ORF1ab
ORF1ab (يُعرف كذلك بـ ORF1a/b) هو جين يجمع إطاري القراءة المفتوحين (ORFs) ORF1a وORF1b، المحفوظان في جينومات الفيروسات العشية، وهي رتبة تحتوي على فصيلة فيروسات كورونا. يُعبر هذا الجين على عديد بروتين كبير يخضع لتحلل بروتيني تنتج منه عدة بروتينات لابنيوية ذات وظائف مختلفة في دورة حياة الفيروس، منها بروتيازات ومكونات مركب ريبلكاز-ترانسكريبتاز (RTC).[1][2][3] أحيانا يُشار إلى إطاري القراءة المفتوحين معا بجين الريبلكاز.[4] وهما متصلان عبر انزياح إطار ريبوسومي مبرمج يسمح للريبوسوم بالاستمرار في الترجمة بعد كودون التوقف الموجود في نهاية ORF1a في إطار قراءة -1 (أي يعود إلى الخلف بنوكليوتيد واحد). تُعرف عديدات البروتين الناتجة بـ:pp1a وpp1ab.[1][2][3][4]
 الترتيب الجينومي لمستخلصة Wuhan-Hu-1، وهي أول عينة من سارس-كوف-2 حُدد تسلسلها، توضح موقع ORF1a وORF1a في الجينوم. | |
| معرف الجينوم في NCBI | 86693 |
|---|---|
| حجم الجينوم | 29,903 bases |
| مشروع الجينوم | 2020 |
| عديد بروتين الريبلكاز | |
|---|---|
| معرفات | |
| الكائن | |
| الرمز | rep |
| يونيبروت | P0DTD1 |
| معلومات أخرى | |
التعبير
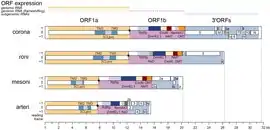
ORF1a هو إطار القراءة المفتوح الأول في النهاية 5' من الجينوم. يشغل الجين ORF1ab حوالي ثلثي الجينوم، ويشفر الثلث الباقي في النهاية 3' البروتينات البنيوية والبروتينات الملحقة.[1][2][3] ويُترجم هذا الجين من الرنا المقبع 5' بترجمة معتمدة على القبعة.[1] لدى الفيروسات العشية نظام معقد لإنتاج جزيئات رنا جينومية فرعية متقطعة يسمح لها بالتعبير عن الجينات من جينوماتها الرنوية الكبيرة نسبيا (عادة 27-32 ألف بالنسبة لفيروسات كورونا[1]) لكن ORF1ab يُترجم مباشرة من الرنا الجينومي.[5] لوحظت تسلسلات ORF1ab في جزيئات رنا جينومية فرعية غير معيارية، وأهميتها الوظيفية غير واضحة.[5]
يسمح انزياح إطار ريبوسومي مبرمج بالقراءة عبر كودون التوقف الذي يُنهي الترجمة في ORF1a والاستمرار في إطار قراءة (1-) وإنتاج عديد البروتين الأطول pp1ab. يحدث انزياح الإطار في تسلسل زلق تليه بنية رنا ثانوية على شكل عقدة كاذبة.[1] وتم قياس فعالية انزياح الإطار بين 20-50% لدى فيروس كورونا الفأري،[6] أو 45-70% لدى سارس-كوف-2،[7] وهو ما يعني التعبير عن بروتين pp1a حوالي 1.5 إلى 2 مرة أكثر من البروتين pp1ab.[2]
الوسط: ترتيب نطاقات بروتين nsp14 (نوكلياز خارجي وناقل ميثيل).
