Rhinesuchidae
Rhinésuchidés
| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Amphibia |
| Ordre | † Temnospondyli |
| Sous-ordre | † Stereospondyli |
Genres de rang inférieur
- † Australerpeton ? Barbarena, 1998
- † Broomistega Shishkin & Rubidge, 2000
- † Laccosaurus Haughton, 1925
- † Rastosuchus Dias et al., 2020
- † Rhineceps Haughton, 1927
- † Rhinesuchoides Olson & Broom, 1937
- † Rhinesuchus (type) Broom, 1908
- † Uranocentrodon van Hoepen, 1917
- Australerpetonidae Barbarena, 1998
- Rhinecepidae Oschev, 1966
- Uranocentrodontidae Romer, 1947
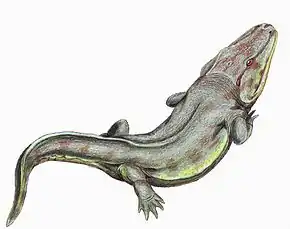
Les rhinésuchidés (Rhinesuchidae) forment une famille éteinte de grands tétrapodes ayant principalement vécu durant le Permien. Ils appartiennent au vaste groupe des temnospondyles, un taxon très diversifié de tétrapodes semi-aquatiques dont les amphibiens modernes seraient probablement les descendants. Les rhinésuchidés peuvent être différenciés des autres temnospondyles par certains détails de leurs crânes, notamment la structure intérieure de leurs échancrures otiques situées à l'arrière de ces derniers. Ils font partie des premiers membres divergents des stéréospondyles, un sous-groupe de temnospondyles ayant des têtes plates et des habitudes aquatiques. Bien que des stéréospondyles plus avancés aient évolué pour atteindre une répartition mondiale durant le Trias, les rhinesuchidés ont vécu principalement dans les environnements de haute latitude du Gondwana, constitué de ce qui est actuellement l'Amérique du Sud et l'Afrique, pendant le milieu et la fin du Permien.
La taxonomie de cette famille est complexe, avec plus de vingt espèces ayant été nommées par le passé, une revue publié en 2017 ne reconnaissant que huit d'entre eux et répartis en sept genres considérés comme valides. Alors que plusieurs membres présumés de ce groupe auraient vécu durant le Trias, la plupart sont soit douteux, soit n'appartiennent pas au groupe. Cependant, au moins un genre valide de rhinésuchidé est daté du début du Trias, et concerne un petit membre connu sous le nom de Broomistega. La définition formelle la plus récente des Rhinesuchidae, préconisée par Claudia A. Marsicano et al. en 2017, est celle d'un clade contenant tous les taxons plus étroitement liés à Rhinesuchus whaitsi qu'à Lydekkerina huxleyi ou Peltobatrachus pustulatus[1]. Une définition alternative similaire proposée par Rainer R. Schoch en 2013 est que Rhinesuchidae est un clade contenant tous les taxons plus étroitement liés à Uranocentrodon senekalensis qu'à Lydekkerina huxleyi, Trematosaurus brauni ou Mastodonsaurus giganteus[2].
Description
Morphologie
Les rhinésuchidés ont généralement un type de corps standard chez les tétrapodes, ces derniers disposant de quatre membres et d'une queue modérément longue. De plus, leurs corps sont généralement allongés et leurs membres sont petits et peu robustes, mais encore assez bien développés. Certains représentants qui sont classés dans cette famille sont très grands comparés aux amphibiens actuels, des genres comme Uranocentrodon mesurant jusqu'à une taille estimée à 3,75 mètres de longueur[N 1],[4]. Comme la plupart des stéréospondyles, leurs crânes sont aplatis et triangulaires, avec des yeux pointant vers le haut. La plupart des rhinésuchidés ont un museau relativement court, bien que celui d'Australerpeton soit très long et fin. Les seuls autres temnospondyles de grande taille du Permien à long museau furent les membres de la famille des archégosauridés, tels que Prionosuchus ou Konzhukovia[1],[5].
Échancrures otiques
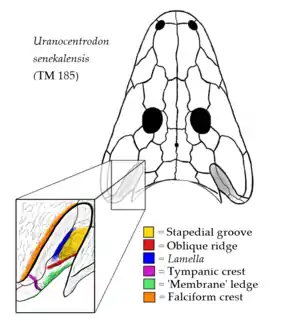
- Rainure stapédienne ;
- Crête oblique ;
- « Lamelle » (désignation basée selon l'étude publiée par Marsicano et al. en 2017) ;
- Crête tympanique ;
- Rebord « membranaire » ;
- Crête falciforme.
