Modèle de Hodgkin–Huxley
Le modèle Hodgkin-Huxley est un modèle mathématique qui décrit comment les potentiels d'action des neurones sont initiés et propagés. Il s'agit d'un ensemble d'équations différentielles non linéaires qui se rapprochent des caractéristiques électriques de cellules excitables telles que les neurones ou les myocytes cardiaques. C'est un système dynamique en temps continu.
Originellement le modèle fut décrit en 1952 par Alan Hodgkin et Andrew Huxley pour expliquer les mécanismes ioniques sous-tendant l'initiation et la propagation des potentiels d'action observé dans l'axone géant du calmar[1]. Le prix Nobel de physiologie ou médecine de 1963 leur fut attribué pour ce travail.
Alors que les expériences originales ne traitaient que des canaux sodium et potassium, le modèle Hodgkin-Huxley peut également être étendu pour tenir compte d'autres espèces de canaux ioniques.
Composants de base
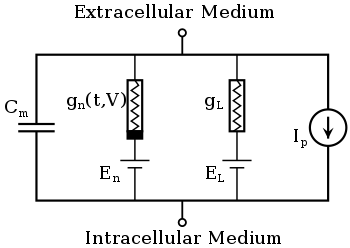
Le modèle de Hodgkin-Huxley traite chaque composant d'une cellule excitable comme un élément électrique (comme le montre la figure). La bicouche lipidique est représentée par une capacité . Les canaux ioniques voltage-dépendants sont représentés par les conductances , soumisent aux gradients électrochimiques entraînant le flux d'ions , (où n est le canal ionique spécifique) fonction de la tension et du temps. Les canaux de fuite sont représentés par des conductances linéaires . Les valeurs de dont les valeurs sont déterminées par le rapport entre les concentrations intra- et extracellulaires des espèces ioniques d'intérêt. Enfin, les pompes à ions sont représentées par des sources de courant (). Le potentiel membranaire est noté .






Ainsi, le courant traversant la bicouche lipidique est donné par:

et le courant traversant un canal ionique est donné par :

où est le potentiel d'inversion du i-ième canal ionique. Ainsi, pour une cellule possédant des canaux sodium et potassium, le courant total traversant la membrane est donné par :

- ,

où I est le courant membranaire total par unité de surface, est la capacité membranaire par unité de surface, et sont les conductances de potassium et de sodium par unité de surface, respectivement, et sont les potentiels d'inversion du potassium et du sodium, respectivement, et et sont respectivement la conductance de fuite par unité de surface et le potentiel d'inversion de fuite. Les éléments dépendant du temps de cette équation sont , et , où les deux dernières conductances dépendent également explicitement de la tension.







Caractérisation du courant ionique
Dans les canaux ioniques voltage-dépendants, la conductance du canal est fonction à la fois du temps et de la tension ( dans la figure), tandis que dans les canaux de fuite est une constante ( dans la figure). Le courant généré par les pompes ioniques dépend des espèces ioniques spécifiques à cette pompe. Les sections suivantes décriront ces formulations plus en détail.

Canaux ioniques voltage-dépendants
À l'aide d'une série d'expériences de voltage imposé et en faisant varier les concentrations extracellulaires de sodium et de potassium, Hodgkin et Huxley ont développé un modèle dans lequel les propriétés d'une cellule excitable sont décrites par un ensemble de quatre équations différentielles ordinaires[1]. Ce modèle est développé originellement pour l'axones de calmar qui ne contiennent que des canaux de sodium et potassium mais peut être étendu à d'autres axones possédant plus de canaux. Ainsi en plus l'équation du courant total donné ci-dessus, on a aussi :
- ,




où I est le courant par unité de surface, et et sont des constantes de vitesse pour le i- ième canal ionique, qui dépendent de la tension mais pas du temps. est la valeur maximale de la conductance. n, m et h sont des quantités sans dimension entre 0 et 1 qui sont respectivement associées à l'activation des canaux potassiques, à l'activation des canaux sodiques et à l'inactivation des canaux sodiques. Pour , et prendre la forme :






- ,

- .

et sont les valeurs d'état stable pour l'activation et l'inactivation, respectivement, et sont généralement représentées par les équations de Boltzmann en tant que fonctions de . Dans l'article original de Hodgkin et Huxley[1], les fonctions et sont donnés par :





où désigne la dépolarisation négative en mV.

Alors que dans de nombreux logiciels actuels[2], les modèles de type Hodgkin-Huxley généralisent et à :

Afin de caractériser les canaux voltage-dépendants, les équations sont ajustées aux données obtenu sous une tension d'entrée constante. Pour une dérivation des équations de Hodgkin-Huxley sous tension d'entrée constante, voir[3]. En bref, pour un potentiel membranaire maintenu à une valeur constante les équations de déclenchement non linéaires se réduisent aux équations de la forme :
- ,

- ,

- .

Ainsi, pour chaque valeur de potentiel de membrane les courants de sodium et de potassium peuvent être décrits par :
- .

Afin d'arriver à la solution complète pour un potentiel d'action propagé, on écrit le courant I du côté gauche de la première équation différentielle en fonction de V, de sorte que l'équation soit pour la tension seule. La relation entre I et V peut être dérivée de la théorie des câbles et est donnée par :
- ,

où a est le rayon de l'axone, R est la résistance spécifique de l'axoplasme et x est la position le long de la fibre nerveuse.
L'algorithme de Levenberg-Marquardt est souvent utilisé pour ajuster ces équations aux données de voltage-clamp[4].
Canaux de fuite
Les canaux de fuite tiennent compte de la perméabilité naturelle de la membrane aux ions et prennent la forme de l'équation des canaux voltage-dépendants, où la conductance est une constante. Ainsi, le courant de fuite dû aux canaux ioniques de fuite passifs dans le formalisme de Hodgkin-Huxley est .


Pompes et échangeurs
Le potentiel membranaire dépend du maintien des gradients de concentration ionique à travers elle. Le maintien de ces gradients de concentration nécessite un transport actif d'espèces ioniques. Les échangeurs sodium-potassium et sodium-calcium sont les plus connus d'entre eux. Certaines des propriétés de base de l'échangeur Na/Ca sont déjà bien établies : la stoechiométrie d'échange est de 3 Na + : 1 Ca 2+ et l'échangeur est électrogène et sensible à la tension. L'échangeur Na/K a également été décrit en détail, avec une stoechiométrie 3 Na + : 2 K + [5],[6].
Propriétés mathématiques
Le modèle Hodgkin-Huxley peut être considéré comme un système d'équations différentielles avec quatre variables d'état, , et , qui changent par rapport au temps . Le système est difficile à étudier car il s'agit d'un système non linéaire et ne peut pas être résolu analytiquement. Cependant, il existe de nombreuses méthodes numériques disponibles pour analyser le système. Certaines propriétés et comportements généraux, tels que les cycles limites, peuvent être prouvés.



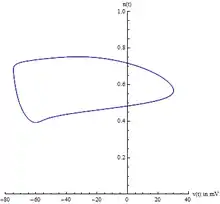
Collecteur central
Comme il existe quatre variables d'état, il peut être difficile de visualiser le chemin dans l'espace des phases. Habituellement, deux variables sont choisies, la tension et la variable de contrôle du potassium , permettant de visualiser le cycle limite. Cependant, il faut être prudent car il s'agit d'une méthode ad hoc de visualisation du système à 4 dimensions. Cela ne prouve pas l'existence du cycle limite.


Une meilleure projection peut être construite à partir d'une analyse minutieuse du Jacobien du système, évalué au point d'équilibre. Plus précisément, les valeurs propres du Jacobien sont indicatives de l'existence de la variété centrale. De même, les vecteurs propres du Jacobien révèlent l'orientation de la variété centrale. Le modèle Hodgkin-Huxley a deux valeurs propres négatives et deux valeurs propres complexes avec des parties réelles légèrement positives. Les vecteurs propres associés aux deux valeurs propres négatives se réduiront à zéro à mesure que le temps t augmente. Les deux vecteurs propres complexes restants définissent la variété centrale. En d'autres termes, le système à 4 dimensions s'effondre sur un plan à 2 dimensions. Toute solution partant du collecteur central se désintégrera vers le collecteur central. De plus, le cycle limite est contenu sur le collecteur central.
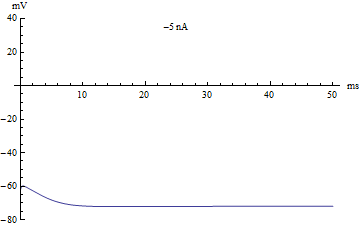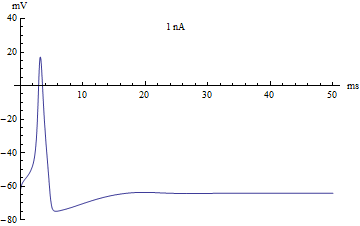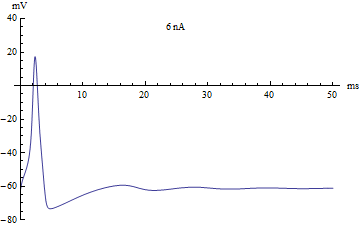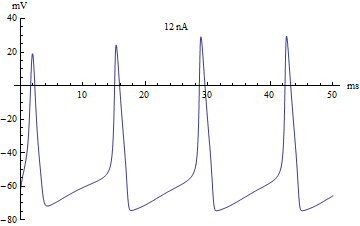
Bifurcations
Si le courant injecté ont été utilisés comme paramètre de bifurcation, alors le modèle Hodgkin-Huxley subit une bifurcation de Hopf. Comme avec la plupart des modèles neuronaux, l'augmentation du courant injecté augmentera le taux de décharge du neurone. Une conséquence de la bifurcation de Hopf est qu'il y a une cadence de tir minimale. Cela signifie que soit le neurone ne se déclenche pas du tout (correspondant à une fréquence nulle), soit se déclenche à la vitesse de déclenchement minimale. En raison du principe tout ou rien, il n'y a pas d'augmentation régulière de l'amplitude du potentiel d'action, mais plutôt un "saut" soudain d'amplitude. La transition qui en résulte est connue sous le nom de canard.

Améliorations et modèles alternatifs
Le modèle Hodgkin-Huxley est considéré comme l'une des grandes réalisations de la biophysique du XXe siècle. Néanmoins, les modèles modernes de type Hodgkin-Huxley ont été étendus de plusieurs manières importantes :
- Des populations de canaux ioniques supplémentaires ont été incorporées sur la base de données expérimentales.
- Le modèle Hodgkin-Huxley a été modifié pour incorporer la théorie des états de transition et produire des modèles thermodynamiques Hodgkin-Huxley[7].
- Les modèles incorporent souvent des géométries très complexes de dendrites et d'axones, souvent basées sur des données de microscopie.
- Modèles stochastiques du comportement des canaux ioniques, conduisant à des systèmes hybrides stochastiques[8].
- Le modèle Poisson-Nernst-Planck (PNP) est basé sur une approximation du champ moyen des interactions ioniques et des descriptions de continuum de concentration et de potentiel électrostatique[9].
Plusieurs modèles neuronaux simplifiés ont également été développés (comme le modèle FitzHugh-Nagumo), facilitant une simulation efficace à grande échelle de groupes de neurones, ainsi qu'un aperçu mathématique de la dynamique de génération de potentiel d'action.
Notes et références
- « A quantitative description of membrane current and its application to conduction and excitation in nerve », The Journal of Physiology, vol. 117, no 4, , p. 500–44 (PMID 12991237, PMCID 1392413, DOI 10.1113/jphysiol.1952.sp004764).
- Nelson ME (2005) Electrophysiological Models In: Databasing the Brain: From Data to Knowledge. (S. Koslow and S. Subramaniam, eds.) Wiley, New York, pp. 285–301.
- Daniel Johnston Gray et Samuel Miao-Sin Wu, Foundations of cellular neurophysiology, Cambridge, Massachusetts [u.a.], 3rd, (ISBN 978-0-262-10053-3).
- Vladimir F. Krapivin, Costas A. Varotsos et Vladimir Yu. Soldatov, New Ecoinformatics Tools in Environmental Science : Applications and Decision-making, Springer, , 37–38 p. (ISBN 9783319139784, lire en ligne).
- « Stoichiometry and voltage dependence of the sodium pump in voltage-clamped, internally dialyzed squid giant axon », The Journal of General Physiology, vol. 93, no 5, , p. 903–41 (PMID 2544655, PMCID 2216238, DOI 10.1085/jgp.93.5.903).
- Bertil Hille, Ion channels of excitable membranes, Sunderland, Massachusetts, 3rd, (ISBN 978-0-87893-321-1).
- Forrest, « Can the Thermodynamic Hodgkin–Huxley Model of Voltage-Dependent Conductance Extrapolate for Temperature? », Computation, vol. 2, no 2, , p. 47–60 (DOI 10.3390/computation2020047, lire en ligne
 ).
). - Pakdaman, Thieullen et Wainrib, « Fluid limit theorems for stochastic hybrid systems with applications to neuron models », Adv. Appl. Probab., vol. 42, no 3, , p. 761–794 (DOI 10.1239/aap/1282924062, Bibcode 2010arXiv1001.2474P, arXiv 1001.2474).
- Zheng et Wei, « Poisson-Boltzmann-Nernst-Planck model », Journal of Chemical Physics, vol. 134, no 19, , p. 194101 (PMID 21599038, PMCID 3122111, DOI 10.1063/1.3581031, Bibcode 2011JChPh.134s4101Z).
Voir aussi
Articles connexes
Bibliographie
- A. L. Hodgkin et A. F. Huxley, « Currents carried by sodium and potassium ions through the membrane of the giant axon of Loligo », The Journal of Physiology, vol. 116, no 4, , p. 449–472 (ISSN 0022-3751, PMID 14946713, PMCID 1392213, lire en ligne, consulté le ).
- A. L. Hodgkin et A. F. Huxley, « The components of membrane conductance in the giant axon of Loligo », The Journal of Physiology, vol. 116, no 4, , p. 473–496 (ISSN 0022-3751, PMID 14946714, PMCID 1392209, lire en ligne, consulté le ).
- A. L. Hodgkin et A. F. Huxley, « The components of membrane conductance in the giant axon of Loligo », The Journal of Physiology, vol. 116, no 4, , p. 473–496 (ISSN 0022-3751, PMID 14946714, PMCID 1392209, lire en ligne, consulté le ).
- A. L. Hodgkin et A. F. Huxley, « A quantitative description of membrane current and its application to conduction and excitation in nerve », The Journal of Physiology, vol. 117, no 4, , p. 500–544 (ISSN 0022-3751, PMID 12991237, PMCID 1392413, lire en ligne, consulté le ).
- A. L. Hodgkin, A. F. Huxley et B. Katz, « Measurement of current-voltage relations in the membrane of the giant axon of Loligo », The Journal of Physiology, vol. 116, no 4, , p. 424–448 (ISSN 0022-3751, PMID 14946712, PMCID 1392219, lire en ligne, consulté le ).
Liens externes
- Simulation Javascript interactive du modèle HH — Fonctionne dans n'importe quel navigateur compatible HTML5. Permet de modifier les paramètres du modèle et l'injection de courant.
- Applet Java interactif du modèle HH — Les paramètres du modèle peuvent être modifiés ainsi que les paramètres d'excitation et les tracés de l'espace des phases de toutes les variables sont possibles.
- Lien direct vers le modèle Hodgkin-Huxley et une description dans la base de données BioModels.
- Impulsions neuronales : le potentiel d'action en action par Garrett Neske, The Wolfram Demonstrations Project.
- Modèle interactif Hodgkin-Huxley par Shimon Marom, The Wolfram Demonstrations Project.
- ModelDB — Une base de données de code source en neurosciences computationnelles contenant 4 versions (dans différents simulateurs) du modèle Hodgkin-Huxley original et des centaines de modèles qui appliquent le modèle Hodgkin-Huxley à d'autres canaux dans de nombreux types de cellules électriquement excitables.
- Plusieurs articles sur la version stochastique du modèle et son lien avec l'original.
 Portail des neurosciences
Portail des neurosciences