Temnospondyli
Temnospondyles

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Super-classe | Tetrapoda |
| Classe | Amphibia |
Taxons de rang inférieur
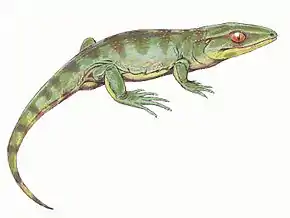
Les temnospondyles (Temnospondyli), forment un ordre très diversifié de tétrapodes ayant prospéré dans le monde entier durant le Carbonifère, le Permien et le Trias, apparaissant rarement dans les registres fossiles du Jurassique et du Crétacé. Au cours d'environ 210 millions d'années d'histoire évolutive, ils s'adaptent à un large éventail d'habitats, y compris les environnements d'eau douce, terrestres et même marins côtiers. Leur histoire de vie est bien comprise, avec des fossiles connus depuis le stade larvaire, la métamorphose et la maturité. La plupart des temnospondyles auraient été semi-aquatiques, même si certains seraient presque entièrement terrestres, ne retournant à l'eau que pour se reproduire. Ces temnospondyles figurent parmi les premiers vertébrés pleinement adaptés à la vie terrestre. Bien que les temnospondyles soient considérés comme des amphibiens, beaucoup ont des caractéristiques inhabituelles telles que des écailles, des griffes et des plaques osseuses en forme d'armure, qui les distinguent des amphibiens modernes.
Les temnospondyles sont connus depuis le début du XIXe siècle et étaient initialement considérés comme des reptiles. Ils seront par la suite décrits à plusieurs reprises comme des batraciens, des stégocéphales et des labyrinthodontes, bien que ces noms soient maintenant rarement utilisés. Les animaux désormais regroupés dans les Temnospondyli sont répartis entre plusieurs groupes d'amphibiens jusqu'au début du XXe siècle, date à laquelle ils s'avèrent appartenir à un taxon distinct en fonction de la structure de leurs vertèbres. Le nom du groupe vient du grec et provient des mots τέμνειν / temnein, « couper » et σπόνδυλος / spondulos « vertèbre », en raison du fait que chaque vertèbre est divisée en plusieurs parties.
Les paléontologues ne sont pas d'accord sur la question de savoir si les temnospondyles seraient les ancêtres des amphibiens modernes (anoures, salamandres et cécilies) ou si l'ensemble du groupe s'est éteint sans laisser la moindre descendance. Différentes hypothèses placent les amphibiens actuels comme les descendants des temnospondyles, ou d'un autre groupe de tétrapodes primitifs appelés lépospondyles, voire comme étant les descendants des deux groupes (les céciliens descendant des lépospondyles et les batraciens des temnospondyles). Des études récentes placent une famille de temnospondyles appelée Amphibamidae comme les parents les plus proches des amphibiens modernes. Des similitudes dans les dents, les crânes et les structures auditives relient les deux groupes.
Description



Si beaucoup de temnospondyles présentaient des tailles comparables à nos amphibiens actuels et ressemblaient à des salamandres, nombre d'entre eux étaient beaucoup plus grands et ressemblaient à première vue à des crocodiles, qui occupent aujourd'hui les mêmes niches écologiques[1]. La plupart avaient de larges têtes plates, parfois courtes (brévirostres), parfois allongées (longirostres). Vus de dessus, leurs crânes étaient arrondis ou triangulaires, et généralement couverts de creux et de crêtes.
De nombreux temnospondyles avaient sur la tête des sillons appelés sensoriels, généralement situés autour des narines et des orbites, et faisant partie d'un système de sillons très probablement utilisés pour détecter les vibrations dans l'eau[1]. Comme les animaux semi-aquatiques, la plupart des temnospondyles avaient de petits membres, avec quatre orteils palmés aux pattes avant et cinq aux pattes arrière. Les espèces terrestres avaient, en plus, des membres puissants et certains avaient même des griffes[2].
Un temnospondyle, Fayella, avait la particularité d'avoir des membres relativement longs par rapport à son corps, ce qui laisse penser qu'il était probablement un prédateur gracile particulièrement actif, peut-être coureur et/ou arboricole. Ce nouveau genre a été attribué à la famille des Dissorophidae et au clade des Olsoniformes[3],[4].
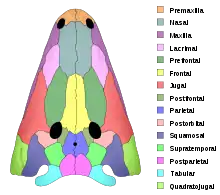
La forme des os de temnospondyles se retrouve chez d'autres tétrapodes anciens, même si chez certains taxons sont apparus des os crâniens supplémentaires, interfrontaux, internasaux et interpariétaux[1]. La plupart avaient des « cornes » à l'arrière du crâne : excroissances osseuses arrondies, séparées par des encoches appelées échancrures otiques. Chez certains temnospondyles comme Zatrachys, elles sont pointues et très saillantes. Les temnospondyles ont souvent des dents sur le palais aussi bien que sur les mâchoires. Certaines de ces dents sont si grandes qu'elles sont appelées défenses. Chez certains temnospondyles comme Nigerpeton, des défenses de la mâchoire inférieure percent le palais et sortent par des ouvertures sur le haut du crâne[5].
Un des traits les plus marquants des temnospondyles est la présence d'une vacuité interptérygoidienne, avec deux gros trous à l'arrière de la bouche. Deux autres trous appelés choanes sont présents en face de ces vacuités mais on ne sait pas s'il y avait une communication fonctionnelle entre la cavité nasale et la bouche, ou si ces trous étaient tendus d'une membrane souple. Dans les deux cas on ne peut qu'imaginer leur fonction : gustative ? olfactive ? aide à la déglutition ? équilibrage de la pression dans l'eau ? émission de "chants" ? tout cela à la fois ? La réponse ne pourra venir que de la découverte de tissus mous fossilisés. On n'en connaît que très peu. Un bloc de grès décrit en 2007, trouvé dans la Formation de Mauch Chunk en Pennsylvanie (datant du début du Carbonifère), présente des empreintes cutanées de trois temnospondyles. Ces empreintes montrent qu'ils avaient la peau lisse, des membres robustes avec des pieds palmés et une crête de peau sur leur face inférieure[6].
