كيناز البروتين
كيناز البروتين(1) هو إنزيمُ كيناز يُعدلُ البروتينات الأخرى عبر إضافة مجموعات فوسفات كيميائيًا إليها، فيما يُعرف بالفسفرة. تؤدي الفسفرةُ عادةً إلى تغييرٍ وظيفي في البروتين المُستهدف (الركيزة) عبر تغيير نشاط الإنزيم أو الموقع الخلوي أو الارتباط ببروتيناتٍ أخرى. يحتوي الجينوم البشري على حوالي 500 جينٍ لكيناز البروتين، حيثُ تشكل حوالي 2% من مجموع الجينات البشرية.[1] توجد كينازات البروتين أيضًا في البكتيريا والنباتات. قد يُعدل حوالي 30% من جميع البروتينات البشرية عبر نشاط كيناز، ومن المعروف أن الكينازات تُنظم معظم المسارات الخلوية، وخاصة تلك التي تشارك في توصيل الإشارة.
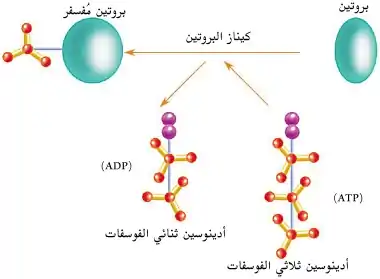
النشاط الكيميائي

يتضمن النشاطُ الكيميائي للكيناز إزالة مجموعة فوسفات من أدينوسين ثلاثي الفوسفات (ATP) وربطها تساهميًا بأحد الأحماض الأمينية الثلاثة التي تحتوي على مجموعة هيدروكسيل حُرة. تعمل مُعظم الكينازات على السيرين والثريونين معًا، ويعملُ البعض الآخر على التيروسين، ويعمل عددٌ آخر (كيناز ثنائي النوعية) على الثلاثة معًا.[2] هناك أيضًا كينازات بروتين تُفسفرُ الأحماض الأمينية الأخرى، ومنها كينازات الهستيدين التي تُفسفر بقايا الهستيدين.[3]
التنظيم
يُعتبر نشاط كينازات البروتين منظمًا للغاية؛ وذلك لأنَّ لها تأثيرٌ كبيرٌ على الخلية. تُشغل أو توقف الكينازات بواسطة الفسفرة (أحيانًا عن طريق الكيناز نفسه فسفرة مقرونة (cis)/فسفرة ذاتية)، عن طريق ربط البروتينات المنشطة أو البروتينات المثبطة، أو الجزيئات الصغيرة، أو عن طريق التحكم في موقعها في الخلية بالنسبة إلى ركائزها.
البنية
الوحدات الفرعية التحفيزية للعديد من كينازات البروتين محفوظة بشدة، والعديد من بُنى هذه الكينازات تم حلها وتحديدها.[4]
كينازات البروتين الخاصة بحقيقيات النوى هي إنزيمات تنتمي إلى عائلة كبيرة جدا من البروتينات التي تتشارك لب تحفيزٍ محفوظ.[5][6][7][8] توجد العديد من المناطق المحفوظة في النطاق التحفيزي لكينازات البروتين. في النهاية الأمينية للنطاق التحفيزي يوجد امتداد غني بوحدات الغليسين بجوار وحدة ليسين، والذي تم إظهار أن له دورا في ارتباط الـATP. في الجزء المركزي من النطاق التحفيزي، يوجد حمض أسبارتيك محفوظ، وهو ضروري للنشاط التحفيزي الخاص بالإنزيم.[9]
كينازات البروتين المختصة بالسيرين/ثريونين
تُفسفر كينازات البروتين المختصة بالسيرين والثريونين ( ر.ت.إ 2.7.11.1 ) مجموعة الهيدروكسيل في السيرين أو الثريونين (اللذين لهما نفس السلسلة الجانبية). يمكن تنظيم نشاط كينازات البروتين هذه بواسطة أحداث معينة (مثل تضرر الدنا)، وكذلك بمجموعة من الإشارات الكيميائية منها cAMP/cGMP وثنائي الغليسيريد والكالسيوم/كالموديولين. أحد أهم مجموعات كيناز البروتين هي كينازات ماب (MAP) (كيناز البروتين المنشط بالميتوجين). ومن المجموعات الفرعية المهمة: العائلة الفرعية ERK المنشطة عادة بإشارات ميتوجينية، وكينازات البروتين المنشطة بالإجهاد JNK وp38. رغم أن كينازات MAP مختصة بالسيرين/ثريونين، فإن تنشيطها يتم عبر فسفرة مشتركة على وحدات السيرين/ثريونين والتيروسين. نشاط كينازات MAP مقيد بواسطة عدد من فوسفاتازات البروتين التي تزيل مجموعات الفوسفات التي أضيفت إلى وحدات سيرين وثريونين محددة، وهي مطلوبة للحفاظ على الكينازات في الهيئة البنيوية النشطة.