Comme la plupart des anciens amphibiens, les rhinésuchidés ont une paire d'indentations sur le bord arrière du crâne appelées échancrures otiques. Bien que parfois considérées comme ayant abrité des organes auditifs tels qu'un tympan, ces encoches sont plus susceptibles d'avoir contenu des spiracles, des trous charnus utilisés pour la respiration. Les rhinesuchidés peuvent être caractérisés par un système unique de crêtes et de rainures situé dans la cavité interne de chaque échancrure otique. Les parois de la cavité de l'échancrure otique, parfois appelée cavité tympanique, sont principalement constituées de la branche ascendante des os ptérygoïdes. Néanmoins, le bord intérieur de chaque cavité est formé par un os tabulaire. Les os tabulaires sont une paire d'os de formes triangulaires situé le long du bord arrière du crâne qui forment des structures pointues appelées cornes tabulaires. La partie supérieure de la paroi externe de la cavité est également formée en partie des os squamosaux, qui occupent principalement la face supérieure plate du crâne. La partie du squamosal qui forme la paroi de la cavité est séparée de la partie situé à l'extérieur de la cavité par une limite prononcée connue sous le nom de crête falciforme[1].
La paroi externe de la cavité présente une rainure longue et prononcée, appelée rainure stapédienne, qui s'étend longitudinalement le long de la paroi. Le bord inférieur de cette rainure est formé par une crête connue sous le nom de crête oblique, bien qu'elle ait également été appelée crista obliqua ou plus simplement rebord otique. Le bord supérieur de la rainure stapédienne est formé par une autre crête bordant l'os squamosal, qu'Eltink et al. () nomment la « crête ptérygoïdienne dorsale »[5]. Cependant, l'étude de Marsicano et al. () préfèrent utiliser le nom de « lamelle » pour cette structure afin qu'elle ne soit pas confondue avec une crête différente présente chez les lydekkérinidés, parfois appelée « crête oblique du ptérygoïde », mais plus communément appelée « crête tympanique »[1],[5]. De manière assez confuse, de nombreux rhinésuchidés sont également connus pour posséder une crête tympanique. Cette crête est positionnée plus en arrière que les autres crêtes, près de l'intersection des os ptérygoïde, du carré et du squamosal, et s'étend vers le bas le long de la face arrière de la joue. Le bord intérieur de la paroi extérieure de la cavité est formé par un rebord que la plupart des études appellent simplement « membrane ». Cette convention existe à la suite de l'ancienne hypothèse et probablement incorrecte selon laquelle les encoches otiques abriteraient les tympans[6]. Selon cette hypothèse, le rebord interne peut être attaché à une membrane s'étendant le long de la cavité interne de l'oreille[5].
Cette combinaison de rainures et de crêtes de la cavité otique est unique aux rhinésuchidés. La lamelle et le sillon stapédien ne sont connus dans aucun autre temnospondyles, bien qu'ils soient présents dans pratiquement tous les rhinesuchidés à l'exception de Broomistega, qui n'a pas de lamelle. La crête tympanique est présente chez la plupart des rhinesuchidés mais absente chez quelques-uns, et elle est également présente chez les lydekkérinidés. La crête oblique et la crête falciforme sont présentes dans la plupart des autres stéréospondyles, bien que la première soit moins bien développée, tandis que le rebord « membranaire » est présent dans pratiquement tous les stéréospondylomorphes[5].
Palais et neurocrâne

Divers os et ouvertures comprennent le palais chez les rhinésuchidés, comme chez d'autres amphibiens. À l'extrémité du palais se trouvent les vomers, tandis que les zones proches du bord de la bouche sont constituées d'os palatins et d'ectoptérygoïdes. Au milieu de la partie arrière de la bouche se trouve un os rectangulaire connu sous le nom de parasphénoïde. La majeure partie du parasphénoïde forme la face inférieure du neurocrâne aplati, bien qu'il possède également une fine tige faisant saillie vers l'avant connue sous le nom de processus cultriforme, qui s'étend le long de la ligne médiane du crâne pour croiser les vomers. Vers l'arrière de la bouche, il y a des os ptérygoïdes disposants de plusieurs dents sur chaque côté du crâne. Chaque ptérygoïde ont plusieurs branches, dont la branche postérieure qui s'étend en arrière et sur le côté du crâne, la courte branche médiale qui s'étend vers l'intérieur et se connecte à l'os parasphénoïde, une branche ascendante qui se projette vers le haut pour former l'échancrure otique, et la branche antérieure qui s'étend vers l'avant le long du palatin et de l'ectoptérygoïde. Les ptérygoïdes de la plupart des rhinésuchidés ont de très longues branches antérieures. Dans la plupart des membres de cette famille, la branche antérieure atteint aussi loin que les vomers, bien qu'Australerpeton a des branches antérieures relativement courtes. Une paire de grandes ouvertures, connues sous le nom de vides interptérygoïdes, remplissent les zones entre ces os, créant ainsi la majorité de l'espace ouvert du palais[1],[5],[7].
Vue de derrière, les branches supérieures du neurocrâne s'étendent d'un côté à l'autre, masquant partiellement la branche ascendante des ptérygoïdes. Chaque processus paroccipital est également perforé d'un petit foramen, appelé fenêtre post-temporale. Ces foramens sont très fins chez les rhinésuchidés. Au-dessus de ces processus paroccipitaux se trouvent les échancrures otiques ainsi que les os tabulaires. Les processus paroccipitaux pointent également vers l'arrière dans une certaine mesure, formant des cornes qui, chez certains rhinesuchidés, sont légèrement plus longues que celles des tabulaires. Vu d'en bas, la partie la plus proéminente du neurocrâne est l'os parasphénoïde. Les coins arrière du parasphénoïde ont de petites « poches » bordées de crêtes appelées cristamuscularis[1]. Ces crêtes peuvent avoir des muscles ancrés capables de manœuvrer la tête sur le cou[6].
Autres caractéristiques du crâne et de la mâchoire
De nombreux os constituent la face supérieure du crâne, bien qu'une paire d'os particulière ait acquis une conception spécifique chez les rhinésuchidés. Ces os sont les os jugaux et préfrontaux allongés, qui forment le bord avant des orbites oculaires. Chez la plupart des rhinésuchidés, le bord entre les deux os possède une forme « étagée », avec une extension externe triangulaire du préfrontal poussant la suture avec le jugal vers une position plus latérale (situé vers l'extérieur). Cependant, la suture est plus droite chez Australerpeton, comme chez les autres stéréospondyles[1].
Les mandibules ont une paire de trous visibles uniquement depuis le bord intérieur de la mâchoire. Le plus grand trou à la partie arrière du complexe osseux, connu sous le nom de foramen de Meckel postérieur, est mince et allongé chez les rhinésuchidés. Un trou supplémentaire sur la face inférieure de l'articulation de la mâchoire n'est visible que d'en bas. Ce trou, la corde du tympan, est de grand taille dans cette famille. Sur la face supérieure de l'articulation de la mâchoire, une fine rainure connue sous le nom de rainure arcadienne s'étend vers le côté lingual de la mâchoire et sépare d'autres bosses osseuses situées entre l'articulation de la mâchoire. Dans l'ensemble, les rainures et les crêtes de l'articulation de la mâchoire sont peu développées chez les rhinesuchidés par rapport à celles de nombreux autres groupes de stéréospondyles, ressemblant plutôt à l'articulation simple des archégosauridés tels que Melosaurus[1].
Paléobiologie
Mode de vie
La plupart des rhinésuchidés ne sont connus qu'à partir de matériels crâniens, bien que quelques membres du groupe comme Uranocentrodon, Broomistega et Australerpeton incluent des spécimens préservant une partie importante du reste des squelettes. Un spécimen juvénile de Broomistega a des chevilles et des vertèbres mal ossifiées, indiquant que ses articulations auraient eu une grande quantité de matériel cartilagineux pour compléter la faible quantité d'os. Ce trait est souvent corrélé à un mode de vie aquatique. Les caractéristiques du crâne, telles que les orbites pointant vers le haut, soutiennent également cette hypothèse[8].
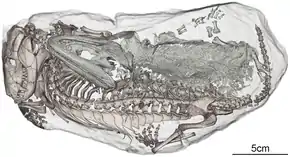
Malgré ce soutien à un mode de vie aquatique, d'autres éléments de preuves montrent que les rhinésuchidés auraient été capables d'un certain mouvement terrestre. Bien que les rhinésuchidés ne possèdent aucune adaptation pour creuser, un spécimen juvénile mal ossifié de Broomistega a été trouvé dans un terrier inondé qui était également habité par un thérapside cynodonte du genre Thrinaxodon[8]. Diverses conditions de conservation de ces animaux indiquent qu'ils cohabitaient paisiblement dans le terrier, susceptibles de survivre à une sécheresse en estivant, c'est-à-dire rester à l'état dormant pendant des conditions chaudes et sèches. Le fait qu'un Broomistega ait pu pénétrer dans le terrier d'un animal terrestre tel que Thrinaxodon indique que les rhinésuchidés ne seraient pas exclusivement aquatiques[8].
De plus, il est noté que les temnospondyles plus gros ont généralement des articulations bien plus ossifiées. Par exemple, de grands spécimens d'Australerpeton possèdent des hanches robustes, plusieurs os de la cheville complètement osseux et des pleurocentra ossifiées (une partie des vertèbres)[9]. Néanmoins, ces squelettes ne sont pas aussi fortement construits que ceux d'Eryops, un temnospondyle supposément terrestre, ayant une ceinture scapulaire plus petite et des sites moins proéminents pour l'attachement musculaire. Dias & Schultz suggèrent en que le mode de vie d'Australerpeton et vraisemblablement d'autres rhinésuchidés, est celui de piscivores semi-aquatiques, préférant chasser dans des plans d'eau douce peu profonds tout en conservant la capacité de marcher sur terre pendant les sécheresses[10].
Une étude histologique de plusieurs fossiles indéterminés de rhinésuchidés indique que les membres de la famille ont grandi de façon saisonnière, comme chez les amphibiens actuels. Les individus avaient également une durée de vie assez longue, un spécimen ayant entre 30 et 35 ans au moment de sa mort en fonction du nombre de lignes de croissance arrêtée (des anneaux dans l'os étant utilisés pour indiquer l'âge), présentes dans un fragment de hanche[11]. Certaines lignes de croissance arrêtée sont très étroites, indiquant que les individus pouvaient réduire leur croissance et leur métabolisme pendant les périodes difficiles. Cette capacité est peut être la raison pour laquelle les rhinésuchidés ont eu un succès évolutif vers la fin du Permien, ainsi que la façon dont quelques petits membres du groupe ont survécu à l'extinction qui en suivi[11].
Branchies
Trois rangées d'os minuscules (osselets branchiaux) recouverts de fines structures semblables à des dents (denticules branchiaux) ont été conservées près du cou d'un spécimen d'Uranocentrodon. Ces os seraient presque certainement attachés aux arcs branchiaux des branchies pendant que l'animal était vivant[4]. Bien que de tels os soient rares parmi les stéréospondyles et inconnus chez tous les autres rhinésuchidés, cela peut simplement être dû au fait que les os d'autres genres ont été préservés dans des sédiments plus rugueux où ces os délicats pourraient être cassés ou difficiles à trouver[12].
Bien qu'il soit évident qu'Uranocentrodon aurait eu des branchies, il est difficile de déterminer de quel type de branchies s'agissait-il. D'une part, il pourrait s'agir de branchies internes comme celles des poissons, peu visibles de l'extérieur du corps. D'autre part, ils auraient pu être des branchies externes en forme de tige comme celles des larves de salamandres modernes ou même des salamandres adultes néoténiques telles que Necturus ou l'axolotl. Les branchies externes ont dû évoluer à partir des branchies internes au cours de l'évolution des amphibiens, bien que l'emplacement précis de cette transition soit contesté. Les os de soutien des branchies conservés chez les anciens amphibiens présentent de nombreuses similitudes avec ceux des branchies des poissons et des salamandres. Les paléontologues qui préfèrent comparer les anciens tétrapodes aux amphibiens modernes trouvent généralement de nombreuses similitudes entre les os fossiles et les os branchiaux des salamandres modernes. D'autre part, les paléontologues qui comparent les tétrapodes fossiles aux poissons fossiles considèrent que les os sont en corrélation avec les branchies internes. Cette énigme, connue sous le nom de paradoxe de Bystrow, rendait difficile l'évaluation des branchies chez les anciens amphibiens tels qu'Uranocentrodon, car différents paléontologues sont arrivés à des conclusions différentes en fonction de leur domaine d'étude[13].

Le paradoxe de Bystrow est finalement résolu par une étude publié en 2011, qui révèle que les structures cérato-branchiales rainurées, qui sont les composants des arcs branchiaux, sont corrélées aux branchies internes. Les anciens tétrapodes conservant des structures cérato-branchiaux rainurés, comme le dvinosaurien Dvinosaurus, n'ont probablement eu que des branchies internes à l'âge adulte. Néanmoins, les branchies externes sont directement préservées en tant que tissus mous chez certains temnospondyles. Cependant, ces situations ne se produisent que chez les spécimens larvaires ou les membres de groupes spécialisés tels que les Branchiosauridae (en). Une espèce vivante de dipneuste (Lepidosiren) a des branchies externes sous forme de larves qui se transforment en branchies internes à l'âge adulte. Bien que les spécimens de dvinosaures adultes aient des caractéristiques squelettiques corrélées aux branchies internes, certains spécimens larvaires d'un autre dvinosaure, Isodectes, conservent les tissus mous de branchies externes. Ainsi, le développement des branchies des dvinosaures (et vraisemblablement d'autres temnospondyles, comme Uranocentrodon) refléterait celui de Lepidosiren. Bien que cette caractéristique soit probablement un exemple d'évolution convergente (car d'autres dipneustes possèdent exclusivement des branchies internes), elle reste un indicateur utile de la façon dont les branchies des temnospondyles se sont développées. Les auteurs de l'étude conclut que les branchies des temnospondyles, y compris Uranocentrodon et d'autres rhinésuchidés qui peuvent avoir possédé des branchies, seraient probablement internes comme ceux d'un poisson en tant qu'adultes, mais externes comme ceux d'une salamandre en tant que juvéniles[13].
Armures dermiques
Un squelette d'Uranocentrodon conserve également de grandes plaques d'écailles ou scutelles osseuses autour du corps. Les scutelles qui auraient été sur le ventre de l'animal sont disposées en rangées diagonales parallèles qui convergent vers la ligne médiane du corps et divergent lorsque les rangées s'étirent vers la queue. Chaque scutelle a une crête au milieu, et les scutelles plus loin vers la ligne médiane chevauchent celles plus éloignées. Le long de la ligne médiane, une rangée d'écailles plates et larges s'étendent de la gorge à la queue. Alors que ces écailles abdominales sont faites d'os, les écailles des autres parties du corps ont moins de structure osseuses et sont probablement faites de kératine à la place. Les écailles sur les côtés du corps sont plus plates et plus petites que les scutelles osseuses du ventre. Les scutelles à l'arrière du corps sont similaires, bien que de forme plus arrondie, avec quelques scutelles plus grandes près de la ligne médiane. Les écailles des membres postérieurs et le dessous de la région des hanches sont similaires à celles du dos, bien qu'aucun tégument ne soit conservé sur les membres antérieurs ou la queue. Ainsi, il est probable qu'au moins la queue aurait été sans armure et uniquement recouverte de peau nue[4].
Des écailles sont également conservées dans les spécimens d'Australerpeton. Ils ont une distribution similaire à ceux d'Uranocentrodon, mais sont généralement de forme plus ronde. Ils possèdent également une structure interne en forme de nid d'abeille et des caractéristiques histologiques indiquent qu'ils étaient profondément enfoncés dans la peau. Par conséquent, il est peu probable qu'ils auraient été visibles de l'extérieur du corps. Les chercheurs ne peuvent pas déterminer si les écailles des rhinésuchidés aurait permis ou restreint la respiration cutanée, c'est-à-dire une respiration passant à travers la peau comme chez les amphibiens modernes. D'autres applications potentielles des écailles comprennent la protection contre les prédateurs, la rétention d'eau pendant les sécheresses et peut-être même le stockage du calcium lorsque les conditions sont difficiles, une technique notamment utilisée par les représentants femelles des crocodiles du Nil. Cette dernière hypothèse est la moins probable, car les rhinésuchidés ne pondirent pas d'œufs à coquille dure comme chez les amniotes, ce qui est la raison pour laquelle les femelles crocodiles ont besoin de stocker du calcium[14].
Classification
Lorsque la famille fut nommée pour la première fois en 1919 par le paléontologue David Meredith Seares Watson[7], les rhinésuchidés sont immédiatement reconnus comme un groupe de stéréospondyles basaux, une position qui est toujours conservé de nos jours[1]. Parmi les traits utilisés pour soutenir cette position, citons le fait que la plupart des rhinésuchidés ont de longues branches antérieures de leurs ptérygoïdes, tandis que les stéréospondyles plus avancés ont des branches antérieures plus courtes. En 1947, Alfred Sherwood Romer place la famille, qui ne comprend que Rhinesuchus selon lui, dans une large super-famille qu'il désigne sous le nom de Rhinesuchoidea[15]. Le taxon Rhinesuchoidea était alors destiné à faire partie d'un grade évolutif de temnospondyles reliant les Rhachitomi « primitifs » tels qu'Eryops aux stéréospondyles « avancés » tels que les Metoposauridae et les Trematosauridae. Ce grade, appelé « Neorhachitomi », était séparé en Capitosauroidea (qui incluait les capitosaures et les benthosuchidés) et Rhinesuchoidea. En plus de contenir les Rhinesuchidae, Rhinesuchoidea contenait également divers autres genres ainsi que les familles Lydekkerinidae, Sclerothoracidae et Uranocentrodontidae. Romer estime que certains taxons (c'est-à-dire Uranocentrodon et le genre douteux potentiellement synonyme "Laccocephalus"[1]), souvent considérés comme des rhinésuchidés, serait mieux placés dans la famille distincte des Uranocentrodontidae, tandis que d'autres (c'est-à-dire Rhinesuchoides) ne sont placés dans aucune famille des Rhinesuchoidea en particulier[15]. D'autres familles sont ensuite placées dans les Rhinesuchoidea, comme les Rhinecepidae en 1966 et les Australerpetonidae en 1998[1].
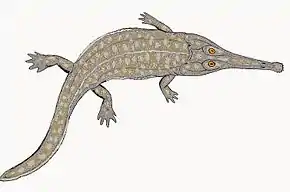
L'arrivée de la cladistique à la fin du XXe siècle fait tomber les grades en disgrâce ces dernières années, remplacés par des clades, qui sont définis par des relations étroites plutôt que par des assemblages ancestraux. Cependant, l'idée de base derrière les Rhinesuchoidea, qui stipule que les stéréospondyles avancés descendent d'animaux similaires aux rhinésuchidés, est toujours considérée comme valide. Les taxons Rhinecepidae et Uranocentrodontidae s'avèrent être synonymes du taxon Rhinesuchidae selon une analyse publié en 2000 par Schoch et Milner[16]. Une étude place les Rhinesuchidae dans la super-famille des Capitosauroidea[17]. Cependant, cette interprétation n'est pas suivie par d'autres études qui considèrent que les rhinésuchidés sont plus basaux que les capitosaures. Les Australerpetonidae, une famille monotypique ne contenant que le genre Australerpeton, est plus difficile à comparer aux Rhinesuchidae. Certaines études placent Australerpeton comme un stéréospondyle basal situé à l'extérieur des Rhinesuchidae[2],[9], tandis que d'autres le considèrent comme un Archegosauridae entièrement à l'extérieur des Stereospondyli[16].
Une revue publiée par Eltink et al. en 2016 privilégie l'hypothèse selon laquelle Australerpeton serait directement imbriqué dans les Rhinesuchidae. Une étude phylogénétique réalisée dans le cadre de l'étude divise la famille en deux clades. L'un des clade est une sous-famille appelée Rhinesuchinae, ce dernier contenant Rhinesuchus et Rhineceps. Cette sous-famille est principalement défini par des caractéristiques du palais, telles qu'une branche antérieure du ptérygoïde dépourvue de crêtes et des os palatins recouverts de minuscules denticules. L'autre clade principal de la famille contient Uranocentrodon ainsi qu'une autre sous-famille appelée Australerpetinae. Ce clade est uni par la présence d'une crête tympanique et d'un foramen magnum, un foramen de la moelle épinière situé à l'arrière du neurocrâne, qui a un bord supérieur incurvé. Australerpetinae est une version modifiée d'Australerpetonidae qui est réduite au statut de sous-famille afin de s'adapter dans les Rhinesuchidae. Cette sous-famille comprend Australerpeton, Broomistega, Laccosaurus et Rhinesuchoides. Les membres de cette sous-famille ont un museau un peu plus long et plus effilé que les autres rhinésuchidés, bien que selon Eltink et al., leurs ptérygoïdes ont de courtes branches antérieures, laissant les os palatins entrer en contact avec les vacuités interptérygoïdiennes. L'arbre le plus parcimonieux, c'est-à-dire le plus simple sur le plan de l'évolution, trouvé par Eltink et al. (2016) est montré ci-dessous[5] :
| ◄ Temnospondyli |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
La structure des Rhinesuchidae suivant l'étude d'Eltink et al. est remise en question par une étude différente sur les rhinesuchidés publiée moins d'un an plus tard. Cette étude, mené par Marsicano et al. en 2017, convient à dire qu'Australerpeton serait un rhinésuchidé, mais le considèrent comme le membre le plus basal de la famille. Ils ne sont pas d'accord avec la reconnaissance par Eltink et al. des branches ptérygoïdiennes antérieures courtes dans plusieurs genres. Selon leur analyse, seul Australerpeton possède ce trait, qui est la principale caractéristique qui le sépare du reste des Rhinesuchidae. D'autres traits qui soutiennent cette séparation incluent le fait que d'autres rhinesuchidés ont un contact jugal-préfrontal étagé et des os coronoïdes édentés dans la mâchoire inférieure. Le reste de la famille est mal résolu dans leur analyse phylogénétique, bien que trois clades aient des valeurs de support de Bremer modérées de 2[1].
Le soutien de Bremer est évalué en comptant le nombre de fois où des traits analysés sont acquis, perdus ou réacquis dans un arbre phylogénétique. Certains arbres phylogénétiques incluent plus de ces transitions que d'autres, ce qui signifie que certains arbres possibles supposent plus que le strict minimum d'évolution aurait eu lieu. L'arbre phylogénétique avec le moins de ces « étapes », ou transitions, est probablement le plus précis, basé sur le principe du rasoir d'Ockham (la réponse la plus simple et la plus précise). Le support de Bremer est utilisé pour étiqueter à quel point les clades sont bien supportés en analysant comment ils sont répartis entre des alternatives plus complexes à l'arbre le plus simple. Les clades qui n'existent pas dans un arbre phylogénétique et qui n'incarne qu'une étape totale plus complexe que l'arbre le plus parcimonieux ont un support de Bremer de 1, ce qui signifie que l'existence du clade est très incertaine. Même si l'arbre de la présente analyse soutient leur existence, de nouvelles données peuvent rendre un arbre phylogénétique concurrent plus parcimonieux, dissolvant des clades qui ne sont pris en charge que dans les arbres actuels. D'autres clades peuvent avoir des valeurs de support de Bremer beaucoup plus élevées, ce qui indique que des hypothèses plus drastiques doivent être formulées pour rendre le clade invalide. Le taxon Rhinesuchidae dans son ensemble, par exemple, a un support Bremer de 6 dans l'étude publié par Marsicano et al. en 2017, ce qui est considéré comme un soutien élevé. Un support Bremer de 2, comme c'est le cas avec trois clades spécifiques dans cette analyse, est considéré comme modéré. L'un de ces clades comprend les deux espèces valides de Rhinesuchoides, tandis qu'un autre clade relit Rhineceps et Uranocentrodon, et le dernier contient Rhinesuchus et Laccosaurus. L'arrangement de ces clades ainsi que le placement de Broomistega n'a pas pu être résolu avec une certitude absolue, ayant des valeurs de soutien de Bremer de seulement 1, quel que soit l'endroit où les trois clades sont placés hors d'Australerpeton. L'arbre le plus parcimonieux trouvé par Marsicano et al. (2017) est montré ci-dessous[1] :
| ◄ Temnospondyli |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Notes et références
Notes
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Rhinesuchidae » (voir la liste des auteurs).
- À titre de comparaison, les plus grands amphibiens existants actuellement sont les urodèles de la famille des Cryptobranchidae, dont les tailles maximalles vont de 70 centimètres à 1,8 mètres de long[3].
Références
- (en) C.A. Marsicano, E. Latimer, B. Rubidge et R.M.H. Smith, « The Rhinesuchidae and early history of the Stereospondyli (Amphibia: Temnospondyli) at the end of the Palaeozoic », Zoological Journal of the Linnean Society, vol. 181, no 2, , p. 357–384 (DOI 10.1093/zoolinnean/zlw032
 , S2CID 89770655)
, S2CID 89770655) - (en) Rainer R. Schoch, « The evolution of major temnospondyl clades: an inclusive phylogenetic analysis », Journal of Systematic Palaeontology, vol. 11, no 6, , p. 673–705 (DOI 10.1080/14772019.2012.699006, S2CID 83906628)
- (en) R. K. Browne, H. Li, Z. Wang, S. Okada, P. Hime, A. McMillan, M. Wu, R. Diaz, D. McGinnity et J. T. Briggler, « The giant salamanders (Cryptobranchidae): Part B. Biogeography, ecology and reproduction », Amphibian and Reptile Conservation, vol. 5, no 4, , p. 30-50 (lire en ligne [archive du ])
- (en) E.C.N. Van Hoepen, « Stegocephalia of Senekal, O.F.S », Annals of the Transvaal Museum, vol. 5, no 2, , p. 125-149 (DOI 10520/AJA00411752_812 , S2CID 134584405, lire en ligne [PDF])
- (en) Estevan Eltink, Eliseu V. Dias, Sérgio Dias-da-Silva, Cesar L. Schultz et Max C. Langer, « The cranial morphology of the temnospondyl Australerpeton cosgriffi (Tetrapoda: Stereospondyli) from the Middle-Late Permian of Paraná Basin and the phylogenetic relationships of Rhinesuchidae », Zoological Journal of the Linnean Society, vol. 176, no 4, , p. 835–860 (ISSN 0024-4082, DOI 10.1111/zoj.12339 , S2CID 85571728)
- (en) D.M.S. Watson, « The Evolution of the Labyrinthodonts », Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, vol. 245, no 723, , p. 219–265 (DOI 10.1098/rstb.1962.0010, JSTOR 2416605, S2CID 85817748)
- (en) D. M. S. Watson, « I. The structure, evolution and origin of the Amphibia. - The "orders' Rachitomi and Stereospondyli », Philosophical Transactions of the Royal Society of London. Series B, Containing Papers of a Biological Character, vol. 209, nos 360-371, , p. 1–73 (ISSN 0264-3960, DOI 10.1098/rstb.1920.0001 , JSTOR 92046, S2CID 86393316, lire en ligne [PDF])
- (en) Vincent Fernandez, Fernando Abdala, Kristian J. Carlson, Della Collins Cook, Bruce S. Rubidge, Adam Yates et Paul Tafforeau, « Synchrotron Reveals Early Triassic Odd Couple: Injured Amphibian and Aestivating Therapsid Share Burrow », PLOS ONE, vol. 8, no 6, , e64978 (ISSN 1932-6203, PMID 23805181, PMCID 3689844, DOI 10.1371/journal.pone.0064978 )
- (en) Estevan Eltink et Max C. Langer, « A New Specimen of the Temnospondyl Australerpeton cosgriffi from the Late Permian of Brazil (Rio Do Rasto Formation, Paraná Basin): Comparative Anatomy and phylogenetic relationships », Journal of Vertebrate Paleontology, vol. 34, no 3, , p. 524–538 (DOI 10.1080/02724634.2013.826667, JSTOR 24523275, S2CID 86083995, lire en ligne [PDF])
- (en) E.V. Dias et C.L. Schultz, « The first Paleozoic temnospondyl postcranial skeleton from South America », Revista Brasileira de Paleontologia, vol. 6, , p. 29–42 (S2CID 59936069, lire en ligne [PDF])
- (en) Julia B. McHugh, « Paleohistology and histovariability of the Permian stereospondyl Rhinesuchus », Journal of Vertebrate Paleontology, vol. 34, no 1, , p. 59–68 (DOI 10.1080/02724634.2013.787429, JSTOR 24523251, S2CID 83906138, lire en ligne)
- (en) Rainer R. Schoch, « The evolution of metamorphosis in temnospondyls », Lethaia, vol. 35, no 4, , p. 309–327 (ISSN 1502-3931, DOI 10.1111/j.1502-3931.2002.tb00091.x, S2CID 83496782)
- (en) Rainer R. Schoch et Florian Witzmann, « Bystrow's Paradox - gills, fossils, and the fish-to-tetrapod transition », Acta Zoologica, vol. 92, no 3, , p. 251–265 (ISSN 1463-6395, OCLC 1461033, DOI 10.1111/j.1463-6395.2010.00456.x, S2CID 85651134, lire en ligne)
- (en) E.V. Dias et M. Richter, « On the squamation of Australerpeton cosgriffi Barberena, a temnospondyl amphibian from the Upper Permian of Brazil », Anais da Academia Brasileira de Ciências, vol. 74, no 3, , p. 477–490 (ISSN 0001-3765, DOI 10.1590/S0001-37652002000300010 , S2CID 12971428, lire en ligne [PDF])
- (en) Alfred Sherwood Romer, « Review of the Labyrinthodontia », Bulletin of the Museum of Comparative Zoology at Harvard College, vol. 99, no 1, , p. 7–368 (OCLC 253748351, lire en ligne)
- (en) R.R. Schoch et A.R. Milner, Stereospondyli, stem-Stereospondyli, Rhinesuchidae, Rhitidostea, Trematosauroidea, Capitosauroidea, vol. 3B, Munich, , 1–203 p.
- (en) M. A. Shishkin et B. S. Rubidge, « A Relict Rhinesuchid (Amphibia: Temnospondyli) From The Lower Triassic Of South Africa », Palaeontology, vol. 43, no 4, , p. 653–670 (ISSN 1475-4983, DOI 10.1111/1475-4983.00144 , S2CID 128561620, lire en ligne [PDF])
Voir aussi
Articles connexes
Liens externes
- Ressources relatives au vivant :
 Portail de la paléontologie
Portail de la paléontologie  Portail de l’herpétologie
Portail de l’herpétologie

Le 4 septembre 2022, Rhinesuchidae a été proposé pour être reconnu comme un « bon article ». Vous pouvez donner votre avis sur cette proposition. Suivi des modifications de l'article depuis la proposition.