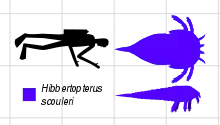
Hibbertopterus
Hibbertopterus est un genre éteint d'euryptérides de grande taille, ayant existé du Dévonien au Carbonifère, il y a entre 387,7 et 323,2 millions d'années. Contrairement à la majorité des euryptérides, il vivait en eau douce. L'espèce type, H. scouleri, a été nommée en tant qu'espèce appartenant au genre Eurypterus par Samuel Hibbert en 1836. Le nom générique Hibbertopterus, inventé plus d'un siècle plus tard, associe le nom du scientifique au mot grec πτερόν (pteron) qui signifie "aile".

| Règne | Animalia |
|---|---|
| Embranchement | Arthropoda |
| Sous-embr. | Chelicerata |
| Ordre | † Eurypterida |
| Famille | † Hibbertopteridae |
Des fossiles d'Hibbertopterus ont été découverts dans des gisements de la période dévonienne en Belgique, en Écosse et aux États-Unis et dans des gisements de la période carbonifère en Écosse, en Irlande, en République tchèque et en Afrique du Sud.
Taille
Hibbertopterus était le plus gros euryptéride du sous-ordre des Stylonurina. L'espèce type H. scouleri pouvait mesurer de 1,8 à 2 m de long. Une espèce sud-africaine, H. wittebergensis, pouvait atteindre 2,5 m de long. C'est l'un des plus grands arthropodes de l'histoire, et il avait une taille comparable à celle des myriapodes diplopode géants (du genre Arthropleura) ou d'autres euryptérides (du genre Pterygotus, et Jaekelopterus rhenaniae).
Biologie
_(7394051300).jpg.webp)
Hibbertopterus était un animal fouisseur, dont le terrain de chasse était les couches superficielles des lits de rivières et autres cours d'eau. Il utilisait ses appendices spécialisés orientés vers l'avant, munis de plusieurs épines, pour ratisser le substrat des environnements dans lesquels il vivait, à la recherche de petits invertébrés à manger, qu'il pouvait ensuite amener vers sa bouche. Bien que l'hypothèse soit ancienne, le fait que les euryptérides soient capables de locomotion terrestre n'a été prouvé qu'en 2005 par la découverte d'une piste fossile de Hibbertopterus en Écosse[1]. La piste a montré qu’un animal mesurant environ 1,6 mètre avait lentement gravi une étendue de terre, traînant son telson sur le sol. La manière dont Hibbertopterus pouvait survivre sur la terre ferme, même brièvement, reste inconnue. Il se pourrait que l'animal était doté de branchies capables de fonctionner dans l'air aussi longtemps qu'elles étaient mouillées, ou qu'il possédait un double système respiratoire, probablement présent chez quelques autres euryptérides.
Autrefois considérés comme des genres distincts, Hibbertopterus, Cyrtoctenus et Dunsopterus représentaient en réalité la même espèce, mais à des stades de développements différents[2],[3],[4]. En parallèle de l'augmentation de la taille de l'individu, ainsi que de la migration des yeux vers le sommet de la tête, les appendices buccaux subissaient des transformations les modifiant de simples appendices ratisseurs permettant de pousser de grosses proies vers la bouche, en véritables filtres qui auraient permis la capture de proies aussi petites que du zooplancton.
Description
Comme d'autres euryptérides connus, Hibbertopterus était très grand et large. C'était le plus grand euryptéridé connu du sous-ordre des Stylonurina, composé de ces euryptérides dépourvus de palettes de natation. Une carapace (la partie de l'exosquelette recouvrant la tête), qui fait référence à l'espèce H. scouleri, originaire du Carbonifère et découverte en actuelle Écosse, mesure 65 cm de large. Étant donné que Hibbertopterus était exceptionnellement large par rapport à sa longueur pour un euryptéride, l'animal en question aurait probablement mesuré environ 1,8 à 2 m de longueur. Même s'il y avait des euryptérides de plus grande longueur (tels que Jaekelopterus, Pentecopterus Pterygotus ), Hibbertopterus était très profond et compact, comparé à d'autres euryptérides et la masse du spécimen en question aurait probablement rivalisé avec celle des autres euryptérides géants (et d'autres arthropodes, actuels ou éteints), s’ils ne sont pas surpassés[5]. Outre les découvertes de fossiles de grands spécimens, des traces de fossiles attribuées à l'espèce H. wittebergensis d'Afrique du Sud indiquent un animal d'environ 2,5 m de longueur (la même taille attribuée au plus grand euryptéride connu, Jaekelopterus ), bien que les plus grands spécimens fossiles connus de l’espèce ne semblent avoir atteint qu'une longueur de 1,35 m.
Les appendices d'Hibbertopterus faisant face à l'avant étaient spécialisés dans la collecte d'aliments. Les podomères distaux (segments de jambe) de ces trois paires de membres étaient couverts de longues épines, et l'extrémité de chaque membre était recouverte d'organes sensoriels. Ces adaptations suggèrent que Hibbertopterus, à l'instar des autres hibbertoptérides (Hibbertopteridae (en) de leur nom scientifique), se serait nourri selon une méthode appelée alimentation par balayage, utilisant ses membres pour balayer le substrat de son environnement à la recherche de nourriture. La quatrième paire d'appendices, bien qu'utilisée dans l'alimentation comme les deuxième et troisième paires, était également utilisée pour la locomotion et les deux dernières paires de pattes (paires cinq et six) étaient uniquement locomotrices. Hibbertopterus aurait eu une démarche hexapodale[2]. Bien que l'on ne connaisse pas assez de matériaux fossiles chez les autres euryptérides de la même famille pour discuter de leurs différences en toute certitude[6], Hibbertopterus est défini sur la base d'un ensemble de caractéristiques définies. Le telson (la "pointe de la queue") était grand et avait la forme d'un glaive ou d'une épée romaine. Sa quille coulait au centre, avec à son tour une petite empreinte en son centre. Les pattes d'Hibbertopterus avaient des extensions à leurs bases et manquaient de rainures postérieures longitudinales dans tous leurs podomères (segments de jambe)[2]. Certaines de ces caractéristiques, en particulier la forme du telson, auraient été partagées par d'autres hibbertoptérides, beaucoup moins bien conservés que Hibbertopterus lui-même.
Tableau des espèces
Les paléontologues allemands Jason A. Dunlop et Denise Jekel et le paléontologue britannique David Penney font état de la situation des 10 espèces énumérées ci-dessous [7] et celle des gammes temporelles[pas clair] d'une étude réalisée en 2009 par les paléontologues américains James Lamsdell et Simon J. Braddy sauf indication contraire. Les caractéristiques distinctives de H. caledonicus, H. dewalquei, H. ostraviensis et H. peachi sont conformes à la description de 1968 de ces espèces[8]. Les descripteurs, le paléontologue norvégien Leif Størmer et le paléontologue britannique Charles D. Waterston, ne considéraient pas que ces espèces représentaient des euryptéridés, bien qu'aucun diagnostic corrigé de celles-ci n'ait encore été publié.
| Species | Auteur | Année | Statut | Longueur | Période géologique | Notes & description |
|---|---|---|---|---|---|---|
| Hibbertopterus caledonicus | Salter | 1863 | Valide | ? | Viséan (Carboniferous) | Rachis isolé fossile trouvé en Ecosse. H. caledonicus se distingue par les longs filaments (plus longs que ceux de H. peachi) de sa deuxième paire d'appendices et par la présence de petits fulcra (tissu de soutien) de forme conique le long des membres[8]. |
| Hibbertopterus dewalquei | Fraipont | 1889 | Valide | ? | Famennian (Devonian) | Rachis isolé fossile trouvé en Belgique. Le trait distinctif de H. dewalquei est la présence de ~110 filaments dans chaque rangée de ses organes de nutrition en forme de peigne. |
| Hibbertopterus dicki | Peach | 1883 | Valide | ? | Givetian (Devonian) | Tergites isolés fossiles trouvés en Ecosse. Le matériel fossile de H. dicki est trop limité pour établir avec certitude des caractéristiques distinctives, mais il peut être attribué avec certitude au genre sur la base de ses "similarités frappantes" avec les autres espèces[9]. |
| Hibbertopterus hibernicus | Baily | 1872 | Classification contesté | ? | Tournaisian (Carboniferous) | Le seul spécimen connu, découvert en Irlande, est probablement perdu et a été initialement attribué à Pterygotus. Il a été référencé à Hibbertopterus en 1964 sans que l'on sache pourquoi[10]. |
| Hibbertopterus ostraviensis | Augusta & Přibyl | 1951 | Valide | ? | Bashkirian (Carboniferous) | Rachis isolé fossile trouvé en République Tchèque. Le matériel fossile de H. ostraviensis est trop limité pour établir avec certitude des caractéristiques distinctives. |
| Hibbertopterus peachi | Størmer & Waterston | 1968 | Valide | ? | Viséan (Carboniferous) | Rachis isolé fossile trouvé en Ecosse. Les caractéristiques distinctives de H. peachi sont les filaments en grand nombre sur sa première paire d'appendices et les filaments plus longs sur la deuxième paire d'appendices, qui ont aussi de grands fulcra. |
| Hibbertopterus scouleri | Hibbert | 1836 | Valide | 180 cm | Viséan (Carboniferous) | L'estimation de la taille est basée sur une carapace complète. En tant qu'espèce type, aucun caractère distinctif particulier n'est établi pour H. scouleri. Sa dernière diagnose publiée est simplement la même que celle du genre lui-même. |
| Hibbertopterus stevensoni | Etheridge Jr. | 1877 | Valide | 80 cm | Viséan (Carboniferous) | Fossiles fragmentaires connus en Ecosse. Connu principalement à partir des segments de pattes, le principal trait distinctif de H. stevesoni est la disposition précise des lunules (marques en forme de croissant) sur lesdits segments de pattes, formant des rangées. Ceci est différent de, par exemple, H. peachi, chez qui ces lunules sont plus nombreuses et moins bien triées en rangées. |
| Hibbertopterus wittebergensis | Waterston, Oelofsen & Oosthuizen | 1985 | Valide | 135 cm (250 cm) |
Tournaisian (Carboniferous) | L'estimation de taille inférieure suit les preuves fossiles connues. L'estimation de taille supérieure dérive des preuves de traces.
H. wittebergensis peut être distingué des autres espèces par la forme ovale de son nœud ocellaire (la partie surélevée de la carapace sur laquelle se trouvent les ocelles) et par les nombreuses crénulations présentes sur les marges distales de ses segments de pattes[11]. |
| Hibbertopterus wrightianus | Dawson | 1881 | Classification contesté | 120 cm | Famennian (Devonian) | Des podomères isolés d'un appendice prosomal ont été découverts aux Etats-Unis. Le matériel fossile de H. wrightianus est trop limité pour établir avec certitude des caractéristiques distinctives, mais il est noté qu'il est similaire à H. stevensoni[12]. |
Histoire de la recherche
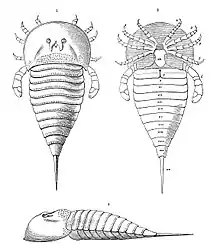
En 1831, le naturaliste écossais John Scouler décrivit la dépouille, composée d'un prosome massif et inhabituel et de plusieurs tergites, d'un grand et étrange arthropode découvert dans des gisements du Carbonifère inférieur, situés en Écosse, sans donner de nom aux fossiles. D'après l'examen de Scouler, les fossiles constituent le deuxième euryptéride à faire l'objet d'une étude scientifique, six ans après la description d' Eurypterus en 1825. Cinq ans plus tard, en 1836, le géologue britannique Samuel Hibbert a redécrit les mêmes spécimens de fossiles, en les appelant Eurypterus scouleri[13].
Le géologue britannique Ben Peach a donné le nom de genre euryptérides Glyptoscorpius à l'espèce G. perornatus (considérée comme espèce type de Glyptoscorpius par des chercheurs ultérieurs bien qu'elle n'ait pas été désignée comme telle à l'origine) en 1882. Le genre était basé sur G. perornatus et l'espèce fragmentaire G. caledonicus, précédemment décrite comme la plante Cycadites caledonicus par le paléontologue anglais John William Salter en 1863. Cette désignation a été renforcée avec davantage de fragments de fossiles découverts dans le brûlage de Coomsdon, dont Peach faisait référence à Glyptoscorpius caledonicus. En 1887, Peach décrivit G. minutisculptus de Mount Vernon, Glasgow et G. kidstoni de Radstock à Somerset. Glyptoscorpius de Peach est très problématique : certaines des caractéristiques diagnostiques utilisées lors de la description sont discutables ou dénuées de sens. Par exemple, la description originale était basée sur G. caledonicus et G. perornatus, mais comme les parties du corps conservées dans les fossiles décrits ne se chevauchent pas complètement, il est impossible de dire si les caractéristiques diagnostiques de Peach s'appliquent réellement aux deux espèces originales[8].

Bien que représentée seulement par deux petits fragments de fossile articulés et vaguement cylindriques, l'espèce reconnue aujourd'hui comme H. wrightianus a eu une histoire taxonomique compliquée. Décrite à l'origine en 1881 comme une espèce de plante, le fossile fragmentaire appelé " Equisetides wrightiana " était réputé représenter les restes fossiles d'un eurypteride par le paléontologue américain James Hall en 1884, trois ans plus tard. Bien que Hall attribue l'espèce à Stylonurus, cette même année les paléontologues britanniques Henry Woodward et Thomas Rupert Jones ont attribué le fossile au genre Echinocaris, croyant que les fossiles représentaient un phyllocarida crustacé. L'affectation à Echinocaris était probablement basée sur la surface légèrement épineuse des fossiles, mais en 1888, Hall et le paléontologue américain John Mason Clarke signalèrent qu'aucun Echinocaris décrit n'avait réellement d'épines semblables à celles suggérées par Woodward et Jones et à Stylonurus, interprétant les fossiles comme des fragments des longues jambes qui marchent. Une cession à Stylonurus a été confirmée par Clarke et paléontologue américain Rudolf Ruedemann leur influence Le eurypterida de New York en 1912, mais pas de traits distinctifs des fossiles ont été donnés en raison de leur caractère fragmentaire[14].
Pterygotus hibernicus (une espèce décrite en Irlande par le paléontologue britannique William Hellier Baily en 1872) a été réaffecté à Hibbertopterus par le paléontologue américain Erik N. Kjellesvig-Waering en 1964 dans le cadre d'un réexamen plus approfondi de les différentes espèces assignées à la famille des Pterygotidae. Kjellesvig-Waering a retenu P. dicki dans Pterygotus[10]. Les paléontologues écossais Lyall I. Anderson et Nigel H. Trewin et le paléontologue allemand Jason A. Dunlop ont noté en 2000 que l'acceptation par Kjellesvig-Waerings de la désignation initiale de Pterygotus dicki était "fastidieuse" car elle reposait sur un matériel extrêmement fragmentaire. P. dicki, comme de nombreuses autres espèces de ptérygotides, représente un autre nom qui s’applique à certains segments épars, une pratique qu’ils considèrent comme "insoutenable en taxonomie". Bien qu'ils aient suggéré que des recherches supplémentaires soient nécessaires pour déterminer si le taxon était valide ou non, ils ont toutefois noté que la présence d'une frange sur les segments formés par leur ornementation était absente chez toutes les autres espèces de Pterygotus, mais "remarquablement similaire". à ce qui était présent dans Cyrtoctenus[9]. Des recherches ultérieures ont traité P. dicki comme une espèce de Cyrtoctenus[7].
Lorsque Kjellesvig-Waering désigna le genre Hibbertopterus en 1959, Eurypterus scouleri avait déjà été référé au Campylocephalus apparenté pendant un certain temps. Kjellesvig-Waering a reconnu Campylocephalus scouleri comme distincte de l'espèce type de ce genre, C. oculatus, en ce que le prosome de Campylocephalus était plus étroit, avait une forme subelliptique (presque elliptique) et avait son point le plus large au milieu plutôt qu'au centre base. D'autres différences ont été notées dans la position et la forme des yeux composés de l'animal, qui chez Hibbertopterus sont entourés d'une forme annulaire de tégument durci (absent chez Campylocephalus). Les yeux d' Hibbertopterus sont également situés près du centre de la tête, tandis que ceux de Campylocephalus sont situés plus en arrière. Le nom générique Hibbertopterus a été choisi pour honorer le descripteur original de H. scouleri, Samuel Hibbert[15].
_(7394046982).jpg.webp)
Classification
Hibbertopterus fait partie de la famille Hibbertopteridae, à laquelle il prête également son nom, une famille d'euryptérides de la superfamille Mycteropoidea, aux côtés des genres Campylocephalus et Vernonopterus. Les hibbertoptérides forment un groupe constitué de gros myctéropoïdes à larges prosomes, un telon hastate semblable à celui d'Hibbertopterus, ornement constitué d'écailles ou d'autres structures similaires sur l'exosquelette, le quatrième couple d'appendices possédant des épines, les tergites postérieures de l'abdomen possédant des écailles en forme de langue près de leurs bords et des lobes positionnés de manière potérolatérale (postérieure des deux côtés) sur le prosome[6]. Historiquement, la morphologie de Hibbertopterus et des autres hibbertoptérides a été considérée comme si inhabituelle qu’on a pensé qu’il s’agissait d’un ordre distinct de celui d’ Eurypterida[16]. Les caractéristiques de Campylocephalus et de Vernonopterus indiquent clairement que les deux genres représentent des euryptéridés hibbertoptérides, mais la nature incomplète de tous les spécimens fossiles qui leur sont référés rend difficile toute étude ultérieure des relations phylogénétiques précises entre les Hibbertopteridae. Les deux genres pourraient même représenter des synonymes d'Hibbertopterus lui-même, bien que la nature très incomplète de leurs restes rende cette hypothèse impossible à confirmer[6].
Paléoécologie

Les hibbertoptérides tels que Hibbertopterus étaient des animaux balayeurs, ayant des épines modifiées sur leurs appendices prosomaux orientés vers l'avant qui leur permettaient de ratisser le substrat de leur milieu de vie. Bien que l’alimentation par balayage soit utilisée comme stratégie par de nombreux genres au sein de la Stylonurina, elle s’est surtout développée au sein des Hibbertopteridae, qui possèdent des pales aux deuxième, troisième et quatrième paires d’appendices[6]. Hibbertopterus et d'autres balayeurs se nourrissaient probablement de ce qu'ils pouvaient trouver dans leur environnement de vie, probablement de petits invertébrés[17]. Cette méthode d'alimentation est assez similaire à l'alimentation par filtre. Ceci a conduit certains chercheurs à penser qu'Hibbertopterus aurait été un animal pélagique, en tant que crustacé filtreur moderne, mais la nature robuste et massive du genre (contrairement aux crustacés modernes filtrants qui sont généralement très petits) rend peu probable une telle conclusion[11].
Dans la vallée écossaise du Midland, à 27 kilomètres à l'ouest d'Edimbourg, la carrière d'East Kirkton contient des dépôts qui étaient autrefois un lac d'eau douce près d'un volcan. La localité a préservé une faune diversifiée de l'âge viséen du Carbonifère (il y a environ 335 millions d'années). Autre que H. scouleri, la faune comprend plusieurs animaux terrestres, tels que anthracosaures, aistopodes, baphetids et temnospondyles, ce qui représente quelques - uns des plus anciens terrestres connus tétrapodes. Plusieurs invertébrés terrestres sont également connus, y compris plusieurs espèces de diplopoda, Gigantoscorpio (l’un des plus anciens scorpions avérés terrestres) et les premiers représentants des Opilion. Le site préserve également une vie végétale abondante, y compris les genres Lepidodendron, Lepidophloios, Stigmaria et Sphenopteris . Localement, les étranges carapaces fossiles de H. scouleri ont reçu le nom commun de "heids de Scouler" ("heid" étant en écossais pour "tête")[18].
La formation Waaipoort, où H. wittebergensis a été découverte, conserve également une faune diversifiée du Carbonifère et certaines espèces de plantes. Interprété comme ayant été un grand lac ouvert, d'eau douce à saumâtre, avec éventuellement des influences occasionnelles d'orage et de processus glaciaires, les restes fossiles récupérés sont le plus souvent ceux de divers types de poissons. Parmi ces types figurent les paléoniscoidés, les requins et les acanthodiens. Bien que le matériel de requin soit trop fragmentaire pour être identifiable, au moins certains fossiles pourraient représenter les restes de protacrodontoïdes. Parmi les acanthodiens, au moins trois genres ont été identifiés à partir des écailles fossiles et épines. Outre H. wittebergensis, les seuls invertébrés connus sont deux espèces rares de bivalves, pouvant représenter des unionidés. Les fossiles végétaux de la Formation de Waaiport sont nettement moins diversifiés que ceux des âges précédents au même endroit, probablement pour des raisons climatiques. Praeramunculus (qui représente probablement un progymnosperme) et Archaeosigillaria (un petit type de lycopodes) font partie des genres présents[19].
Notes et références
- (en) Whyte, « Palaeoecology: A gigantic fossil arthropod trackway », Nature, vol. 438, no 7068, , p. 576 (PMID 16319874, DOI 10.1038/438576a, Bibcode 2005Natur.438..576W)
- Jeram, Andrew J.; Selden, Paul A. (1993). "Eurypterids from the Viséan of East Kirkton, West Lothian, Scotland". Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 84 (3-4): 301–308. doi:10.1017/S0263593300006118. ISSN 1755-6929
- (en) Lamsdell, « Redescription of Drepanopterus pentlandicus Laurie, 1892, the earliest known mycteropoid (Chelicerata: Eurypterida) from the early Silurian (Llandovery) of the Pentland Hills, Scotland », Earth and Environmental Science Transactions of the Royal Society of Edinburgh, vol. 103, , p. 77–103 (DOI 10.1017/S1755691012000072, lire en ligne)
- Hughes, Emily Samantha (2019), "Discerning the Diets of Sweep-Feeding Eurypterids Through Analyses of Mesh-Modified Appendage Armature". Graduate Theses, Dissertations, and Problem Reports. 3890
- (en) Tetlie, « Hallipterus excelsior, a Stylonurid (Chelicerata: Eurypterida) from the Late Devonian Catskill Delta Complex, and Its Phylogenetic Position in the Hardieopteridae », Bulletin of the Peabody Museum of Natural History, vol. 49, , p. 19–99 (DOI 10.3374/0079-032X(2008)49[19:HEASCE]2.0.CO;2)
- (en) James C. Lamsdell, Simon J. Braddy & O. Erik Tetlie, « The systematics and phylogeny of the Stylonurina (Arthropoda: Chelicerata: Eurypterida) », Journal of Systematic Palaeontology, vol. 8, no 1, , p. 49–61 (DOI 10.1080/14772011003603564)
- Dunlop, J. A., Penney, D. & Jekel, D. 2018. A summary list of fossil spiders and their relatives. In World Spider Catalog. Natural History Museum Bern
- (en) Waterston et Størmer, « IV. Cyrtoctenus gen. nov., a large late Palaeozoic Arthropod with pectinate Appendages* », Earth and Environmental Science Transactions of the Royal Society of Edinburgh, vol. 68, no 4, , p. 63–104 (ISSN 2053-5945, DOI 10.1017/S0080456800014563)
- (en) Trewin, Dunlop et Anderson, « A Middle Devonian chasmataspid arthropod from Achanarras Quarry, Caithness, Scotland », Scottish Journal of Geology, vol. 36, no 2, , p. 151–158 (ISSN 0036-9276, DOI 10.1144/sjg36020151, lire en ligne)
- Kjellesvig-Waering, « A Synopsis of the Family Pterygotidae Clarke and Ruedemann, 1912 (Eurypterida) », Journal of Paleontology, vol. 38, no 2, , p. 331–361 (ISSN 0022-3360, JSTOR 1301554)
- D. Waterston, W. Oelofsen et D. F. Oosthuizen, « Cyrtoctenus wittebergensis sp. nov. (Chelicerata: Eurypterida), a large sweep-feeder from the Carboniferous of South Africa », Transactions of the Royal Society of Edinburgh: Earth Sciences, vol. 76, nos 2–3, , p. 339–358 (DOI 10.1017/S0263593300010555, lire en ligne)
- (en) Waterston, « I.—Further Observations on the Scottish Carboniferous Eurypterids* », Earth and Environmental Science Transactions of the Royal Society of Edinburgh, vol. 68, no 1, , p. 1–20 (ISSN 2053-5945, DOI 10.1017/S0080456800014472)
- Kjellesvig-Waering, « The Silurian Eurypterida of the Welsh Borderland », Journal of Paleontology, vol. 35, no 4, , p. 789–835 (ISSN 0022-3360, JSTOR 1301214)
- Clarke et Ruedemann, « The Eurypterida of New York », Memoir (New York State Museum and Science Service), vol. 14, (lire en ligne)
- Kjellesvig-Waering, « A Taxonomic Review of Some Late Paleozoic Eurypterida », Journal of Paleontology, vol. 33, no 2, , p. 251–256 (ISSN 0022-3360, JSTOR 1300755)
- Tollerton, V P, « Morphology, Taxonomy, and Classification of the Order Eurypterida Burmeister, 1843 », Journal of Paleontology, vol. 63, no 5, , p. 642–657 (DOI 10.1017/S0022336000041275)
- Selden, P.A., Corronca, J.A. & Hünicken, M.A, « The true identity of the supposed giant fossil spider Megarachne », Biology Letters, vol. 1, no 1, , p. 44–48 (PMID 17148124, PMCID 1629066, DOI 10.1098/rsbl.2004.0272)
- Rolfe, Durant, Fallick et Hall, « An early terrestrial biota preserved by Visean vulcanicity in Scotland », Volcanism and Fossil Biota, Geological Society of America Special Publication, vol. 244, , p. 13–24 (ISBN 9780813722443, DOI 10.1130/SPE244-p13, lire en ligne)
- (en) Evans, « Palaeobiology of Early Carboniferous lacustrine biota of the Waaipoort Formation (Witteberg Group), South Africa », Palaeontologica Africana, vol. 35, , p. 1–6 (lire en ligne)
 Portail de la paléontologie
Portail de la paléontologie  Portail de la zoologie
Portail de la zoologie  Portail de l’Afrique du Sud
Portail de l’Afrique du Sud  Portail de l’Europe
Portail de l’Europe