عاملان أساسيان يؤثران على نشاط كينازات MAP: أ) الإشارات التي تنشط المستقبلات عبر الغشائية (سواء الربائط الطبيعية أو عوامل التشبيك) والبروتينات المرتبطة بها (الطفرات التي تحاكي حالة النشاط). ب) الإشارات التي تثبط الفوسفاتازات التي تقيد عمل كينازات MAP. ومن هذه الإشارات الإجهاد التأكسدي.[10]
كينازات البروتين المختصة بالتيروسين
تُفسفِر كينازات البروتين المختصة بالتيروسين ( ر.ت.إ 2.7.10.1 و ر.ت.إ 2.7.10.2 ) وحدات الأحماض الأمينية تيروسين وتُستخدم في توصيل الإشارة مثل كينازات البروتين المختصة بالسيرين/ثريونين. يعمل هذا النوع من الكينازات أساسا كمستقبلات عوامل النمو، وفي تفاعلات التأشير مع المسار بعد استقبال عوامل النمو،[11] ومن بعض الأمثلة:
- مستقبل عامل النمو المشتق من الصفيحات (PDGFR).
- مستقبل عامل النمو البشروي (EGFR).[12]
- مستقبل الأنسولين ومستقبل عامل النمو المشابه للأنسولين 1 (IGF1R)
- مستقبل عامل الخلية الجذعية ويسمى كذلك c-kit (طالع مقال ورم معدي معوي).
كينازات التيروسين المستقبلة
تتكون هذه الكينازات من مستقبلات عبر غشائية مع نطاقِ كينازِ تيروسينٍ ممتد إلى السيتوبلازم، وتلعب دورا مهما في تنظيم الانقسام الخلوي، التمايز الخلوي والتشكل الحيوي. أزيد من 50 كيناز تيروسين مستقبل معروف لدى الثدييات.
البنية
النطاق خارج الخلوي هو الجزء الذي يرتبط بالربيطة من الجزيء، ويمكن أن يكون وحدة منفصلة تتصل بباقي المستقبل عبر رابطة ثنائي كبريتيد. يمكن أن تُستخدم نفس الآلية لربط مستقبلين معا لتشكيل مثنوي متماثل أو متغاير. الجزء عبر الغشائي هو لولب ألفا تأشيري. النطاق داخل الخلوي أو السيتوبلازمي مسؤول على نشاط الكيناز (المحفوظ بشدة) وكذلك على العديد من الوظائف التنظيمية.
التنظيم
يسبب ارتباط الربائط تفاعلين:
- تثني (تشكيل مثنوي) الكينازات المستقبلة الموحودية أو استقرار مثنوي متهدل (غير مستقر). العديد من ربائط كينازات التيروسين المستقبلة متعددة التكافؤ. بعض كينازات التيروسين المستقبلة (مثل مستقبل عامل النمو المشتق من الصفيحات يمكن أن تشكل مثنويات متغايرة مع كينازات أخرى مشابهة لكنها غير متطابقة من نفس العائلة الفرعية، وهذا يسمح باستجابات شديدة التنوع للإشارة خارج الخلوية.
- فسفرة ذاتية-مفروق (فسفرة بواسطة الكيناز الآخر في المثنوي) للكيناز المستقبل للربيطة.
