Histoire évolutive des primates
L'ordre des Primates constituerait l'une des plus anciennes lignées de mammifères placentaires, dont l'apparition daterait de 80 à 65 millions d'années. Issus d'un ancêtre vraisemblablement insectivore (dont aucun fossile n'atteste l'existence à l'heure actuelle) qui évoluait au Crétacé supérieur, les Primates ont connu une radiation évolutive importante tout au long du Cénozoïque. Plusieurs divergences successives ont conduit à la formation des cinq grands ensembles de primates actuels : les lémuriens, les loris et galagos, les tarsiers, les singes du Nouveau Monde (Platyrrhiniens), et ceux de l'Ancien Monde (Catarrhiniens).

Historique
Par une curieuse ironie du sort, il revient au naturaliste français Georges Cuvier l'honneur d'avoir découvert le premier primate fossile. En effet, ce fervent détracteur de la théorie évolutionniste avait déclaré en 1812 : « L'homme fossile n'existe pas ». En 1821, il décrit l'espèce Adapis parisiensis, un fossile extrait des carrières de gypse de Montmartre, qu'il prend par erreur pour un ongulé (Adapis signifie « vers Apis », le taureau sacré égyptien)[1]. La véritable nature de ces ossements ne sera comprise que près de quatre-vingts ans plus tard.
Le premier fossile explicitement attribué à un primate fut découvert en 1836 dans les Siwaliks, en Inde, par deux ingénieurs de l'armée britannique, William E. Baker et Henry Marion Durand. Il s'agissait d'un demi-maxillaire d'un singe de la taille d'un orang-outan, mais dont la dentition était semblable à celle des semnopithèques. Ce fossile est désormais reconnu comme l'holotype de l'espèce Procynocephalus subhimalayanus, un ancêtre probable des macaques.
Un an plus tard, en 1837, le paléontologue Édouard Lartet découvre à Sansan (Gers) les restes d'un grand singe, proche des gibbons. Nommé par la suite Pliopithecus antiquus, ce fossile met la communauté scientifique en émoi puisqu'il semble à la fois confirmer l'idée d'évolution et indiquer que des singes primitifs évoluaient autrefois sous des latitudes tempérées. Devant l'Académie des Sciences, Isidore Geoffroy Saint-Hilaire proclame que « la découverte de la mâchoire fossile du singe de M. Lartet [...] parait appelée à commencer une ère nouvelle du savoir humanitaire »[2]. Lartet se permet d'affirmer que « l'existence paléontologique de l'homme est une supposition qui n'a rien d'invraisemblable », ouvrant ainsi la voie à l'acceptation de l'existence de l'homme fossile[3].
Origines
La problématique de l'origine des primates se doit de répondre à trois questions fondamentales :
- Qui étaient les premiers primates ?
- Quand sont-ils apparus ?
- Pourquoi ont-ils développé les caractéristiques qu'on leur connait aujourd'hui ?
Définition
L'ordre des primates a été créé par Carl von Linné en 1758 et pensé comme l'ordre « premier », celui qui domine tous les autres. Il comprenait alors quatre genres : Homo (l'Homme), Simia (les singes), Lemur (les prosimiens), et Vespertilio (les chauves-souris). Bien que les critères choisis par le père de la nomenclature zoologique aient été largement revus depuis, aucune autre classification convaincante n'a été adoptée, et il manque aux primates le trait distinctif fondamental que possèdent d'autres ordres de mammifères. Les spécialistes s'accordent néanmoins pour reconnaître une série de caractéristiques communes, notamment[4] :
- des orbites de grande taille orientées vers l’avant, permettant la vision binoculaire et stéréoscopique ;
- des pieds et des mains à cinq doigts, préhensiles et dotés, la plupart du temps, d'un pouce opposable ;
- des pieds et des mains terminés par des ongles plats, plutôt que des griffes, et possédant des coussinets tactiles ridés au bout des doigts (empreintes digitales) ;
- une réduction du museau et du sens de l’olfaction ;
- des portées peu nombreuses et une gestation, ainsi qu'un développement juvénile, plus longs que chez les mammifères de taille comparable ;
- un cerveau généralement plus développé et aux caractères anatomiques uniques.
Composantes
| Euarchontes | |||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
| Phylogénie reconstituée des euarchontes. D'après Bloch et al. (2007)[5]. |
En 1910, le paléontologue américain William King Gregory propose la création du super-ordre des archontes qui regroupe les primates au sens strict, les macroscélides (musaraignes à trompe), les scandentiens (toupayes), les dermoptères (lémurs volants) et les chiroptères (chauves-souris)[6]. Cette division a été plus ou moins soutenue pendant tout le XXe siècle, en excluant les macroscélides. Plusieurs auteurs ont placé les toupayes parmi les primates, dans un sous-ordre avec les plesiadapiformes, celui des Praesimii[7]. Les études moléculaires ont rapidement exclu les chiroptères, mais ont appuyé l'existence de ce groupe, rebaptisé « Euarchontes ».
D'après le registre fossile
L'hypothèse la plus ancienne fait du Cénozoïque « l'âge des mammifères », par opposition au Mésozoïque qui aurait été « l'âge des dinosaures ». Les mammifères placentaires auraient proliféré peu avant ou juste après la limite K/T (environ 65 millions d'années), au moment de la disparition de la plupart des autres formes de vie qui parcouraient la terre à ce moment-là (y compris les dinosaures). Le dépeuplement soudain de la planète aurait ouvert de nouvelles niches écologiques profitant à la radiation évolutive rapide de groupes comme les primates.
Cette théorie est étayée par le registre fossile des primates et de nombreux autres mammifères. En effet, aucune trace de la présence d'un euarchonte avant le début du Paléocène n'a été découverte à ce jour. Une étude basée sur les caractéristiques morphologiques de plusieurs fossiles récemment découverts[5] a soutenu l'hypothèse de l'apparition des primates (comprenant les plesiadapiformes) en Amérique du Nord à la fin du Crétacé et de la séparation des euprimates au Paléocène supérieur.
D'après l'horloge moléculaire

Les études basées sur l'hypothèse de l'horloge moléculaire concluent pour la plupart à une séparation plus ancienne, entre 80 et 116 millions d'années. Les primates constitueraient l'une des toutes premières lignées de mammifères placentaires à s'être différenciée et parmi eux la séparation entre haplorhiniens et strepsirhiniens serait survenue au Crétacé supérieur[8].
D'après la biogéographie
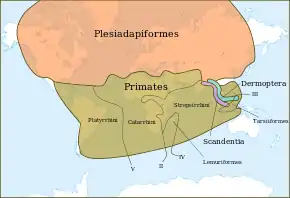
Si l'on se base non pas sur l'âge des fossiles les plus anciens mais sur les limites biogéographiques qui séparent les principaux clades, il est possible d'avancer l'apparition des premiers primates de plusieurs dizaines de millions d'années. Ainsi, en se basant sur les grands événements tectoniques, un ancêtre largement distribué se serait différencié en un groupe septentrional (Plesiadapiformes), un groupe méridional (Primates) et deux groupes orientaux (Dermoptères et Scandentiens), à la suite de l'éclatement de la Pangée au Jurassique inférieur, il y a 185 millions d'années[9].
Parmi les primates, la séparation entre strepsirhiniens et haplorhiniens serait due au volcanisme du monoclinal du Lebombo, dans le Sud-Est de l'Afrique, et se serait produite il y a 180 Ma. Au sein de ces deux groupes, des divergences auraient eu lieu ultérieurement lors de la formation du canal du Mozambique (Jurassique moyen, 160 ma) entre les lémuriens et les loris, et encore plus tard entre les singes de l'Ancien et ceux du Nouveau Monde, à la suite de l'ouverture de l'Atlantique Sud (Crétacé inférieur, 130 Ma)[9].
Ce modèle, bien que marginal au sein de la communauté scientifique, permet d'expliquer la distribution des platyrrhiniens en Amérique du Sud et des lémuriens à Madagascar. Les autres reconstruction requièrent un ou plusieurs événements de dispersion transocéanique au moyen de radeaux de végétation. Mais comment les primates auraient-ils pu traverser ainsi l'Atlantique et le canal du Mozambique, alors qu'ils n'ont, par exemple, jamais franchi le bras de mer de 20 km qui sépare les Célèbes de l'Australasie ?
Hypothèse « arboricole »
Selon cette première théorie, la plus ancienne[10], l'origine des primates représenterait une radiation évolutive à partir de mammifères arboricoles ancestraux. L'impulsion de ce virage évolutif serait le changement entre un régime alimentaire insectivore et un régime principalement herbivore. Les mains et les pieds de ces premiers primates auraient ainsi évolué pour faciliter la locomotion à travers les réseaux complexes de branches d'arbres.
Si cette théorie permet de supporter les adaptations au niveau de la dentition et de la morphologie des membres, elle présente plusieurs contradictions. De nombreux mammifères non-primates ont des modes de vie arboricole sans montrer les mêmes adaptations. Ainsi, les animaux au mode de locomotion similaire aux écureuils se déplacent dans les branches avec plus de stabilité. De même, les griffes semblent plus adaptées que les ongles pour monter et descendre le long de supports verticaux. Les autres animaux arboricoles montrent également une préférence marquée pour la communication olfactive, laquelle est particulièrement peu développée chez la plupart des primates. Enfin, pourquoi les autres mammifères au mode de vie similaire ne sont-ils pas dotés de la convergence des orbites en position faciale (qui permet la vision stéréoscopique), dont bénéficient les primates et certains oiseaux de proie comme les faucons ou les hiboux ?
Hypothèse de la « prédation visuelle »
Pour pallier les carences de la thèse arboricole, l'anthropologue américain Matt Cartmill[11] propose que c'est plutôt pour s'adapter à la capture de proies dans les arbres que les primates ont développé la plupart des caractéristiques qu'on leur connait. Ainsi, les mains et les pieds préhensiles et la réduction des griffes en ongles auraient été une adaptation pour la recherche nocturne d'insectes dans les branches terminales des arbres. La convergence orbitale aiderait à mesurer la distance avec la proie, sans avoir à bouger la tête (à la manière des hiboux), et la régression olfactive s'expliquerait justement par le rapprochement des parois des orbites oculaires.
Hypothèse du « régime alimentaire mixte »
La dernière théorie sur l'origine des primates, développée par l'anthropologue Robert Sussman[12], est basée sur la co-évolution avec les angiospermes. Elle s'appuie sur le développement plus ou moins parallèles des plantes à fleurs, qui auraient offert de nouvelles opportunités nutritives et une niche écologique alors inexploitée. Les premiers primates auraient été des omnivores capables de se nourrir de fruits, de fleurs, de gommes, de nectars et des insectes qui évoluaient sur ces objets situés aux extrémités des branches. La vision stéréoscopique aurait été développée pour distinguer les produits alimentaires à de faibles niveaux de luminosité et leur manipulation aurait nécessité une meilleure coordination œil-main.
Évolution
Comme le consensus scientifique n'est pas encore formé quant aux liens qui relient les différents fossiles aux espèces actuelles, il est usuel de présenter l'évolution des primates par époque géologique.
Paléocène
Durant le Paléocène, l'Amérique du Nord, l'Amérique du Sud, l'Afrique et l'Inde étaient des continents-îles. La majeure partie de l'Europe et du Moyen-Orient était submergée, et l'Australie venait à peine de se séparer de l'Antarctique. Le climat global était beaucoup plus chaud et plus humide qu'aujourd'hui et une grande part de l'hémisphère nord était recouverte de forêts tempérées. C'est de cette partie du monde que proviennent la plupart des primates fossiles de cette époque.
Purgatorius : un ancêtre contesté

Purgatorius est un genre dont on connait actuellement quatre espèces et qui a principalement été décrit à partir de fossiles du début du Paléocène. Sa réputation de « tout premier primate » provient d'une unique molaire découverte dans le Montana et datée du Crétacé supérieur, qui lui a été attribuée. Classé par la plupart des auteurs comme un représentant primitif de la famille des Paromomyidae[13], il serait donc un ancêtre des Plesiadapiformes plutôt que des Primates modernes. Il n'en reste pas moins le fossile connu le plus proche de l'ancêtre commun à ces deux groupes, et corroborerait la thèse de leur séparation avant le début du Cénozoïque.
Plésiadapides

Les plésiadapides forment un groupe très diversifié de mammifères arboricoles euraméricains dont l'appartenance ou non à l'ordre des primates est régulièrement débattue. D'une taille allant de 100 g à 5 kg, ces animaux étaient caractérisés par une museau assez allongé, de grandes incisives proéminentes, des orbites placés latéralement, et la présence de griffes plutôt que d'ongles au bout de leurs phalanges. Ils étaient vraisemblablement nocturnes, s'alimentaient de graines et d'insectes et se déplaçaient dans les arbres en trottinant, à la manière des écureuils. Ces traits primitifs les rendent assez éloignés morphologiquement des euprimates (ou « vrais primates ») de l’Éocène.
Pendant une grande partie du XXe siècle, les plésiadapides ont été considérés comme des primates, mais de nombreux auteurs ont depuis préféré les classer dans un ordre à part, les Plesiadapiformes. Néanmoins, une étude de 2007, incluant plusieurs fossiles du Paléocène récemment découverts, est revenue sur cette distinction et a affirmé qu'ils faisaient pleinement partie des primates[5].
Éocène
Au début de l'Éocène, le climat global était extrêmement chaud et humide. L'Amérique du Sud et l'Afrique étaient alors des continents séparés, mais il existait des connexions terrestres entre l'Asie et l'Europe, ainsi qu'entre cette dernière et l'Amérique du Nord. Par la suite, durant plusieurs millions d'années, cette dernière connexion fut perdue, ce qui a provoqué l'évolution différenciée des animaux sur ces continents. La plupart des mammifères de l'Éocène étaient de taille et de masse plus réduites que leurs équivalents actuels.
Adapides : les premiers strepsirrhiniens

Les adapides étaient de petits primates archaïques qui évoluaient en Amérique du Nord et en Europe durant tout l'Éocène, et dont on a décrit plus d'une centaine d'espèces. Leur masse corporelle variait entre 100 g et 6,9 kg, ce qui leur donnait une taille comparable aux Strepsirrhiniens actuels. Leurs orbites réduites indiquent un mode de vie vraisemblablement diurne et leur morphologie dentaire, caractérisée par de petites incisives et de grandes canines, dénote un régime alimentaire plutôt folivore et frugivore. Dotés d'un museau allongé, d'une longue queue et de longues pattes, les adapides avaient une apparence générale proche des lémuriens malgaches.
Malgré ces similitudes, les Adapiformes ne sont pas les ancêtres directs des Lemuriformes ni des Lorisiformes actuels, mais plutôt un groupe ayant évolué parallèlement. Ils sont constitués de trois familles, Sivaladapidae, Adapidae et Notharctidae (dont fait partie Darwinius masillae).
Omomyides : les premiers tarsiers ?

Les omomyides avaient une taille comprise entre 45 g et 2,5 kg, mais la plupart des espèces retrouvées sont en général plus petites que les adapides. Comme eux, ils évoluaient en Amérique du Nord et en Europe, ainsi qu'en Asie. Ils avaient le museau court et la taille de leurs orbites laisse présager un mode de vie nocturne. Leur régime alimentaire, extrapolé à partir de leur morphologie dentaire, semble avoir été de type insectivore pour les petites espèces, et de type frugivore et même folivore pour les grandes. Quelques espèces montrent une fusion entre le tibia et la fibula, comme c'est le cas chez les tarsiers actuels.
L'unique famille des Omomyidae comprend de nombreuses subdivisions et est souvent classée parmi les Tarsiiformes. Certains auteurs insistent sur les divergences avec ces derniers, et rangent les omomyides dans leur propre infra-ordre (Omomyiformes).
Les premiers singes
Les premiers singes connus, dotés de trois prémolaires, apparaissent en Asie de l'Est au milieu de l'Éocène, il y a environ 45 millions d'années. Ils appartiennent à deux familles archaïques : les Afrotarsiidae et les Eosimiidae (dont font partie Eosimias et Bahinia). On trouve ensuite en Asie du Sud-Est la famille des Pondaungidae.
Les singes du Fayoum
Les singes primitifs comptaient parmi les mammifères afro-eurasiatiques les plus courants à la fin de l'Éocène. La majeure partie de nos connaissances provient des fouilles effectuées dans la dépression du Fayoum, en Égypte. Cette région, à présent sèche et aride, était couverte de marais tropicaux il y a 35 Ma. Les taxons identifiés appartiennent notamment à trois groupes, dont les relations phylogénétiques ne sont pas encore totalement élucidées : Parapithecidae, Oligopithecidae, et Proteopithecidae[14].
Les Parapithecidae évoluaient à la fin de l’Éocène et au début de l'Oligocène. Leur taille variait de celle d'un ouistiti pour le genre Qatrania à celle d'un cercopithèque pour le genre Parapithecus. Leur formule dentaire était identique à celle des Platyrrhiniens actuels, ce qui laisse penser qu'il s'agit de la formule primitive des singes.
Ces différentes familles aujourd'hui éteintes précèdent la divergence entre Platyrrhiniens et Catarrhiniens. Bien qu'aucun fossile de ces deux micro-ordres antérieur à la Grande Coupure n'ait encore été trouvé, on pense que leur divergence s'est produite avant la fin de l'Éocène.
Oligocène
La Grande Coupure, il y a 34 millions d'années, est marquée par un refroidissement rapide du climat, qui entraine une extinction massive des espèces. De nombreux adapiformes disparaissent à cette époque. Durant cette césure, la plupart des primates semblent avoir disparu d'Amérique du Nord et d'Europe.
Durant l'Oligocène, il n'y avait pas de pont terrestre entre les deux Amériques. La séparation de l'Australie et de l'Amérique du Sud d'avec l'Antarctique créa un changement important des courants océaniques autour du pôle Sud, ce qui eut pour principales conséquences de maintenir la baisse des températures et du niveau des mers. La réduction consécutive des précipitations fit reculer les forêts, surtout dans l'hémisphère nord.

Les premiers Catarrhiniens
La dépression du Fayoum a aussi livré des fossiles de singes catarrhiniens primitifs, les Propliopithecidae, dont fait partie Aegyptopithecus zeuxis[14].
Le groupe des Pliopithecidae, qui lui succède, a connu un grand succès évolutif, au point de se répandre sur plusieurs continents. Le premier spécimen, Pliopithecus antiquus, a été identifié en 1837 à Sansan (Gers, France) par Édouard Lartet.
Les Platyrrhiniens
Tous les singes modernes sud-américains descendraient d’une souche unique, mais cet ancêtre commun n’a pu encore être identifié. Les plus vieux fossiles exhumés (espèces des genres Branisella et Dolichocebus) datent d’environ 25 millions d’années.
En débarquant il y a au moins 34 millions d’années dans le Nouveau Monde, les singes ont trouvé une terre d’accueil éminemment propice à leur épanouissement et à leur diversification. Les seuls mammifères occupant des niches écologiques voisines sont les paresseux, les opossums, les porcs-épics et les écureuils. De nos jours, 30 % des espèces de primates recensées vivent sur ce seul continent et d’autres restent à découvrir dans les régions inexplorées d’Amazonie.
L’Amérique du Sud eut une position insulaire durant presque toute l’ère tertiaire. Une faune unique s’y développa pendant plusieurs dizaines de millions d’années. À plusieurs reprises lors des périodes de sécheresse, le bloc de la forêt humide se fragmenta en poches géographiques, créant des biotopes séparés appelés « refuges », qui entrainèrent à leur tour une modification des préférences écologiques et un isolement reproductif, deux facteurs-clés dans le développement de nouvelles espèces. Durant le tertiaire, les seuls grands prédateurs d’Amérique du Sud étaient les borhyènes et Thylacosmilus, des marsupiaux carnivores, trop lents pour constituer un danger pour les primates.
Au début du Pléistocène, lorsqu’une connexion fut rétablie entre les deux continents au niveau de Panama, la faune nord-américaine déferla vers le sud où régnaient de meilleures conditions climatiques. Tous les marsupiaux (hormis les opossums) furent évincés par les nouveaux arrivants. Des carnivores plus agressifs firent leur apparition, comme le smilodon (« tigre » à dents de sabre) ou le cerdocyon (« renard » des prairies, omnivore). Pourtant, aucun des prédateurs venus du nord ne put mettre en danger la moindre espèce de primates. De nos jours, l’ocelot et le chat-tigre constituent de réelles menaces pour les petits singes, mais ces derniers ripostent par la vélocité, la vigilance et une communication évoluée.
Miocène
Le principal évènement tectonique du Miocène est la collision entre la plaque africaine et la plaque eurasiatique au niveau de la péninsule Arabique, qui est intervenue il y a 17 à 18 Ma. Le résultat fut la réduction de la Téthys et l'érection de chaines de montagnes dans les régions méditerranéennes, le Caucase et les monts Zagros.
Durant la première moitié du Miocène, le climat global devint plus chaud et plus humide, favorisant l'expansion des forêts dans le sud de l'Eurasie. Parmi les nombreux groupes d'espèces qui profitèrent de cet optimum climatique et de la nouvelle connexion entre Afrique et Eurasie figurent les hominoïdes. On peut ainsi diviser cette époque entre une première partie, où les hominoïdes fossiles ne sont connus qu'en Afrique, et une seconde partie, où ils sont également connus en Eurasie.
Les hominoïdes du Miocène inférieur

Le départ de la « lignée humaine »
C'est dans la seconde moitié du Miocène, entre -10 et -6 Ma, que s'effectue la séparation entre la lignée humaine et celle des chimpanzés. Le plus ancien fossile d'hominidé bipède, Sahelanthropus tchadensis, date de -7 Ma, mais on a trouvé aussi dans le sud de l'Europe et en Afrique vers -9 Ma Oreopithecus bambolii. Les australopithèques font leur apparition au Pliocène et les premiers humains (genre Homo), dès le début du Pléistocène. Cette lignée ne compte plus qu'une seule espèce vivante : l'Homme moderne, qui a conquis toute la surface du globe.
Pliocène
Durant le Pliocène, le niveau des mers s'abaissa et la température globale diminua par rapport à la fin du Miocène. Cette époque fut surtout marquée par la fermeture de l'isthme de Panama et le grand échange interaméricain qui s'ensuivit.
Les lémuriens subfossiles
À ce jour, aucun fossile de lémurien antérieur à l'Holocène n'a été clairement identifié. L'hypothèse la plus probable de leur origine à Madagascar est qu'ils auraient traversé le canal du Mozambique durant l’Éocène, emportés depuis l'Afrique par un radeau de végétation. L'île comprenait néanmoins de nombreux genres et espèces aujourd'hui éteints, dont certains étaient caractérisés par une taille largement supérieure aux formes actuelles. La plupart de ces extinctions se sont produites à la suite de l'arrivée des humains, il y a 2000 ans.
Notes et références
- (en) David R. Begun, « Planet of the Apes », dans Evolution : A Scientific American Reader, University of Chicago Press, (lire en ligne), p. 270
- Académie des Sciences de Paris, Comptes rendus hebdomadaires des séances de l'Académie des Sciences, vol. 5, Gauthier-Villars, (lire en ligne), p. 41.
- Francis Duranthon, Histoires de mammifères, Éditions Bréal, , p. 59
- Robert Boyd et Joan Silk, « Introduction aux primates », dans L'aventure humaine : Des molécules à la culture, De Boeck Supérieur, (lire en ligne), p. 125
- (en) Jonathan I. Bloch, Mary T. Silcox, Doug M. Boyer et Eric J. Sargis, « New Paleocene skeletons and the relationship of plesiadapiforms to crown-clade primates », Proceedings of the National Academy of Sciences of the United States of America, vol. 104, no 4, , p. 1159–1164 (lire en ligne).
- (en) William King Gregory, « The orders of mammals », Bulletin of the American Museum of Natural Society, no 27, , p. 1-524.
- (en) Philip D. Gingerich, « Primate evolution: evidence from the fossil record, comparative morphology, and molecular biology », Yearbook of Physical Anthropology, no 27, , p. 57-72 (lire en ligne).
- (en) Mark S. Springer, William J. Murphy, Eduardo Eizirik et Stephen J. O'Brien, « Placental mammal diversification and the Cretaceous–Tertiary boundary », Proceedings of the National Academy of Sciences of the United States of America, vol. 100, no 3, , p. 1056-1061 (lire en ligne)
- (en) Michael Heads, « Evolution and biogeography of primates : a new model based on molecular phylogenetics, vicariance and plate tectonics », Zoologica Scripta, vol. 39, no 2, , p. 107–127 (lire en ligne).
- (en) F. S. Szalay, « Paleobiology of the earliest primates », dans R. Tuttle, The Functional and Evolutionary Biology of Primates, Chicago, Aldine-Atherton, (lire en ligne), p. 3-35.
- (en) Matt Cartmill, « New views on primate origins », Evolutionary Anthropology, vol. 1, no 3, , p. 105-111.
- (en) Robert W. Sussman, « Primate origins and the evolution of angiosperms », American Journal of Primatology, vol. 23, no 4, , p. 209-223.
- (en) William A. Clemens, « Purgatorius, an early paromomyid primate (Mammalia) », Science, vol. 184, no 4139, , p. 903-905 (résumé)
- (en) Erik R. Seiffert, Elwyn L. Simons, John G. Fleagle et Marc Godinot, « Paleogene Anthropoids », dans Cenozoic Mammals of Africa, University of California Press, .
Bibliographie
- (en) Walter Carl Hartwig, The Fossil Primate Record, Cambridge University Press, , 530 p. (lire en ligne).
- (en) Matthew J. Ravosa et Marian Dagosto, Primate Origins : Adaptations and Evolution, Springer Science & Business Media, , 864 p. (lire en ligne).
- (en) John G. Fleagle, Primate Adaptation and Evolution, Academic Press, , 464 p. (lire en ligne).
Voir aussi
Articles connexes
- Histoire évolutive des mammifères
- Primates (classification phylogénétique)
- Liste des primates fossiles
 Portail origine et évolution du vivant
Portail origine et évolution du vivant  Portail des primates
Portail des primates