الأسفل: مكونات مركب الريبلكاز-ترانسكريبتاز الخاص بفيروس كورونا.[8]
المعالجة
يحتوي عديدا البروتين pp1a وpp1ab على حوالي 13 إلى 17 بروتين لابنيوي.[3] ويخضعان لتحلل بروتيني ذاتي لإنتاج بروتينات لابنيوية (nsp) تقوم به نطاقات بروتياز السيستين.[1][2][3]
يوجد في فيروسات كورونا 16 بروتينا لابنيويا، يحتوي عديد البروتين pp1a على البروتينات اللابنيوية nsp1-11 ويحتوي عديد البروتين pp1ab على nsp1-10 وnsp12-16. يقوم بالمعالجة التحللية (القص) بروتيازان: نطاق بروتياز مماثل للبابايين يتواجد في البروتين nsp3 متعدد النطاقات ويقص البروتينات من nsp1 حتى nsp4، والبروتياز 3CL (ويُعرف بالبروتياز الرئيسي، وهو البروتين nsp5) يقوم بباقي عمليات القص حتى النهاية الكربوكسيلية لعديد البروتين.[1][2] تحتوي البروتينات nsp12-16 المكونة للنهاية الكربوكسيلية لعديد البروتين pp1ab على مركز النشاطات الإنزيمية الضرورية لتضاعف الفيروس. بعد المعالجة التحللية، تتجمع البروتينات غير الوظيفية إلى مركب بروتيني كبير يُعرف بمركب ريبلكاز-تراسكريبتاز (RTC) الذي يقوم بنسخ ومضاعفة الفيروس.[1][2]
المكونات
نطاقات لب الناسخ
توجد 5 نطاقات بروتينية محفوظة في «لب الناسخ» لدى جميع سلالات الفيروسات العشية (الشريانية، العشية المتوسطة [الإنجليزية]، الرونية، وكورونا) وهي: من الـORF1a البروتياز الرئيسي وعلى طرفيه نطاقين عبر غشائيين، من الـORF1b نطاق ناقل النوكليوتيديل [الإنجليزية] ويسمى NiRAN، بوليمراز الرنا المعتمد على الرنا (RdRp)، نطاق مرتبط بالزنك، وهيليكاز.[3][9] (وقد تُعتبر هذه 7 نطاقات عند احتساب المناطق عبر غشائية بشكل منفصل.[4]) يوجد نطاق ريبونوكلياز داخلي لدى جميع الفيروسات العشية التي تصيب البشر. لا تملك الفيروسات الشريانية -التي لديها أصغر جينوم في السلالات العشية- ناقلات ميثيل ولا ريبونوكليازات خارجية مصححة، وهو نطاق محفوظ لدى الفيروسات العشية كبيرة الجينوم.[3] يُعتقد أن وظيفة التصحيح هذه مطلوبة من أجل دقة كافية لتضاعف جينومات الرنا، لكن يمكن أن تكون لها أدوار إضافية أخرى عند بعض الفيروسات.[9]
فيروسات كورونا
في فيروسات كورونا، يحتوي عديدا البروتين pp1a وpp1ab معا على 16 بروتينا لابنيويا تملك الوظائف التالية:[1][2][10][11]
| البروتين اللابنيوي | الوظيفة |
|---|---|
| البروتين اللابنيوي 1 | تفكيك الرنا الرسول الخلوي، تثبيط الترجمة الخاصة بالخلية المضيفة، تثبيط الإنترفيرون، غير متواجد لدى فيروسات كورونا غاما |
| البروتين اللابنيوي 2 | غير معروفة، يرتبط بالبروهيبتين |
| البروتين اللابنيوي 3 | بروتين متعدد النطاقات مع نطاق أو اثنين وظيفتهما ببروتياز مماثل للبابايين لقص البروتينات، مناهض للإنترفيرون، عدة أدوار أخرى. |
| البروتين اللابنيوي 4 | تكوين الحويصلات مزدوجة الغشاء |
| البروتين اللابنيوي 5 | بروتياز 3CL لمعالجة وقص عديد البروتين، تثبيط الإنترفيرون |
| البروتين اللابنيوي 6 | تكوين الحويصلات مزدوجة الغشاء |
| البروتين اللابنيوي 7 | عامل مرافق وعامل عملياتية [الإنجليزية] لبوليمراز الرنا المعتمد على الرنا (RdRp)، يشكل مركبا مع nsp8 وnsp12. |
| البروتين اللابنيوي 8 | عامل مرافق وعامل عملياتية لبوليمراز الرنا المعتمد على الرنا، يشكل مركبا مع nsp7 وnsp12 |
| البروتين اللابنيوي 9 | يرتبط بالرنا مفرد السلسلة |
| البروتين اللابنيوي 10 | عامل مرافق للبروتينين nsp14 و nsp16. |
| البروتين اللابنيوي 11 | غير معروفة |
| البروتين اللابنيوي 12 | بوليمراز الرنا المعتمد على الرنا، وناقل النوكليوتيديل [الإنجليزية] |
| البروتين اللابنيوي 13 | هيليكاز، وثلاثي فوسفاتاز الرنا. |
| البروتين اللابنيوي 14 | نوكلياز خارجي مصحح، تكوين قبعة الرنا، ناقل ميثيل-N7 غوانوزين. |
| البروتين اللابنيوي 15 | ريبونوكلياز داخلي، تجنب المناعة. |
| البروتين اللابنيوي 16 | ناقل ميثيل2'-O- ريبوز، تكوين قبعة الرنا. |
التطور
بنية وترتيب الجينوم -بما في ذلك الجينان ORF1a وORF1b وانزياح الإطار الذي يفصل بينهما- محفوظة بين الفيروسات العشية، وُصفت بعض البُنى الجينومية «غير المعيارية» لدى الفيروسات العشية، ولها علاقة بالأساس مع اندماج الجينات [الإنجليزية].[4] يملك أكبر فيروس عشي معروف وهو الفيروس العشي للخلية المفرزة للمستورقة [الإنجليزية] (41 ألف زوج قاعدي) بنية جينوم غير معيارية تكون فيها: الإطارات ORF1a وORF1b وباقي الإطارات مع الاتجاه التي تشفر بروتينات بنيوية وملحقة مدمجة معا، ويُعبر عن إطار قراءة مفتوح مفرد كبير يتجاوز 13 ألف حمض أميني.[4][12] في هذه الجينومات غير المعيارية، يمكن أن تُستخدم مناطق انزياح إطار أخرى أو قراءات عبر كودون التوقف لتنظيم نسبة إنتاج البروتينات الفيروسية.[4]
تختلف الفيروسات العشية كثيرا في حجم الجينوم، من الفيروسات الشريانية بجينوم طوله حوالي 12-15 ألف إلى فيروسات كورونا بطول 27-32 ألف. تاريخها التطوري هو محل اهتمام البحوث العلمية لفهم تضاعف جينوماتٍ رنويةٍ كبيرة رغم انخفاض دقة آلية التضاعف الخاصة ببوليمراز الرنا المعتمد على الرنا.[4] تُشفر الفيروسات العشية كبيرة الجينوم (فوق 20 ألف) ريبونوكلياز خارجي مصحح (البروتين اللابنيوي 14 في فيروسات كورونا)، ويُعتقد أنه ضروري لدقة التضاعف.[1][9]
بين فيروسات كورونا، إطار القراءة المفتوح ORF1ab أكثر انحفاظا من الإطارات 3' المشفرة للبروتينات البنيوية.[11] خلال جائحة كوفيد 19، تمت سَلسَلة [الإنجليزية] جينوم فيروس سارس-كوف-2 مرات كثيرة، ونتج عن ذلك تحديد الآلاف من المتحورات. في تحليل لمنظمة الصحة العالمية بتاريخ يوليو 2020: كان ORF1ab أكثر جين متعرض للطفرات، يلية الجين S الخاص بالبروتين الشوكي. أكبر بروتين تعرض للطفرات داخل ORF1ab هو البروتياز المماثل للبابايين (nsp3)، وأكثر طفرة مغلطة متواترة كانت في بوليمراز الرنا المعتمد على الرنا.[13]
مراجع
- Hartenian؛ Nandakumar؛ Lari؛ Ly؛ Tucker؛ Glaunsinger (سبتمبر 2020)، "The molecular virology of coronaviruses"، Journal of Biological Chemistry، 295 (37): 12910–12934، doi:10.1074/jbc.REV120.013930.
- V’kovski؛ Kratzel؛ Steiner؛ Stalder؛ Thiel (مارس 2021)، "Coronavirus biology and replication: implications for SARS-CoV-2"، Nature Reviews Microbiology، 19 (3): 155–170، doi:10.1038/s41579-020-00468-6.
- Posthuma؛ te Velthuis؛ Snijder (أبريل 2017)، "Nidovirus RNA polymerases: Complex enzymes handling exceptional RNA genomes"، Virus Research، 234: 58–73، doi:10.1016/j.virusres.2017.01.023.
- Gulyaeva؛ Gorbalenya (يناير 2021)، "A nidovirus perspective on SARS-CoV-2"، Biochemical and Biophysical Research Communications، 538: 24–34، doi:10.1016/j.bbrc.2020.11.015.
- Wang؛ Jiang؛ Feng؛ Li؛ Guo؛ Sajid؛ Wu؛ Zhang؛ Ponty؛ Will, Sebastian؛ Liu, Feiyan؛ Yu, Xinghai؛ Li, Shaopeng؛ Liu, Qianyun؛ Yang, Xing-Lou؛ Guo, Ming؛ Li, Xingqiao؛ Chen, Mingzhou؛ Shi, Zheng-Li؛ Lan, Ke؛ Chen, Yu؛ Zhou, Yu (مايو 2021)، "The SARS-CoV-2 subgenome landscape and its novel regulatory features"، Molecular Cell، 81 (10): 2135–2147.e5، doi:10.1016/j.molcel.2021.02.036.
- Irigoyen؛ Firth؛ Jones؛ Chung؛ Siddell؛ Brierley (26 فبراير 2016)، "High-Resolution Analysis of Coronavirus Gene Expression by RNA Sequencing and Ribosome Profiling"، PLOS Pathogens، 12 (2): e1005473، doi:10.1371/journal.ppat.1005473.
- Finkel؛ Mizrahi؛ Nachshon؛ Weingarten-Gabbay؛ Morgenstern؛ Yahalom-Ronen؛ Tamir؛ Achdout؛ Stein؛ Israeli, Ofir؛ Beth-Din, Adi؛ Melamed, Sharon؛ Weiss, Shay؛ Israely, Tomer؛ Paran, Nir؛ Schwartz, Michal؛ Stern-Ginossar, Noam (07 يناير 2021)، "The coding capacity of SARS-CoV-2"، Nature، 589 (7840): 125–130، doi:10.1038/s41586-020-2739-1.
- Smith؛ Denison (05 ديسمبر 2013)، "Coronaviruses as DNA Wannabes: A New Model for the Regulation of RNA Virus Replication Fidelity"، PLoS Pathogens، 9 (12): e1003760، doi:10.1371/journal.ppat.1003760.
- Ogando؛ Ferron؛ Decroly؛ Canard؛ Posthuma؛ Snijder (07 أغسطس 2019)، "The Curious Case of the Nidovirus Exoribonuclease: Its Role in RNA Synthesis and Replication Fidelity"، Frontiers in Microbiology، 10: 1813، doi:10.3389/fmicb.2019.01813.
- Rohaim؛ El Naggar؛ Clayton؛ Munir (يناير 2021)، "Structural and functional insights into non-structural proteins of coronaviruses"، Microbial Pathogenesis، 150: 104641، doi:10.1016/j.micpath.2020.104641.
- Chen؛ Liu؛ Guo (أبريل 2020)، "Emerging coronaviruses: Genome structure, replication, and pathogenesis"، Journal of Medical Virology، 92 (4): 418–423، doi:10.1002/jmv.25681.
- Saberi؛ Gulyaeva؛ Brubacher؛ Newmark؛ Gorbalenya (01 نوفمبر 2018)، "A planarian nidovirus expands the limits of RNA genome size"، PLOS Pathogens، 14 (11): e1007314، doi:10.1371/journal.ppat.1007314.
- Koyama؛ Platt؛ Parida (01 يوليو 2020)، "Variant analysis of SARS-CoV-2 genomes"، Bulletin of the World Health Organization، 98 (7): 495–504، doi:10.2471/BLT.20.253591، PMC 7375210.
 بوابة علم الوراثة
بوابة علم الوراثة بوابة علم الأحياء الخلوي والجزيئي
بوابة علم الأحياء الخلوي والجزيئي بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية