo de secreción se lo conoce como «secreción de tipo I», y requiere la participación de tres componentes: el exportador ABC, una proteína de fusión de membrana y un factor de membrana exterior. Un ejemplo lo constituye el sistema de secreción de hemolisina en E. coli, formado por el exportador ABC HlyB, la proteína de fusión de la membrana interior HlyD y el factor de membrana exterior TolC.[10]
Mecanismo de transporte en los exportadores
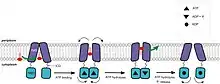
En el estado de reposo, los secretores adoptan la orientación hacia dentro, y los dominios transmembrana (TM) y de unión al nucleótido (ABC) se encuentran a una distancia relativamente amplia para poder acomodar sustratos anfifílicos o hidrófobos. La unión al sustrato marca el inicio del ciclo. La asociación con el ATP, necesaria para la dimerización del dominio ABC y la formación de la casete de ATP, conduce al cambio de confomación del dominio TM a una orientación hacia afuera. La hidrólosis de una molécula de ATP ensancha el orificio hacia el periplasma y empuja el sustrato hacia la lámina externa de la capa bilipídica. Al hidrolizarse la segunda molécula de ATP y liberarse el fosfato, los subdominios ABC se separan y el transportador adopta la configuración de reposo.[33][39][42]
Papel en la farmacorresistencia
Los transportadores ABC desempeñan un papel crucial en el desarrollo de resistencia a múltiples fármacos. Los pacientes afectados experimentan resistencia tanto a la medicación que toman como a otros tipos de fármacos. Esta condición depende de varios factores, uno de los cuales es un incremento de la expulsión del fármaco de la célula por transportadores ABC. Por ejemplo, la proteína ABCB1 (P-glicoproteína) elimina medicamentos antitumorales de la célula.[17] Otro transportador involucrado en el desarrollo de la farmacorresistencia es el ABCG2, también conocido como BCRP (proteína de resistencia al cáncer de mama), que confiere resistencia a la mayoría de los inhibidores de la topoisomerasa I o II, tales como el topotecán, el irinotecán, y la doxorrubicina.[43] ABCC1, prevalente en los neuroblastomas y cáncer en el pulmón, pecho y próstata es un tercer ejemplo.[44]
No está claro exactamente cómo estas proteína pueden trasladar una variedad tan amplia de fármacos. Según una teoría conocida como «modelo de aspiradora hidrofóbica» los fármacos son extraídos indiscriminadamente de la fase lipídica según su hidrofobia.
Para resolver los problemas asociados con la farmacorresistencia, se pueden usar medicamentos que no sean sustratos de los transportadores ABC o moléculas capaces de inhibir los transportadores ABC. Entre los primeros se cuentan la ciclofosfamida, ciertos antimetabolitos como el 5-fluorouracilo, y derivados de la antraciclina, como la annamicina y la doxorubicina. Los inhibidores de los transportadores ABC se usan conjuntamente con los fármacos habituales. Los sustratos que revierten la resistencia a los anticancerosos se denominan quimiosensibilizadores.[4]
Subfamilias
ABCA
En el ser humano, la subfamilia ABCA se compone de doce transportadores divididos a su vez en dos subgrupos. El primer subgrupo consta de siete miembros:ABCA1, ABCA2, ABCA3, ABCA4, ABCA7, ABCA12 y ABCA13, productos de genes repartidos en seis cromosomas. El otro subgrupo está formado por ABCA5, ABCA6, ABCA8, ABCA9 y ABCA10, que forman un grupo de genes en el cromosoma 17. Los genes en el segundo subgrupo se distinguen de los del primero en tener 37-38 exones en lugar de los 50 exones en ABCA1. El subgrupo ABCA1 está implicado en varias enfermedades genéticas. Por ejemplo, la enfermedad de Tangier surge por una mutación en la proteína ABCA1. Varias enfermedades oculares se asocian a mutaciones de ABCA4, como la enfermedad de Stargardt, así como la retinosis pigmentaria con herencia recesiva y la mayoría de distrofias recesivas de conos y bastones.
ABCB
La subfamilia ABCB se compone de cuatro transportadores completos y dos dominos de transportadores. ABCB1 está sobreexpresada en ciertas células tumorales resistentes a fármacos. Se expresa principalmente en la barrera hematoencefálica y en el hígado y se cree que su función es proteger a las células de las toxinas.[5]
ABCC
La subfamilia ABCC contiene trece miembros de los que nueve se conocen como «proteínas de farmacoresistencia múltiple» (MRP). Las proteínas MRP se encuentran en todos los seres vivos y median muchas funciones importantes.[45] Se sabe que están implicadas en el transporte de iones, la secreción de toxinas y la transducción de señales.[5] Cuatro de las proteínas MRP (MRP4, MRP5, MRP8 y MRP9 o ABCC4, ABCC5, ABCC11 y ABCC12, llamadas MPR cortas, presentan una estructura ABC típica con cuatro dominios —dos dominios transmembrana, seguidos cada uno de un dominio de unión al nucleótido—. Las cinco MRP restantes (MRP1, MRP2, MRP6, MRP7 o ABCC1, ABCC2, ABCC3, ABCC6 yABCC 10) se conocen como MRP largas y cuentan con un quinto dominio adicional en su extremo aminoterminal.[45] El regulador de la conductancia transmembrana de la fibrosis quística, cuya mutación y pérdida de función causa fibrosis quística, se considera parte de esta subfamilia.[5] Los receptores de sulfonilurea (SUR), implicados en la secreción de insulina, la función neuronal y la función muscular, también forman parte de esta familia de proteínas. Las mutaciones en las proteínas SUR son una causa potencial de diabetes mellitus neonatal. SUR es también el sitio de unión para fármacos como las sulfonilureas y ciertos activadores de apertura de canales de potasio como el diazóxido.
ABCD
La subfamilia ABCD consta de los productos de cuatro genes, expresados exclusivamente en el peroxisoma. La ABCD1 está asociada a una forma de la adrenoleucodistrofia (ALD) ligada al cromosoma X , enfermedad caracterizada por la neurodegeneración y la deficiencia suprarrenal que generalmente se inicia en la infancia tardía. Las células de los pacientes con ALD presentan una acumulación de ácidos grasos saturados no ramificados, pero el papel exacto de ABCD1 en el proceso aún no está determinado. La función de otros miembros de la subfamilia ABCD tampoco se ha dilucidado aún, pero se piensa que están relacionadas con el metabolismo de los ácidos grasos.[5]
ABCE y ABCF
Ambos subgrupos constan de proteínas parecidas a los dominios de unión al nucleótido de los transportadores completos, pero que carecen de dominio transmembrana. De la subfamilia ABCE solo se conoce un miembro, OABP or ABCE1, que reconoce algunos oligodendrocitos producidos en respuesta a ciertas infecciones virales.[5]
ABCG
La subfamilia ABCG está compuesta por seis transportadores con sitios de unión al ATP en el extremo aminoterminal y dominios transmembrana en el extremo carboxilo, al contrario que las demás subfamilias de transportadores ABC. El ser humano solo tiene cinco genes pertenecientes a este grupo, pero el genoma de la Drosophila contiene quince, y el de las levaduras, diez. La proteína ABCG2 se halló en células resistentes a la mitoxantrona en la ausencia de ABCB1 y ABCC1. ABCG2 es capaz de exportar antraciclinas usadas contra el cáncer, así como topotecán, mitoxantrona, o doxorubicina. El incremento de la farmacorrestencia asociada a la ABCG2 tiene su origen en translocaciones cromosómicas, y se desconoce cuál es la función habitual de esta proteína.[5]
Referencias
- Jones PM, George AM (Mar 2004). «The ABC transporter structure and mechanism: perspectives on recent research». Cellular and Molecular Life Sciences 61 (6): 682-699. PMID 15052411. doi:10.1007/s00018-003-3336-9.
- Ponte-Sucre A, ed. (2009). ABC Transporters in Microorganisms. Caister Academic Press. ISBN 978-1-904455-49-3.
- Davidson AL, Dassa E, Orelle C, Chen J (Jun 2008). «Structure, function, and evolution of bacterial ATP-binding cassette systems». Microbiology and Molecular Biology Reviews 72 (2): 317-64, table of contents. PMC 2415747. PMID 18535149. doi:10.1128/MMBR.00031-07.
- Choi CH (Oct 2005). «ABC transporters as multidrug resistance mechanisms and the development of chemosensitizers for their reversal». Cancer Cell International 5: 30. PMC 1277830. PMID 16202168. doi:10.1186/1475-2867-5-30.
- Dean M, Hamon Y, Chimini G (Jul 2001). «The human ATP-binding cassette (ABC) transporter superfamily». Journal of Lipid Research 42 (7): 1007-1017. PMID 11441126.
- Henderson DP, Payne SM (Nov 1994). «Vibrio cholerae iron transport systems: roles of heme and siderophore iron transport in virulence and identification of a gene associated with multiple iron transport systems». Infection and Immunity 62 (11): 5120-5125. PMC 303233. PMID 7927795.
- Cangelosi GA, Ankenbauer RG, Nester EW (Sep 1990). «Sugars induce the Agrobacterium virulence genes through a periplasmic binding protein and a transmembrane signal protein». Proceedings of the National Academy of Sciences of the United States of America 87 (17): 6708-6712. PMC 54606. PMID 2118656. doi:10.1073/pnas.87.17.6708.
- Kemner JM, Liang X, Nester EW (Abr 1997). «The Agrobacterium tumefaciens virulence gene chvE is part of a putative ABC-type sugar transport operon». Journal of Bacteriology 179 (7): 2452-2458. PMC 178989. PMID 9079938.
- Poolman B, Spitzer JJ, Wood JM (Nov 2004). «Bacterial osmosensing: roles of membrane structure and electrostatics in lipid-protein and protein-protein interactions». Biochimica et Biophysica Acta 1666 (1–2): 88-104. PMID 15519310. doi:10.1016/j.bbamem.2004.06.013.
- Davidson AL, Chen J (2004). «ATP-binding cassette transporters in bacteria». Annual Review of Biochemistry 73: 241-268. PMID 15189142. doi:10.1146/annurev.biochem.73.011303.073626.
- Poole RK, Gibson F, Wu G (Abr 1994). «The cydD gene product, component of a heterodimeric ABC transporter, is required for assembly of periplasmic cytochrome c and of cytochrome bd in Escherichia coli». FEMS Microbiology Letters 117 (2): 217-223. PMID 8181727. doi:10.1111/j.1574-6968.1994.tb06768.x.
- Goffeau A, de Hertogh B, Baret PV (2013). «ABC Transporters». En Lane, William J.; Lennarz, M. Daniel, eds. Encyclopedia of Biological Chemistry (2.ª edición). London: Academic Press. ISBN 978-0-12-378631-9. doi:10.1016/B978-0-12-378630-2.00224-3.
- Gedeon C, Behravan J, Koren G, Piquette-Miller M (2006). «Transport of glyburide by placental ABC transporters: implications in fetal drug exposure». Placenta 27 (11–12): 1096-1102. PMID 16460798. doi:10.1016/j.placenta.2005.11.012.
- Pohl A, Devaux PF, Herrmann A (Mar 2005). «Function of prokaryotic and eukaryotic ABC proteins in lipid transport». Biochimica et Biophysica Acta 1733 (1): 29-52. PMID 15749056. doi:10.1016/j.bbalip.2004.12.007.
- Randolph GJ (2001). «Dendritic cell migration to lymph nodes: cytokines, chemokines, and lipid mediators». Seminars in Immunology 13 (5): 267-274. PMID 11502161. doi:10.1006/smim.2001.0322.
- Shuman HA (1982). «Active transport of maltose in Escherichia coli K12. Role of the periplasmic maltose-binding protein and evidence for a substrate recognition site in the cytoplasmic membrane». J. Biol. Chem. 257 (10): 5455-5461.
- Scott MP, Lodish HF, Berk A, Kaiser, C, Krieger M, Bretscher A, Ploegh H, Amon A (2012). Molecular Cell Biology. San Francisco: W. H. Freeman. ISBN 1-4292-3413-X.
- Rees DC, Johnson E, Lewinson O (Mar 2009). «ABC transporters: the power to change». Nature Reviews Molecular Cell Biology 10 (3): 218-227. PMC 2830722. PMID 19234479. doi:10.1038/nrm2646.
- Hollenstein K, Frei DC, Locher KP (Mar 2007). «Structure of an ABC transporter in complex with its binding protein». Nature 446 (7132): 213-216. PMID 17322901. doi:10.1038/nature05626.
- Locher KP, Lee AT, Rees DC (mayo de 2002). «The E. coli BtuCD structure: a framework for ABC transporter architecture and mechanism». Science 296 (5570): 1091-1098. PMID 12004122. doi:10.1126/science.1071142.
- Pinkett HW, Lee AT, Lum P, Locher KP, Rees DC (Ene 2007). «An inward-facing conformation of a putative metal-chelate-type ABC transporter». Science 315 (5810): 373-377. PMID 17158291. doi:10.1126/science.1133488.
- Moody JE, Millen L, Binns D, Hunt JF, Thomas PJ (Jun 2002). «Cooperative, ATP-dependent association of the nucleotide binding cassettes during the catalytic cycle of ATP-binding cassette transporters». The Journal of Biological Chemistry 277 (24): 21111-21114. PMC 3516282. PMID 11964392. doi:10.1074/jbc.C200228200.
- Hung LW, Wang IX, Nikaido K, Liu PQ, Ames GF, Kim SH (Dec 1998). «Crystal structure of the ATP-binding subunit of an ABC transporter». Nature 396 (6712): 703-707. PMID 9872322. doi:10.1038/25393.
- Verdon G, Albers SV, Dijkstra BW, Driessen AJ, Thunnissen AM (Jul 2003). «Crystal structures of the ATPase subunit of the glucose ABC transporter from Sulfolobus solfataricus: nucleotide-free and nucleotide-bound conformations». Journal of Molecular Biology 330 (2): 343-358. PMID 12823973. doi:10.1016/S0022-2836(03)00575-8.
- Karpowich N, Martsinkevich O, Millen L, Yuan YR, Dai PL, MacVey K, Thomas PJ, Hunt JF (Jul 2001). «Crystal structures of the MJ1267 ATP binding cassette reveal an induced-fit effect at the ATPase active site of an ABC transporter». Structure 9 (7): 571-586. PMID 11470432. doi:10.1016/S0969-2126(01)00617-7.
- Chen J, Lu G, Lin J, Davidson AL, Quiocho FA (Sep 2003). «A tweezers-like motion of the ATP-binding cassette dimer in an ABC transport cycle». Molecular Cell 12 (3): 651-661. PMID 14527411. doi:10.1016/j.molcel.2003.08.004.
- Diederichs K, Diez J, Greller G, Müller C, Breed J, Schnell C, Vonrhein C, Boos W, Welte W (Nov 2000). «Crystal structure of MalK, the ATPase subunit of the trehalose/maltose ABC transporter of the archaeon Thermococcus litoralis». The EMBO Journal 19 (22): 5951-5961. PMC 305842. PMID 11080142. doi:10.1093/emboj/19.22.5951.
- Gaudet R, Wiley DC (Sep 2001). «Structure of the ABC ATPase domain of human TAP1, the transporter associated with antigen processing». The EMBO Journal 20 (17): 4964-4972. PMC 125601. PMID 11532960. doi:10.1093/emboj/20.17.4964.
- Schmitt L, Benabdelhak H, Blight MA, Holland IB, Stubbs MT (Jul 2003). «Crystal structure of the nucleotide-binding domain of the ABC-transporter haemolysin B: identification of a variable region within ABC helical domains». Journal of Molecular Biology 330 (2): 333-342. PMID 12823972. doi:10.1016/S0022-2836(03)00592-8.
- Yuan YR, Blecker S, Martsinkevich O, Millen L, Thomas PJ, Hunt JF (Ago 2001). «The crystal structure of the MJ0796 ATP-binding cassette. Implications for the structural consequences of ATP hydrolysis in the active site of an ABC transporter». The Journal of Biological Chemistry 276 (34): 32313-32321. PMID 11402022. doi:10.1074/jbc.M100758200.
- Smith PC, Karpowich N, Millen L, Moody JE, Rosen J, Thomas PJ, Hunt JF (Jul 2002). «ATP binding to the motor domain from an ABC transporter drives formation of a nucleotide sandwich dimer». Molecular Cell 10 (1): 139-149. PMC 3516284. PMID 12150914. doi:10.1016/S1097-2765(02)00576-2.
- Dawson RJ, Locher KP (Sep 2006). «Structure of a bacterial multidrug ABC transporter». Nature 443 (7108): 180-185. PMID 16943773. doi:10.1038/nature05155.
- Ward A, Reyes CL, Yu J, Roth CB, Chang G (Nov 2007). «Flexibility in the ABC transporter MsbA: Alternating access with a twist». Proceedings of the National Academy of Sciences of the United States of America 104 (48): 19005-19010. PMC 2141898. PMID 18024585. doi:10.1073/pnas.0709388104.
- Fetsch EE, Davidson AL (Jul 2002). «Vanafecha-catalyzed photocleavage of the signature motif of an ATP-binding cassette (ABC) transporter». Proceedings of the National Academy of Sciences of the United States of America 99 (15): 9685-9690. PMC 124977. PMID 12093921. doi:10.1073/pnas.152204499.
- Ambudkar SV, Kim IW, Xia D, Sauna ZE (Feb 2006). «The A-loop, a novel conserved aromatic acid subdomain upstream of the Walker A motif in ABC transporters, is critical for ATP binding». FEBS Letters 580 (4): 1049-1055. PMID 16412422. doi:10.1016/j.febslet.2005.12.051.
- Geourjon C, Orelle C, Steinfels E, Blanchet C, Deléage G, Di Pietro A, Jault JM (Sep 2001). «A common mechanism for ATP hydrolysis in ABC transporter and helicase superfamilies». Trends in Biochemical Sciences 26 (9): 539-544. PMID 11551790. doi:10.1016/S0968-0004(01)01907-7.
- Ye J, Osborne AR, Groll M, Rapoport TA (Nov 2004). «RecA-like motor ATPases--lessons from structures». Biochimica et Biophysica Acta 1659 (1): 1-18. PMID 15511523. doi:10.1016/j.bbabio.2004.06.003.
- Zaitseva J, Jenewein S, Jumpertz T, Holland IB, Schmitt L (Jun 2005). «H662 is the linchpin of ATP hydrolysis in the nucleotide-binding domain of the ABC transporter HlyB». The EMBO Journal 24 (11): 1901-1910. PMC 1142601. PMID 15889153. doi:10.1038/sj.emboj.7600657.
- Higgins CF, Linton KJ (Oct 2004). «The ATP switch model for ABC transporters». Nature Structural & Molecular Biology 11 (10): 918-926. PMID 15452563. doi:10.1038/nsmb836.
- Senior AE, al-Shawi MK, Urbatsch IL (Dec 1995). «The catalytic cycle of P-glycoprotein». FEBS Letters 377 (3): 285-289. PMID 8549739. doi:10.1016/0014-5793(95)01345-8.
- Oldham ML, Davidson AL, Chen J (Dic 2008). «Structural insights into ABC transporter mechanism». Current Opinion in Structural Biology 18 (6): 726-733. PMC 2643341. PMID 18948194. doi:10.1016/j.sbi.2008.09.007.
- Gutmann DA, Ward A, Urbatsch IL, Chang G, van Veen HW (Ene 2010). «Understanding polyspecificity of multidrug ABC transporters: closing in on the gaps in ABCB1». Trends in Biochemical Sciences 35 (1): 36-42. PMC 4608440. PMID 19819701. doi:10.1016/j.tibs.2009.07.009.
- Leonard GD, Fojo T, Bates SE (2003). «The role of ABC transporters in clinical practice». The Oncologist 8 (5): 411-424. PMID 14530494. doi:10.1634/theoncologist.8-5-411.
- Munoz, Marcia; Henderson, Michelle; Haber, Michelle; Norris, Murray (2007). «Role of the MRP1/ABCC1 Multidrug Transporter Protein in Cancer». IUBMB Life 59 (12): 752-757. doi:10.1080/15216540701736285.
- Chen ZS, Tiwari AK (Sep 2011). «Proteínas de resistencia a múltiples fármacos (MRP / ABCC) en quimioterapia contra el cáncer y enfermedades genéticas». La revista FEBS 278: 3226-3245. PMC 3168698. PMID 21740521. doi:10.1111/j.1742-4658.2011.08235.x.
Enlaces externos
 Wikimedia Commons alberga una categoría multimedia sobre Transportadores ABC.
Wikimedia Commons alberga una categoría multimedia sobre Transportadores ABC.
- Clasificación de los transportadores ABC en la base de datos de clasificación de transportadores.
| Control de autoridades |
|
|---|
 Datos: Q286958
Datos: Q286958- Multimedia: ABC transporters / Q286958