Des empreintes de pas attribuées à de petits temnospondyles ont également été découvertes dans des roches du Carbonifère et du Permien. Ces empreintes sont habituellement trouvées dans des couches sédimentaires autour des points d'eau, ce qui, comme les formes larvaires aquatiques, montre leurs liens avec l'eau[7].
.JPG.webp)
À la différence des amphibiens modernes, la plupart des temnospondyles étaient couverts de petites écailles serrées, voire de grandes plaques dermiques dans la région ventrale. Pendant les premiers stades de leur développement, ils avaient simplement de petites écailles arrondies. Lorsque les animaux grandissaient, les écailles du dessous du corps se développaient en plaques ventrales. C'est pour cela que, par le passé, on les a pris pour des reptiles. Les plaques se chevauchaient de façon à permettre un large éventail de mouvements. Il semble que, par la suite, les temnospondyles semi-aquatiques comme les trématosaures et les capitosaures aient perdu leurs écailles, permettant probablement des mouvements plus souples dans l'eau ou une respiration cutanée[8].
Plusieurs groupes de temnospondyles présentent de grandes plaques osseuses sur le dos. Ceux du genre Peltobatrachus avaient des plaques qui couvraient aussi bien leur dos que leur ventre[9]. Le genre Laidleria avait aussi un vaste plaquage sur son dos. La plupart des membres de la famille des Dissorophidae avaient une protection dorsale, même si elle ne couvrait que la ligne médiane du dos, grâce à deux rangées de plaques étroites[10]. D'autres temnospondyles, comme ceux du genre Eryops avaient de petites plaques osseuses en forme de disques, qui étaient probablement intégrées dans leur peau. Tous ces temnospondyles étaient adaptés à un mode de vie terrestre. Leur armure peut leur avoir offert une protection contre la dessication, ainsi que contre les prédateurs dans le cas de Peltobatrachus[9]. Les écailles peuvent aussi avoir assuré la stabilité de la colonne vertébrale, en limitant sa flexibilité grâce à de puissants ligaments qui reliaient les écailles entre elles[11].
Les temnospondyles tels que Sclerothorax et Eryops, qui peuvent avoir été au moins partiellement terrestres, avaient aussi de longs processus épineux au-dessus de leurs vertèbres, qui pouvaient stabiliser leur colonne vertébrale[12]. Des plaques osseuses ont également été observées chez des plagiosaures, mais contrairement aux Peltobatrachus, Laidleria, Eryops et les dissorophidés, ces animaux semblent avoir été totalement aquatiques. Les plagiosaures peuvent avoir hérité leur armure d'un ancêtre terrestre, car tant Peltobatrachus que Laidleria sont considérés comme proches du groupe[9].
Les temnospondyles doivent leur nom à leurs vertèbres en plusieurs segments. Chez les tétrapodes actuels, le corps principal de la vertèbre est en un seul morceau appelé le centrum, mais chez les temnospondyles il est partagé en un pleurocentrum et un intercentrum. On distingue deux types de vertèbres chez les temnospondyles : les rhachitomes et les stéréospondyles.
- Dans les rhachitomes, l’intercentrum est grand et en forme de coin, et le pleurocentrum est formé de blocs relativement petits qui s’emboîtent entre eux. Les deux éléments soutiennent l'épine neurale, et des excroissances osseuses bien développées appelées zygapophyses qui viennent renforcer les liens entre les vertèbres. Ce type d'épine dorsale solide et fort rencontré chez plusieurs temnospondyles leur a permis d'être partiellement et, dans certains cas totalement, terrestres.
- Pour les stéréospondyles, le pleurocentrum a entièrement disparu, l’intercentrum élargi devenant la partie principale des vertèbres. Ce type d'ossature fragile indique que les temnospondyles à stéréospondyles passaient plus de temps dans l'eau[13].
Histoire de leur étude
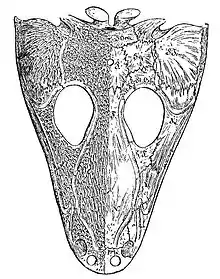
Le nom de Temnospondyli a été créé par le paléontologue allemand Karl Alfred von Zittel dans sa seconde édition du Handbuch der Palaeontologie, publié en 1888. Des restes de temnospondyles étaient déjà connus depuis le début du XIXe siècle. Le premier à être décrit a été le fameux Mastodonsaurus, nommé par Georg Friedrich Jaeger en 1828, et dont un exemplaire reconstitué trône à la Haus der Natur de Salzbourg (Autriche). Jaeger avait nommé Mastodonsaurus une seule dent qu'il a estimé être celle d'un reptile. Mastodonsaurus signifie « lézard à dents en mamelon » d'après la forme de la pointe de la dent[14].
La dénomination des premiers spécimens a cependant été contestée. Léopold Fitzinger a renommé l'animal Batrachosaurus en 1837. En 1841, le paléontologue anglais Richard Owen a proposé le nom Labyrinthodon pour décrire ses dents très pliées ou labyrinthiques. Owen pensait que le nom Mastodonsaurus « ne devait pas être conservé, car il rappelle inévitablement l'idée de mastodonte (un genre de mammifères), ou bien une forme de dent en mamelon… et parce que le deuxième élément du nom saurus, indique une fausse parenté, l'animal n'étant pas un saurien, mais un batracien. »[15]. Owen avait reconnu que l'animal n'était pas un reptile, aussi il le rapprocha du genre Phytosaurus de Jaeger. Bien que les deux aient été de taille similaire avec des dents coniques, Phytosaurus s'est révélé plus tard être un reptile ressemblant à un crocodile. Des éléments supplémentaires, notamment le crâne, ont solidement placé le genre Labyrinthodon parmi les amphibiens. Jaeger avait également créé l'espèce Salamandroides giganteus en 1828, en se basant sur une portion d'occiput. En 1833, il a décrit un crâne complet de S. giganteus qui avait les mêmes dents que son Mastodonsaurus : c'était en fait le premier crâne complet d'un temnospondyle. Comme le nom Mastodonsaurus avait été attribué en premier, il a eu priorité sur les autres noms, qui sont devenus des synonymes[16]. Batrachosaurus est encore utilisé comme nom d'un temnospondyle de la famille des brachyopidés.
Mastodonsaurus et d'autres animaux similaires ont été placés dans les labyrinthodontes en raison de leurs dents fortement repliées en coupe transversale. Labyrinthodon jaegeri a été trouvé plus tard par le paléontologue William Buckland à Guys Cliffe, en Angleterre. D'autres spécimens ont été trouvés dans les grès rouges du Warwickshire. Comme de nombreux fossiles ont été découverts en Angleterre, Owen décrivit ces labyrinthodontes comme les « plus évolués des batraciens » et les a comparés aux crocodiles, qu'il considérait comme la forme la plus évoluée des reptiles. Il a également noté que ces grands labyrinthodontes du Keupérien (un étage géologique de la fin du Trias) étaient plus récents que les reptiles les plus évolués ayant vécu au Zechstein, un sous-étage du Permien supérieur. Owen a utilisé ces fossiles pour contrer l'idée que les reptiles avaient évolué à partir des premiers amphibiens (qu'il appelait «poissons métamorphosés »)[17].
En plus de Mastodonsaurus, parmi les premiers genres de Temnospondyles figurent Metopias et Rhombopholis en 1842, Zygosaurus en 1848, Trematosaurus en 1849, Baphetes et Dendrerpeton en 1853, Capitosaurus en 1858 et Dasyceps en 1859[18]. Baphetes est maintenant cconsidéré comme un tétrapode primitif placé en dehors des Temnospondyli et Rhombopholis comme un reptile prolacertiforme[19],[20].
Plus tard au xixe siècle, les temnospondyles ont été classés dans les Stegocephalia, un nom inventé par le paléontologue américain Edward Drinker Cope en 1868. Cope avait placé les stégocéphaliens dans la classe des batraciens (nom remplacé ultérieurement par celui d'amphibiens). Stegocephalia signifie en grec « tête en toit », une référence à la large tête plate de temnospondyles et autres tétrapodes primitifs. À cette époque, les paléontologues considéraient les temnospondyles comme des amphibiens, car ils en possédaient trois caractéristiques principales : les arcs branchiaux retrouvés dans les squelettes des juvéniles, indiquant qu'ils étaient amphibies au moins dans la première partie de leur vie ; les côtes qui ne se connectent pas à la face inférieure de la cage thoracique ; et des fosses profondes au niveau du crâne interprétées comme un espace pour les glandes à mucus[21].
Plusieurs sous-ordres de stégocephaliens ont été créés à la fin du xixe et du début du xxe siècle. Des animaux aujourd'hui considérés comme des temnospondyles étaient considérés au départ des labyrinthodontes, mais certains ont été classés dans les Branchiosauria. Les branchiosaures étaient de petits animaux avec des dents coniques simples, tandis que les labyrinthodontes étaient plus grands et avaient des dents plus complexes faites de dentine et d'émail. Les branchiosaures ne comprenaient que quelques genres tels que le Branchiosaurus d'Europe et l’Amphibamus d'Amérique du Nord, qui avait un squelette réduit, des branchies externes et pas de côtes. Quelques squelettes d’Amphibamus ont été retrouvés avec des côtes normales, ce qui incite à les réaffecter aux Microsauria (mais des études plus détaillées ont trouvé que c'était un temnospondyle)[22]. Des tissus mous comme des écailles et des branchies externes ont été trouvés sur de nombreux fossiles bien conservés de branchiosaures en Allemagne. Au début du xxe siècle, on s'est rendu compte que les branchiosaures étaient en fait des formes larvaires de temnospondyles auxquels il manquait de nombreuses fonctionnalités typiques qui définissent le groupe.

D'autres animaux plus tard classés dans les temnospondyles ont été placés dans un groupe appelé Ganocephala, caractérisé par des os du crâne en plateau, de petits membres, des écailles et des arcs branchiaux. Contrairement aux labyrinthodontes, ils n'ont pas de foramen pariétal, un petit trou dans le crâne en arrière des orbites. Archegosaurus, Dendrerpeton, Eryops et Trimerorhachis ont été placés dans ce groupe et ont été considérés comme les membres les plus primitifs des Reptilia. Leurs vertèbres rhachitomes, leur notochorde et le manque de condyles occipitaux (qui joignent la tête au cou) sont des caractéristiques qu'ils partagent avec les poissons. Aussi, ils ont été considérés comme un lien entre les premiers poissons et les formes plus avancées comme les stégocephaliens[23].
La structure générale, le crâne, et notamment les dents pédicellées bicuspides ressemblent fortement à ceux des anoures et salamandres actuels.
Classification
Classifications standard
À l'origine, les temnospondyles étaient classés selon la structure de leurs vertèbres. Les premières formes, disposants des vertèbres complexes composées d'un certain nombre d'éléments séparés, ont été placées dans le sous-ordre des Rachitomi (en), et les grandes formes aquatiques du Trias avec des vertèbres plus simples ont été placées dans le sous-ordre des Stereospondyli. Avec l'essor récent de la phylogénétique, cette classification n'est plus considéré comme viable. La condition rhachitomique de base se trouve dans de nombreux tétrapodes primitifs et n'est pas unique aux groupe des temnospondyles. De plus, la distinction entre les vertèbres rhachitomeuses et stéréospondyles n'est pas tout à fait claire. Certains temnospondyles ont des vertèbres rhachitomes, semi-rhachitomes et stérospondyles en différents points d'une même colonne vertébrale. D'autres taxons ont des morphologies intermédiaires qui ne rentrent dans aucune catégorie. Rachitomi n'est plus reconnu en tant que groupe, mais Stereospondyli est toujours considéré comme valide[24],[25]. Ci-dessous, une taxonomie simplifiée des temnospondyles montrant les groupes actuellement reconnus :



- Ordre Temnospondyli
- Super-famille Edopoidea
- Clade Eutemnospondyli
- Famille Dendrerpetidae
- Clade Rhachitomi
- Sous-ordre Dvinosauria
- Famille Trimerorhachidae (en)
- Super-famille Dvinosauroidea
- Super-famille Dissorophoidea
- Famille Micromelerpetontidae (en)
- Clade Xerodromes (en)
- Clade Amphibamiformes (en)
- Family Amphibamidae
- Family Branchiosauridae (en)
- Family Micropholidae (en)
- Clade Olsoniformes (en)
- Famille Dissorophidae
- Famille Trematopidae
- Clade Amphibamiformes (en)
- Super-famille Eryopoidea (en)
- Famille Eryopidae (en)
- Famille Zatracheidae
- Sous-ordre Dvinosauria
- Clade Limnarchia
- Clade Stereospondylomorpha
- Famille Archegosauridae
- Famille Intasuchidae (en) (placement incertain)
- Famille Sclerocephalidae (en)
- Sous-ordre Stereospondyli
- Famille Peltobatrachidae (en)
- Famille Lapillopsidae
- Famille Rhinesuchidae
- Famille Rhytidosteidae
- Famille Lydekkerinidae
- Clade Neostereospondyli
- Clade Capitosauria
- Super-famille Mastodonsauroidea (Capitosauroidea)
- Infra-ordre Trematosauria (en)
- Super-famille Trematosauroidea
- Famille Metoposauridae
- Famille Trematosauridae
- Super-famille Trematosauroidea
- Super-famille Brachyopoidea
- Famille Brachyopidae (en)
- Famille Chigutisauridae (en)
- Famille Plagiosauridae
- Clade Capitosauria
- Clade Stereospondylomorpha
Phylogénie
Dans l'une des premières analyses phylogénétiques du groupe, Gardiner (1983) a reconnu cinq caractéristiques qui faisaient de Temnospondyli un clade : un os situé à l'arrière du crâne, le parasphénoïde, qui est relié à un autre os situé sous le crâne, le ptérygoïde, de grandes ouvertures appelées vides interptérygoïdes sont présentes entre les ptérygoïdes, l'étrier (un os impliqué dans l'ouïe) est relié au parasphénoïde et se projette vers le haut, le cleithrum (un os de la ceinture pectorale) est mince, et une partie de la vertèbre appelée interdorsale s'attache à l'arc neural[26]. Des caractéristiques supplémentaires ont été données par Godfrey et al. en 1987, y compris le contact entre le postpariétal (en) et l'os occipital à l'arrière du crâne, de petites saillies, appelées processus unciné, sur les côtes et une ceinture pelvienne avec chaque côté ayant une seule lame iliaque[27]. Ces caractéristiques communes sont appelées synapomorphies.
Les temnospondyles sont généralement placés comme des tétrapodes basaux dans les analyses phylogénétiques, leur positionnement exact variant d'une étude à l'autre[28]. Selon la classification des amphibiens modernes, ils sont soit inclus dans le groupe-couronne Tetrapoda ou au plus grand clade Stegocephalia. Les représentants du groupe-couronne des tétrapodes sont les descendants de l'ancêtre commun le plus récent de tous les tétrapodes vivants et les proto-tétrapodes sont des formes qui ne font pas partie du groupe-couronne. Les amphibiens modernes ont récemment été suggérés comme étants les descendants des temnospondyles, ce qui les placerait dans le groupe-couronne Tetrapoda. Ci-dessous, un cladogramme de Ruta et al. (2003) plaçant Temnospondyli dans les Tetrapoda[29] :
| ◄ Stegocephalia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
D'autres études placent les amphibiens modernes comme les descendants des lépospondyles et placent les temnospondyles dans une position plus basale dans la tige des Tetrapoda. Ci-dessous, un cladogramme de Laurin et Reisz (1999) plaçant Temnospondyli à l'extérieur du groupe-couronne Tetrapoda[30].
| ◄ Stegocephalia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
La plupart des analyses phylogénétiques sur les interrelations des temnospondyles se concentrent sur les familles individuelles. L'une des premières études à grande échelle sur la phylogénie des temnospondyles a été menée par le paléontologue Andrew Milner en 1990[31]. Une étude de 2007 a créé un « super-arbre » de toutes les familles de temnospondyles, combinant les arbres au niveau familial des précédentes études. Le cladogramme suivant est modifié par Ruta et al. (2007)[32] :
|
|
1 Temnospondyli, 2 Edopoidea, 3 Dvinosauria, 4 Euskelia, 5 Eryopoidea (en), 6 Dissorophoidea, 7 Limnarchia, 8 Archegosauroidea (en), 9 Stereospondyli, 10 Rhytidostea (en), 11 Brachyopoidea, 12 Capitosauria, 13 Trematosauria (en), 14 Metoposauroidea (en)
Le groupe le plus basal de temnospondyles sont la super-famille Edopoidea. Les édopoïdes ont plusieurs caractéristiques primitives ou plésiomorphes, dont un seul condyle occipital et un os appelé l'intertemporal qui est absent chez les autres temnospondyles. Les édopoïdes comprennent le genre Edops datant du Carbonifère supérieur et la famille des Cochleosauridae (en). Les Dendrerpetontidae ont également été inclus dans les Edopoidea et constituent la plus ancienne famille de temnospondyles connue. Balanerpeton woodi est la plus ancienne espèce identifié à ce jour, ayant été présente il y a plus de 330 millions d'années au stade Viséen du Carbonifère inférieur. Des analyses récentes placent les Dendrerpetontidae en dehors des Edopoidea dans une position plus dérivée[33],[34]. D'autres temnospondyles primitifs incluent Capetus et Iberospondylus (en). Saharastega et Nigerpeton, tous deux décrits en 2005 au Niger, sont également primitifs mais sont datées du Permien supérieur. Ils sont près de 40 millions d'années plus jeunes que les autres temnospondyles basaux, ce qui implique l'existence une longue lignée fantôme (en) d'espèces qui ne sont pas encore connues dans les archives fossiles[35].
En 2000, les paléontologues Adam Yates et Anne Warren ont éffectué une phylogénie révisée de temnospondyles plus dérivés, nommant ainsi plusieurs nouveaux clades. Deux clades principaux sont Euskelia et Limnarchia. Euskelia comprend les temnospondyles qui étaient autrefois appelés Rhachitomi et comprend deux sous-familles, les Dissorophoidea et les Eryopoidea (en). Les dissorophoïdes comprennent de petits temnospondyles principalement terrestres qui peuvent être les ancêtres des amphibiens modernes. Les éryopoïdes comprennent des temnospondyles plus grands comme Eryops. Le deuxième clade majeur, Limnarchia, comprend la plupart des temnospondyles du Mésozoïque, ainsi que certains groupes du Permien. Au sein de Limnarchia se trouvent la super-famille des Archegosauroidea (en) et les temnospondyles les plus dérivés, les stéréospondyles[25].
Yates et Warren ont également nommé le taxon Dvinosauria, un clade de petits temnospondyles aquatiques du Carbonifère, du Permien et du Trias[25]. Ces derniers placent Dvinosauria dans Limnarchia, mais des études plus récentes sont en désaccord sur leur position. Par exemple, une étude de 2007 les place comme étant encore plus basaux que les euskéliens, tandis qu'une étude de 2008 les maintient en tant que limnarchiens basaux[32],[36].
Au sein des Stereospondyli, Yates et Warren ont érigé deux clades majeurs : Capitosauria et Trematosauria. Les capitosaures comprennent de grands temnospondyles semi-aquatiques comme Mastodonsaurus avec des têtes plates et des yeux situé près de l'arrière du crâne. Les trématosaures comprennent une diversité de temnospondyles, y compris de grands trématosauroïdes marins, des plagiosaures (en) aquatiques, des brachyopoïdes qui ont survécu jusqu'au Crétacé et des métoposauroïdes (en) avec des yeux près de l'avant de la tête. En 2000, les paléontologues Rainer Schoch et Andrew Milner ont nommé un troisième clade majeur de stéréospondyles, les Rhytidostea (en)[37]. Ce groupe comprenait des stéréospondyles plus primitifs qui ne pouvaient être placés ni dans Capitosauria ni dans Trematosauria, et comprenait des groupes comme Lydekkerinidae, Rhytidosteidae et Brachyopoidea. Alors que Capitosauria et Trematosauria sont encore largement utilisés, Rhytidostea n'est pas souvent pris en charge comme un véritable clade dans les analyses récentes. Les rhytidostéidés et les brachyopoïdes sont maintenant regroupés avec les trématosauriens, mais les lydekkérinidés sont toujours considérés comme une famille primitive de stéréospondyles[38],[39].
Une nouvelle phylogénie des temnospondyles a été proposée par le paléontologue Rainer Schoch en 2013. Elle soutient de nombreux clades trouvés par Yates et Warren, mais ne trouve pas de support pour leur division des stéréospondyles dérivés en Euskelia et Limnarchia. Les éryopidés se sont avérés plus étroitement liés aux stéréospondyles qu'aux dissorophoïdes, qui ont été regroupés avec les dvinosaures. Le clade comprenant les Eryopidae et les Stereospondylomorpha a été nommé Eryopiformes (en). De plus, Schoch nomme le clade contenant tous les temnospondyles à l'exception des édopoïdes sous le terme Eutemnospondyli et rétablit le nom Rhachitomi pour le clade contenant tous les temnospondyles à l'exception des édopoïdes et des dendrerpetontidés. Ci-dessous le cladogramme de l'analyse de Schoch (2013)[40] :
| ◄ Temnospondyli |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Liens de parenté avec les amphibiens actuels

Les amphibiens modernes (regroupant les grenouilles, salamandres et cécilies) sont classés dans les Lissamphibia, un taxon dont les premiers représentants semblent être apparus durant le Permien. Les estimations de l'horloge moléculaire placent le premiers lissamphibiens au Carbonifère supérieur, mais les chercheurs estiment que le premier membre des Batrachia, le taxon regroupant les grenouilles et salamandres, seraient apparu au Permien moyen en utilisant la même technique[41],[42].
En utilisant les preuves fossiles, il existe trois théories principales sur l'origine des amphibiens modernes. La première est qu'ils ont évolué à partir de temnospondyles du groupe des dissorophoïdes[43],[44]. La seconde est qu'ils ont évolué à partir des lépospondyles, très probablement des lysorophiens[45]. La troisième hypothèse est que les céciliens descendent des lépospondyles mais que les grenouilles ainsi que les salamandres ont évolué à partir des dissorophoïdes[46].
Récemment, la théorie selon laquelle les temnospondyles seraient les ancêtres de tous les lissamphibiens a reçu un large soutien. La morphologie du crâne de certains petits temnospondyles a été comparée à celle des grenouilles et des salamandres modernes, mais la présence de dents bicuspides et pédicellées (en) chez les petits temnospondyles pédomorphes ou immatures est cité comme l'argument le plus convaincant en faveur de l'origine temnospondyle des lissamphibiens[47]. Vu chez les lissamphibiens et de nombreux temnospondyles dissorophoïdes, les dents pédicellées ont des pointes et des bases calcifiées. Au cours du développement de la plupart des tétrapodes, les dents commencent à se calcifier à leur extrémité. La calcification se poursuit normalement vers le bas jusqu'à la base de la dent, mais la calcification à partir de la pointe s'arrête brusquement dans les dents pédicellées. La calcification reprend à la base, laissant une zone au centre de la dent non calcifiée, un trait qui est observable chez les amphibiens préhistorique et actuels[48].
Les chercheurs pensent que la famille de dissorophoïde des Amphibamidae est la plus étroitement liée aux Lissamphibia. En 2008, un amphibamidé appelé Gerobatrachus hottoni a été découvert au Texas et est nommé ainsi de par sa tête semblable à celui d'une grenouille et son corps en forme de salamandre. Ce genre était autrefois considéré comme le temnospondyle le plus étroitement apparenté aux lissamphibiens et a été placé comme taxon frère du groupe dans une analyse phylogénétique. Les paléontologues pensent maintenant qu'une autre espèce d'amphibamidé, appelée Doleserpeton annectens (en), est encore plus étroitement liée aux lissamphibiens. Contrairement à Gerobatrachus, Doleserpeton était connu depuis 1969, et la présence de dents pédicellées dans ses mâchoires a conduit certains paléontologues à en conclure peu de temps après son appellation qu'il s'agissait d'un parent des amphibiens modernes. Il a d'abord été décrit comme un « proto-lissamphibien », et l'épithète spécifique annectens, signifiant « connecter », est nommé en référence à sa position transitionnelle déduite entre les temnospondyles et les lissamphibiens[47]. La structure du tympanum (en), une membrane en forme de disque qui fonctionne comme un tympan, est similaire à celle des grenouilles et a également été utilisée comme preuve d'une relation phylogénétique étroite[49],[50]. D'autres caractéristiques inclut la forme du palais et de l'arrière du crâne, les côtes courtes et la surface lisse du crâne indiquent également qu'il s'agit d'un parent plus proche des lissamphibiens que Gerobatrachus. Ci-dessous, un cladogramme modifié de Sigurdsen et Bolt (2010) montrant les relations entre Gerobatrachus, Doleserpeton et Lissamphibia[51] :
| ◄ Temnospondyli |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Chinlestegophis (en), un stéréospondyle putatif du Trias considéré comme apparenté aux Metoposauroidea (en) tels que Rileymillerus (en), partage de nombreuses caractéristiques avec les céciliens, un groupe vivant d'amphibiens fouisseurs sans pattes. Si Chinlestegophis est en effet à la fois un stéréospondyle dérivé et un parent des céciliens, cela signifierait que bien que tous les lissamphibiens descendent des temnospondyles, les différents groupes seraient descendus de différentes branches de l'arbre généalogique des temnospondyles. Les anoures et les urodèles seraient donc des dissorophoïdes tandis que les céciliens seraient les uniques représentants actuels des stéréospondyles[52].
Notes et références
Notes
Références
- (en) J.-S. Steyer, and Laurin, M., « Temnospondyli », Tree of Life Web Project, (consulté le ).
- (en) A.P. Hunt, « A new amphibamid (Amphibia: Temnospondyli) from the Late Pennsylvanian (Middle Stephanian) of central New Mexico, USA », Paläontologische Zeitschrift, vol. 70, nos 3-4, , p. 555-565 (DOI 10.1007/BF02988092).
- (en) Bryan M. Gee, Diane Scott, Robert R. Reisz Reappraisal of the Permian dissorophid Fayella chickashaensis, Revue canadienne des sciences de la Terre, 2018, 55(10): 1103-1114, https://doi.org/10.1139/cjes-2018-0053
- (en) E.C. Olson, « Fayella chickashaensis, the Dissorophoidea and the Permian terrestrial radiations », Journal of Paleontology, vol. 46, no 1, , p. 104-114.
- (en) J.S. Steyer, « The vertebrate fauna of the Upper Permian of Niger. IV. Nigerpeton ricqlesi (Temnospondyli: Cochleosauridae), and the edopoid colonization of Gondwana », Journal of Vertebrate Paleontology, vol. 26, no 1, , :18–28 (DOI 10.1671/0272-4634(2006)26[18:TVFOTU]2.0.CO;2).
- (en) C. Stratton, « Ancient Amphibians Left Full-Body Imprints », GSA Newsroom, The Geological Society of America, (consulté le ).
- (en) A.P. Hunt, and Lucas, S.G., Pennsylvanian Footprints in the Black Warrior Basin of Alabama, vol. 1, Alabama Paleontological Society, (lire en ligne), « Tetrapod ichnofacies and their utility in the Paleozoic », p. 113-119.
- (en) F. Witzmann, « The evolution of the scalation pattern in temnospondyl amphibians », Zoological Journal of the Linnean Society, vol. 150, no 4, , p. 815–834 (DOI 10.1111/j.1096-3642.2007.00309.x).
- (en) A.L. Panchen, « A new armoured amphibian from the Upper Permian of East Africa », Philosophical Transactions of the Royal Society of London B, vol. 242, no 691, , p. 207-281 (DOI 10.1098/rstb.1959.0005).
- (en) J.R. Bolt, « Armor of dissorophids (Amphibia: Labyrinthodontia): an examination of its taxonomic use and report of a new occurrence », Journal of Paleontology, vol. 48, no 1, , p. 135-14.
- (en) D.W. Dilkes, « Comparison and biomechanical interpretations of the vertebrae and osteoderms of Cacops aspidephorus and Dissorophus multicinctus (Temnospondyli, Dissorophidae) », Journal of Vertebrate Paleontology, vol. 29, no 4, , p. 1013-1021 (DOI 10.1671/039.029.0410).
- (en) R.R. Schoch, « Anatomy and relationships of the Triassic temnospondyl Sclerothorax », Acta Palaeontologica Polonica, vol. 52, no 1, , p. 117–136.
- (en) E.H. Colbert, Evolution of the Vertebrates, New York, John Wiley & Sons, , 2e éd..
- G.F. Jaeger, Über die fossile reptilien, welche in Würtemberg aufgefunden worden sind, Stuttgart, J.B. Metzler, (lire en ligne), « Reptilien aus dem Alaunschiefer », p. 34-38
- (en) W. Jardine, « Proceedings of Learned Societies: Geological Society », The Annals and Magazine of Natural History, vol. 8, no 48, , p. 58-61
- (en) M. Moser, « Revision of the type material and nomenclature of Mastodonsaurus giganteus (Jaeger) (Temnospondyli) from the Middle Triassic of Germany », Palaeontology, vol. 50, no 5, , p. 1245–1266 (DOI 10.1111/j.1475-4983.2007.00705.x)
- (en) R. Owen, « Report on British fossil reptiles », Report of the Eleventh Meeting of the British Association for the Advancement of Science, vol. 11, , p. 60-204
- (en) R. Owen, Palaeontology or A systematic summary of extinct animals and their geological relations, Édimbourg, Adam and Charles Black, (lire en ligne), « Order II: Labyrinthodontia », p. 206-218
- (en) A.C. Milner, « Postcranial remains of Baphetes and their bearing on the relationships of the Baphetidae (= Loxommatidae) », Zoological Journal of the Linnean Society, vol. 22, no 1, , p. 211–235 (DOI 10.1111/j.1096-3642.1998.tb02530.x)
- (en) M.J. Benton, « Rhombopholis, a prolacertiform reptile from the Middle Triassic of England », Palaeontology, vol. 39, no 3, , p. 763-782
- (en) A.S. Woodward, Outlines of vertebrate palaeontology for students of zoology, Cambridge, University Press, , « Class Batrachia », p. 470
- (en) R.J. Moodie, « A contribution to a monograph of the extinct amphibia of North America. New forms from the Carboniferous », The Journal of Geology, vol. 17, no 1, , p. 38-82
- (en) R. Owen, Systematic summary of extinct animals and their geological relations, Édimbourg, Adam and Charles Black, (lire en ligne), « Order I: Ganocephala », p. 168-183
- (en) M. Laurin et Steyer, J.-S., « Phylogeny and Apomorphies of Temnospondyls », sur Tree of Life Web Project, (consulté le )
- (en) A.M. Yates et Warren, A.A., « The phylogeny of the 'higher' temnospondyls (Vertebrata: Choanata) and its implications for the monophyly and origins of the Stereospondyli », Zoological Journal of the Linnean Society, vol. 128, no 1, , p. 77–121 (DOI 10.1111/j.1096-3642.2000.tb00650.x
 )
) - (en) B.G. Gardiner, « Gnathostome vertebrae and the classification of the Amphibia », Zoological Journal of the Linnean Society, vol. 79, no 1, , p. 1–59 (DOI 10.1111/j.1096-3642.1983.tb01160.x)
- (en) S.J. Godfrey, Fiorillo, A.R. et Carroll, R.L., « A newly discovered skull of the temnospondyl amphibian Dendrerpeton acadianum Owen », Canadian Journal of Earth Sciences, vol. 24, no 4, , p. 796–805 (DOI 10.1139/e87-077, Bibcode 1987CaJES..24..796G)
- (en) Marcello Ruta, Jonathan E. Jeffery et Michael I. Coates, « A supertree of early tetrapods », Proceedings of the Royal Society of London B: Biological Sciences, vol. 270, no 1532, , p. 2507–2516 (ISSN 0962-8452, PMID 14667343, PMCID 1691537, DOI 10.1098/rspb.2003.2524, S2CID 22867564, lire en ligne [PDF])
- (en) M. Ruta, Coates, M.I. et Quicke, D.L.J., « Early tetrapod relationships revisited », Biological Reviews, vol. 78, no 2, , p. 251–345 (PMID 12803423, DOI 10.1017/S1464793102006103, S2CID 31298396, lire en ligne [PDF])
- (en) M. Laurin et Reisz, R.R., « A new study of Solenodonsaurus janenschi, and a reconsideration of amniote origins and stegocephalian evolution », Canadian Journal of Earth Sciences, vol. 36, no 8, , p. 1239–1255 (DOI 10.1139/e99-036, Bibcode 1999CaJES..36.1239L, lire en ligne [PDF])
- (en) A.R. Milner et Larwood, G.P., Major Evolutionary Radiations, Oxford, Clarendon Press, , 321–349 p., « The radiations of temnospondyl amphibians »
- (en) M. Ruta, Pisani, D., Lloyd, G. T. et Benton, M. J., « A supertree of Temnospondyli: cladogenetic patterns in the most species-rich group of early tetrapods », Proceedings of the Royal Society B: Biological Sciences, vol. 274, no 1629, , p. 3087–3095 (PMID 17925278, PMCID 2293949, DOI 10.1098/rspb.2007.1250, lire en ligne [PDF])
- (en) A.R. Milner, « The temnospondyl amphibian Dendrerpeton from the Upper Carboniferous of Ireland », Palaeontology, vol. 23, no 1, , p. 125–141 (lire en ligne [archive du ] [PDF])
- (en) R.B. Holmes, Carroll, R.L. et Reisz, R.R., « The first articulated skeleton of Dendrerpeton acadianum (Temnospondyli: Dendrerpentonidae) from the Lower Pennsylvanian locality of Joggins, Nova Scotia, and a review of its relationships », Journal of Vertebrate Paleontology, vol. 18, no 1, , p. 64–79 (DOI 10.1080/02724634.1998.10011034)
- C.A. Sidor, O'Keefe, F.R., Damiani, R.J., Steyer, J.-S., Smith, R.M.H., Larsson, H.C.E., Sereno, P.C., Ide, O. et Maga, A., « Permian tetrapods from the Sahara show climate-controlled endemism in Pangaea », Nature, vol. 434, no 7035, , p. 886–889 (PMID 15829962, DOI 10.1038/nature03393, Bibcode 2005Natur.434..886S, S2CID 4416647, lire en ligne [archive du ] [PDF], consulté le )
- (en) J. Englehorn, Small, B.J et Huttenlocker, A., « A redescription of Acroplous vorax (Temnospondyli: Dvinosauria) based on new specimens from the Early Permian of Nebraska and Kansas, U.S.A », Journal of Vertebrate Paleontology, vol. 28, no 2, , p. 291–305 (DOI 10.1671/0272-4634(2008)28[291:AROAVT]2.0.CO;2)
- (de) Rainer R. Schoch et Andrew R. Milner, Handbuch der Paläoherpetologie Part 3B. Stereospondyli, Munich, Verlag Dr. Friedrich Pfeil, , 203 p. (ISBN 978-3-931516-77-2)
- (en) A. Warren et Marsicano, C., « A phylogeny of the Brachyopoidea », Journal of Vertebrate Paleontology, vol. 20, no 3, , p. 462–483 (DOI 10.1671/0272-4634(2000)020[0462:APOTBT]2.0.CO;2)
- (en) A.M. Yates, « A new tiny rhytidosteid (Temnospondyli: Stereospondyli) from the Early Triassic of Australia and the possibility of hidden temnospondyl diversity », Journal of Vertebrate Paleontology, vol. 20, no 3, , p. 484–489 (DOI 10.1671/0272-4634(2000)020[0484:ANTRTS]2.0.CO;2)
- (en) R. R. Schoch, « The evolution of major temnospondyl clades: An inclusive phylogenetic analysis », Journal of Systematic Palaeontology, vol. 11, no 6, , p. 673–705 (DOI 10.1080/14772019.2012.699006, S2CID 83906628)
- (en) P. Zhang, Zhou, H., Chen, Y.-Q., Liu, L.-F. et Qu, L.-H., « Mitogenomic perspectives on the origin and phylogeny of living amphibians », Systematic Biology, vol. 54, no 3, , p. 391–400 (PMID 16012106, DOI 10.1080/10635150590945278 , lire en ligne [PDF])
- (en) D. San Mauro, Gower, D.J., Oommen, O.V., Wilkinson, M. et Zardoya, R., « Phylogeny of caecilian amphibians (Gymnophiona) based on complete mitochondrial genomes and nuclear RAG1 », Molecular Phylogenetics and Evolution, vol. 33, no 2, , p. 413–427 (PMID 15336675, DOI 10.1016/j.ympev.2004.05.014, lire en ligne [PDF])
- (en) Michael Benton, Vertebrate Palaeontology, Wiley, (ISBN 978-1-118-40764-6, lire en ligne), p. 398
- (en) Laurie J. Vitt et Janalee P. Caldwell, Herpetology: An Introductory Biology of Amphibians and Reptiles, Academic Press, (ISBN 978-0-12-386920-3, lire en ligne), p. 84
- (fr + en) Michel Laurin, « L'importance de la parcimonie globale et du biais historique pour la compréhension de l'évolution des tétrapodes. Partie I. Systématique, évolution de l'oreille moyenne et suspension mandibulaire », Annales des Sciences Naturelles - Zoologie et Biologie Animale, Paris, vol. 19, no 1, , p. 1–42 (DOI 10.1016/S0003-4339(98)80132-9)
- (en) J.S. Anderson, Reisz, R.R., Scott, D., Fröbisch, N.B. et Sumida, S.S., « A stem batrachian from the Early Permian of Texas and the origin of frogs and salamanders », Nature, vol. 453, no 7194, , p. 515–518 (PMID 18497824, DOI 10.1038/nature06865, Bibcode 2008Natur.453..515A, S2CID 205212809, lire en ligne [archive du ] [PDF])
- (en) J.R. Bolt, « Lissamphibian origins: possible protolissamphibian from the Lower Permian of Oklahoma », Science, vol. 166, no 3907, , p. 888–891 (PMID 17815754, DOI 10.1126/science.166.3907.888, Bibcode 1969Sci...166..888B, S2CID 10813454)
- (en) A.B. Vasil'eva et Smirnov, S.V., « Pedicellate teeth and the problems of amphibian phylogeny », Doklady Biological Sciences, vol. 376, no 5, , p. 89–90 (DOI 10.1023/A:1018858917237, S2CID 19553896)
- (en) J.R. Bolt et Lombard, R.E., « Evolution of the amphibian tympanic ear and the origin of frogs », Biological Journal of the Linnean Society, vol. 24, no 1, , p. 83–99 (DOI 10.1111/j.1095-8312.1985.tb00162.x)
- (en) T. Sigurdsen, « The otic region of Doleserpeton (Temnospondyli) and its implications for the evolutionary origin of frogs », Zoological Journal of the Linnean Society, vol. 154, no 4, , p. 738–751 (DOI 10.1111/j.1096-3642.2008.00459.x )
- (en) T. Sigurdsen et Bolt, J.R., « The Lower Permian amphibamid Doleserpeton (Temnospondyli: Dissorophoidea), the interrelationships of amphibamids, and the origin of modern amphibians », Journal of Vertebrate Paleontology, vol. 30, no 5, , p. 1360–1377 (DOI 10.1080/02724634.2010.501445, S2CID 85677757)
- (en) Jason D. Pardo, Bryan J. Small et Adam K. Huttenlocker, « Stem caecilian from the Triassic of Colorado sheds light on the origins of Lissamphibia », Proceedings of the National Academy of Sciences, vol. 114, no 27, , E5389–E5395 (ISSN 0027-8424, PMID 28630337, PMCID 5502650, DOI 10.1073/pnas.1706752114 )
Voir aussi
Articles connexes
Liens externes
- (en) Temnospondyli sur Tree of Life Web Project
- (en) Temnospondyli sur Palaeos (en)
- Ressources relatives au vivant :
- (cs + en) BioLib
- (en) Paleobiology Database
- (en) Interim Register of Marine and Nonmarine Genera
- (en + en) New Zealand Organisms Register
- Notices dans des dictionnaires ou encyclopédies généralistes :
 Portail de la paléontologie
Portail de la paléontologie  Portail de l’herpétologie
Portail de l’herpétologie