تسبب الفسفرة الذاتية انزياح النطاقان الفرعيان للكيناز المستقبل وهذا يفتح نطاق الكيناز لارتباط الـATP. في الهيئة غير النشطة النطاقان الفرعيان للكيناز متراصفان بشكل يمنع الـATP من الوصول إلى المركز التحفيزي الخاص بالكيناز. حين تتواجد عدة أحماض أمينية قابلة للفسفرة في نطاق الكيناز (مثل مستقبل عامل النمو المشابه للأنسولين 1) يمكن أن يزيد نشاط الكيناز بازدياد عدد الأحماض الأمينية المفسفرة، في هذه الحالة يقال بأنها فسفرة ذاتية-مقرون تحوِّل الكيناز من «الخمود» إلى «الاستعداد»
توصيل الإشارة
يفسفِر كيناز التيروسين النشط بروتينات مستهدفة محددة، والتي هي إنزيمات في حد ذاتها. أحد أهداف هذه الكينازات هو بروتينات راس التي تحدث سلسلة تفاعلات لتوصيل الإشارة.
كينازات التيروسين المرتبطة بالمستقبل
تسمى كينازات التيروسين التي يتم توظيفها إلى مستقبل بعد ارتباط هرومون «كينازات التيروسين المرتبطة بالمستقبل» ولها دور في العديد من تفاعلات التأشير المتسلسلة، وبشكل خاص تلك التي لها دور في تأشير السيتوكين (وكذلك نواقل إشارة أخرى بما في ذلك هرمونات النمو). أحد كينازات التيروسين المرتبطة بالمستقبل هو كيناز جانوس (JAK) الذي تتوسط بروتينات ستات (STAT) العديد من تأثيراته. (انظر مسار JAK-STAT).
كينازات البروتين الخاصة بالهستيدين
كينازات الهستيدين متميزة بنيويا عن كينازات البروتين الأخرى وتتواجد في الغالب لدى بدائيات النوى كجزء من آليات نقل إشارة مكونة من مكونين. تُضاف أولا مجموعة الفوسفات من الـATP إلى وحدة هستيدين داخل الكيناز ثم تُنقل لاحقا إلى وحدة أسبارتات على «النطاق المستقبل» لبروتين مختلف وأحيانا للكيناز في حد ذاته. وبعد ذلك تنشط وحدة الأسبارتيل فوسفات هذه في توصيل الإشارة.
تتواجد كينازات الهستيدين بشكل واسع لدى بدائيات النوى، وكذلك في النبات، الفطريات وحقيقيات النوى. عائلة نازعة هيدروجين البيروفات من الكينازات لدى الحيوانات ذات صلة بنيوية بكينازات الهستيدين، لكن تقوم بفسفرة وحدات السيرين بدل الهستيدين، وربما لا تستخدم وسيط الفوسفو-هستيدين.
كينازات مختلطة
لبعض الكينازات نشاطات مختلطة. على سبيل المثال كيناز كيناز البروتين المنشط بالميتوجين (MAPKK) الذي له دور في سلسلة تفاعلات كيناز MAP له وظيفة كيناز السيرين/ثريونين ووظيفة كيناز التيروسين وبذلك هو كيناز مزدوج التخصص.
مثبطات
يُعتبر عدم انتظام نشاط الكيناز من الأسباب الشائعة لحدوث الأمراض، وخاصةً السرطان، حيث ينظم الكيناز جوانب عديدة تتحكم في نمو الخلايا وحركتها وموتها. يتم تطوير أدويةٌ تثبط كينازاتٍ معينةٍ لعلاج العديد من الأمراض، وبعضها قيد الاستخدام السريري حاليًا، وتشمل غليفيك (إيماتينب) وآيريسا (جيفيتينيب).
الهوامش
1. كِينازُ البرُوتين[ِ 1][ِ 2][ِ 3][ِ 4] (بالإنجليزية: Protein kinase)، حرفيًا بروتين كيناز.[ِ 4]
المراجع
باللغة الإنجليزية
- Manning G, Whyte DB, Martinez R, Hunter T, Sudarsanam S (2002)، "The protein kinase complement of the human genome"، Science، 298 (5600): 1912–1934، doi:10.1126/science.1075762، PMID 12471243.
- Dhanasekaran N, Premkumar Reddy E (سبتمبر 1998)، "Signaling by dual specificity kinases"، Oncogene، 17 (11 Reviews): 1447–55، doi:10.1038/sj.onc.1202251، PMID 9779990.
- Besant PG, Tan E, Attwood PV (مارس 2003)، "Mammalian protein histidine kinases"، Int. J. Biochem. Cell Biol.، 35 (3): 297–309، doi:10.1016/S1357-2725(02)00257-1، PMID 12531242، مؤرشف من الأصل في 01 يوليو 2018.
- Stout TJ, Foster PG, Matthews DJ (2004)، "High-throughput structural biology in drug discovery: protein kinases"، Curr. Pharm. Des.، 10 (10): 1069–82، doi:10.2174/1381612043452695، PMID 15078142، مؤرشف من الأصل في 21 مارس 2020.
- Hanks SK (2003)، "Genomic analysis of the eukaryotic protein kinase superfamily: a perspective"، Genome Biol.، 4 (5): 111، doi:10.1186/gb-2003-4-5-111، PMC 156577، PMID 12734000، مؤرشف من الأصل في 19 أبريل 2015.
- Hanks SK, Hunter T (مايو 1995)، "Protein kinases 6. The eukaryotic protein kinase superfamily: kinase (catalytic) domain structure and classification"، FASEB J.، 9 (8): 576–96، PMID 7768349، مؤرشف من الأصل في 13 مارس 2020.
- Hunter T (1991)، "Protein kinase classification"، Meth. Enzymol.، 200: 3–37، doi:10.1016/0076-6879(91)00125-G، PMID 1835513.
- Hanks SK, Quinn AM (1991)، "Protein kinase catalytic domain sequence database: identification of conserved features of primary structure and classification of family members"، Meth. Enzymol.، 200: 38–62، doi:10.1016/0076-6879(91)00126-H، PMID 1956325.
- Knighton DR, Zheng JH, Ten Eyck LF, Ashford VA, Xuong NH, Taylor SS, Sowadski JM (يوليو 1991)، "Crystal structure of the catalytic subunit of cyclic adenosine monophosphate-dependent protein kinase"، Science، 253 (5018): 407–14، doi:10.1126/science.1862342، PMID 1862342.
- Vlahopoulos S, Zoumpourlis VC. JNK: a key modulator of intracellular signaling. Biochemistry (Mosc). 2004 Aug;69(8):844-54. Review. PMID 15377263
- Higashiyama S, Iwabuki H, Morimoto C, Hieda M, Inoue H, Matsushita N. Membrane-anchored growth factors, the epidermal growth factor family: beyond receptor ligands. Cancer Sci. 2008 Feb;99(2):214-20. Review. PMID 18271917
- Carpenter G. The EGF receptor: a nexus for trafficking and signaling. Bioessays. 2000 Aug;22(8):697-707. Review. PMID 10918300
باللغة العربيَّة
- "ترجمة (Protein kinase) في المعجم الصيدلي الموحد"، مكتبة لبنان ناشرون، مؤرشف من الأصل في 13 مارس 2020، اطلع عليه بتاريخ 10 فبراير 2020.
- "ترجمة (Protein kinase) في المعجم الطبي الموحد"، مكتبة لبنان ناشرون، مؤرشف من الأصل في 11 مارس 2020، اطلع عليه بتاريخ 10 فبراير 2020.
- "ترجمة (Protein kinase) في قاموس المعاني"، قاموس المعاني، مؤرشف من الأصل في 13 مارس 2020، اطلع عليه بتاريخ 10 فبراير 2020.
- "ترجمة (Protein kinase) في موقع القاموس"، alqamoos، مؤرشف من الأصل في 13 مارس 2020، اطلع عليه بتاريخ 10 فبراير 2020.
مراجع إضافية
روابط خارجية
- Human and mouse protein kinases in UniProt: classification and index
- Kinase.Com: Genomics, evolution and large-scale analysis of protein kinases (موقع غير تجاري).
- KinMutBase: A registry of disease-causing mutations in protein kinase domains
- Human kinome by Manning et al.
- KLIFS (Kinase-Ligand Interaction Fingerprints and Structures) Database -- analysis of kinase/inhibitor interactions
- Orientations of C1 domains of protein kinases in membranes
- Orientations of C2 domains of protein kinases in membranes
 بوابة علم الأحياء
بوابة علم الأحياء بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية