فيروس كورونا
فيروسات كورونا أو الكورونافيرينات المستقيمة (الاسم العلمي: Orthocoronavirinae) هي عائلة فرعية من فيروسات الرنا مفردة السلسلة موجبة الدلالة المغلفة التي تصيب الثدييات والطيور،[6] وتنتمي إلى عائلة الفيروسات التاجية من رتبة الفيروسات العشية.[7] أول فيروس كورونا تم تحديده هو فيروس التهاب الشعب الهوائية المعدي الذي يصيب الدجاج في عقد 1920، واكتُشفت فيروسات كورونا التي تسبب الزكام للبشر في عقد 1960. سميت هذه الفيروسات باسم كورونا بسبب مظهرها تحت المجهر الإلكتروني الذي يبدو كهالة أو تاج بسبب بروتينات الشوكة.
اضغط هنا للاطلاع على كيفية قراءة التصنيف فيروس كورونا | |
|---|---|
 | |
 تمثيل لفيروس سارس-كوف-2 الذي سبب جائحة كوفيد 19.
الأزرق: غلاف ليبيد ثنائي الطبقة
أزرق فاتح: بروتين الشوكة السكري
أحمر: بروتين الغلاف
أخضر: بروتين الغشاء
برتقالي: جليكان تمثيل لفيروس سارس-كوف-2 الذي سبب جائحة كوفيد 19.
الأزرق: غلاف ليبيد ثنائي الطبقة أزرق فاتح: بروتين الشوكة السكري أحمر: بروتين الغلاف أخضر: بروتين الغشاء برتقالي: جليكان | |
| المرتبة التصنيفية | أسرة[1][2] |
| تصنيف الفيروسات | |
| المجموعة: | فيروس |
| النطاق: | ريبوفيريا |
| المملكة: | أرثوورنافيرات |
| الشعبة: | بيسوفيريكوتا |
| الطائفة: | بيسونيفريسيتس |
| الرتبة: | فيروسات عشية |
| الرتيبة: | كورنيدوفرينيات |
| الفصيلة: | فيروسات تاجية |
| الأسرة: | فيروس كورونا |
| الاسم العلمي | |
| Orthocoronavirinae[1][2] | |
| الأجناس[3] | |
| مرادفات [4] | |
| كورونافيرينات | |
يتراوح طول جينوم فيروسات كورونا بين 26 ألف و32 ألف نوكليوتيد وهو أطول الجينومات بين فيروسات الرنا،[8] ويشفر أربع بروتينات بنيوية: الشوكة (S)، الغلاف (E)، الغشاء (M)، القفيصة النووية (N) وفي بعض الفيروسات بروتين خامس هو إستراز الراصة الدموية (HE) كما يشفر جين الريبلكاز والعديد من البروتينات الملحقة. فضلا على الحفاظ على بنية الفيروس، تملك هذه البروتينات وظائف متعددة مثل تعزيز الإصابة ومقاومة الاستجابات المناعية للمضيف. يقوم بروتين الشوكة بالارتباط بمستقبلاتٍ على سطح الخلية المضيفة ودمج الغلاف الفيروسي مع الغشاء الخلوي الخاص بالخلية المضيفة للدخول إليها. بعد الدخول إلى السيتوبلازم تعمل بعض البروتينات اللابنيوية على إعادة تنظيم الغشاء الداخلي لإنشاء عضيات التضاعف التي يحدث فيها النسخ والتضاعف.[9] أثناء النسخ، يحدث تبديل للقالب لإنشاء جزيئات رنا جينومية فرعية بأطوال مختلفة تشفر واحدا أو عدة بروتينات بنيوية وهي ميزة تتميز بها الفيروسات العشية، قد يكون تبديل القالب هو آلية التأشيب الجيني بين مختلف فيروسات كورونا التي تسمح لها بتوليد سلالات جديدة أو الانتقال عبر الأنواع. يخضع جين الريبليكاز لانزياح إطار ريبوسومي -1 أثناء الترجمة لنسخ عديد البروتين الكبير 1ab الذي يخضع لقص ذاتي بواسطة وظيفة البروتياز الخاصة بالبروتينين اللابنيويين 3 و5 وينتج عنه 16 بروتينا لا بنيويا لها وظائف متنوعة أهمها البروتين اللابنيوي 12 الذي يملك وظيفة بوليميراز الرنا المعتمد على الرنا ويقوم بعملية النسخ، والبروتين اللابنيوي 14 الذي يملك وظيفة تصحيح تزيد من دقة النسخ، وربما يكون السبب في قدرة فيروسات كورونا على امتلاك جينوم بهذا الطول. الإصابة بفيروس كورونا تسبب استجابات خلوية عديدة مثل استجابة إجهاد الشبكية الإندوبلازمية التي تؤدي إلى إفراز السيتوكينات وحدوث الاستماتة، وتؤثر على العديد من مسارات التأشير وتثير استجابات مناعية، لكن يملك الفيروس بعض الآليات لمقاومة هذه الاستجابات وتثبيط مسارات تخليق الإنترفيرونات.[10]
تنقسم عائلة فيروسات كورونا الفرعية إلى أربع أجناس: فيروس كورونا ألفا، فيروس كورونا بيتا، فيروس كورونا غاما، فيروس كورونا دلتا وتصيب أجناس ألفا وبيتا الثدييات فقط وسلفهما المشترك يمكن أن يكون فيروسا خفاشيا، أما غاما ودلتا فيصيب معظمها الطيور بشكل رئيسي مع أنواع قليلة تصيب الثدييات ويُفترض أن سلفهما المشترك فيروس يصيب الطيور. ينتمي فيروسا سارس-كوف وسارس-كوف-2 إلى نوع الفيروسات المرتبطة بالسارس الذي ينتمي إلى الجنس الفرعي الفيروسات الساربيكوية الذي ينتمي إلى جنس فيروسات بيتا.[11] يشمل الجنس الفرعي فيروس إمبيكو الفيروسات التي تصيب الفئران ومن بينها فيروس التهاب الكبد الفأري وهو فيروس نموذجي في الأبحاث وقد تم توضيح العديد من الآليات الجزيئة لفيروسات كورونا عبر دراسته.
توجد سبع فيروسات تصيب البشر، 4 منها (فيروس كورونا البشري OC43، فيروس كورونا البشري HKU1، فيروس كورونا البشري 229E وفيروس كورونا البشري NL63) يمكن أن تسبب الزكام، والثلاثة الأخرى سببت تفشيات أمراض خطيرة حول العالم وهي فيروس كورونا المرتبط بالمتلازمة التنفسية الحادة الشديدة النوع 1 (سارس-كوف)، فيروس كورونا المرتبط بمتلازمة الشرق الأوسط التنفسية (ميرس-كوف) وفيروس كورونا المرتبط بالمتلازمة التنفسية الحادة الشديدة النوع 2 (سارس-كوف-2). فضلا عن ذلك، توجد العديد من فيروسات كورونا التي يمكن أن تصيب الماشية والدواجن والحيوانات الأليفة وحيوانات المختبر والحيوانات البرية، مثل فيروس كورونا الطيري الذي يصيب الدجاج والعديد من فيروسات كورونا الخنزيرية التي تصيب الخنازير وفيروس كورونا الكلبي وفيروس كورونا القططي اللذان يصيبان الكلاب والقطط، وفيروس كورونا مورين الذي يصيب الفئران والجرذان المختبرية.
التأثيل
اشتُق اسم "coronavirus" (عربيًا: فيروس كورونا. اختصارًا CoV) من الكلمة (اللاتينية corona) وتعني «التاج» أو «الإكليل» المأخوذة بدورها من الكلمة (الإغريقية κορώνη) وتعني إكليل زهور أو الإكليل.[12][13] صيغ المصطلح بواسطة جون ألميدا وديفيد تيريل اللذان كانا أول من لاحظ ودرس فيروسات كورونا البشرية.[14] استُعمل المصطلح لأول مرة في منشور سنة 1968 بواسطة مجموعة من علماء الفيروسات غير الرسميين في مجلة نيتشر لتسمية عائلة الفيروسات الجديدة.[15] يُشير الاسم إلى المظهر المميز للفيريونات (الشكل المُعدي للفيروس) والذي يظهر عبر المجهر الإلكتروني، حيث تمتلك خُملًا/زغاباتٍ من البروزات السطحية البصلية الكبيرة، مما يُظهرها على شكل تاج الملك أو الهالة الشمسية.[14][15] يحدث هذا التشكُل عبر قسيماتٍ فولفية للشوكة الفيروسية (S)، وهي بروتيناتٌ تملأ سطح الفيروس وتحدد انتحاء المضيف.[16]
قُبل الاسم العلمي "Coronavirus" بواسطة اللجنة الدولية لتسمية الفيروسات التي أعيدت تسميتها لاحقا إلى اللجنة الدولية لتصنيف الفيروسات (ICTV) في عام 1971.[17] ومع ازدياد عدد الأنواع تم تقسيم هذه العائلة الفرعية إلى أربع الجنس هي: فيروس كورونا ألفا، فيروس كورونا بيتا، فيروس كورونا غاما وفيروس كورونا دلتا عام 2009.[18] يُستخدم الاسم الشائع فيروس كورونا للإشارة إلى أي عضو في العائلة الفرعية "Orthocoronavirinae" من عائلة الفيروسات التاجية.[7] واعتبارا من 2020، يوجد 45 نوعا من فيروسات كورونا معترف بها رسميا.[19]
في اللغة العربية، تُعتبر تسمية فيروس كورونا(1) أكثر شيوعًا من باقي التسميات الأُخرى، ولكن التسميات الأُخرى أكثر دقةٍ في الوصف، حيثُ يُسمى: الفيروس التاجي،[ِ 1] فيروس الهالة،[ِ 1] الفيروسة المُكَلَّلَة[ِ 2][ِ 3] (أو الفيروس المكلل)،[ِ 1] الحُمّة التاجيّة[ِ 4][ِ 5][ِ 1] الحُمَة الإكليلية،[ِ 6] الحمة التاجية المكللة.[ِ 1]
تاريخ الأبحاث

ظهرت أولى التقارير عن إصابة حيواناتٍ بفيروس كورونا في أواخر العقد 1920، عندما تفشت عدوى تنفسية حادة بين دجاجٍ مستأنس في أمريكا الشمالية.[21] حيث كتب آرثر شالك وم.س. هاون عام 1931 أول تقرير مفصل يصف عدوى تنفسية جديدة بين الدجاج في داكوتا الشمالية. اتسمت إصابة الكتاكيت حديثة الولادة باللهاث والفتور مع معدلات نفوق عالية بين 40-90٪.[22] عزل ليلاند ديفيد وكارل ألفريد الفيروس الذي تسبب في الإصابة بالعدوى عام 1933،[23] وعُرف آنذاك باسم فيروس التهاب الشعب الهوائية المعدي (IBV). تمكن تشارلز د.هدسون وفريد روبرت بيوديت من زرع الفيروس لأول مرة عام 1937،[24] وأصبحت تلك العينة تُعرف باسم سلالة بيوديت. في أواخر العقد 1940، اكتُشف نوعان آخران من فيروسات كورونا الحيوانية: JHM الذي يتسبب في مرض الدماغ (التهاب الدماغ عند الفئران) وفيروس التهاب الكبد الفأري (MHV) الذي يسبب التهاب الكبد في الفئران.[25] لم يتم في ذلك الوقت إدراك أن هذه الفيروسات الثلاثة المختلفة كانت لها صلة ببعضها.[17][26]
اكتُشفت فيروسات كورونا البشرية في العقد 1960[27][28] باستخدام طريقتين مختلفتين في المملكة المتحدة والولايات المتحدة.[29] جمع إ.س. كيندال، ومالكولم بينو، وديفيد تيريل في عام 1961 أثناء عملهم في وحدة الزكام التابعة لمجلس البحوث الطبية البريطاني فيروسا فريدا يسبب الزكام سُمي بـB814.[30][31][32] ولم يتمكنوا من زراعته باستخدام التقنيات المعيارية التي نجحت سابقا في زراعة الفيروسات الأنفية والفيروسات الغدية وغيرها من الفيروسات التي تسبب الزكام. في عام 1965، نجح تيريل وبينو في زراعة الفيروس الجديد عبر تمريره بشكل متسلسل في مزارع أعضاء لقصبات هوائية جنينية بشرية.[33] وهي طريقة زراعة جديدة أحضرها برتيل هورن إلى ذلك المختبر.[34] تسبَّبَ الفيروس المعزول عند تلقيح المتطوعين به عن طريق الأنف في الإصابة بالزكام وتم تثبيطه بواسطة الإيثر وهو ما يشير إلى امتلاكه لغلاف دهني.[30][35] عزلت دوروثي هامري[36] وجون بروكنو في جامعة شيكاغو فيروس زكام جديد من طلاب طبٍ في عام 1962، وقاما بتنميته في مزرعة نسيج الكلى وسمياه 229E. سبّب الفيروس الجديد زكاما لدى المتطوعين، وعلى غرار B814 ، تم تثبيطه بواسطة الإيثر.[37]

قارنت عالمة الفيروسات الاسكتلندية جون ألميدا من مستشفى سانت توماس في لندن -بالتعاون مع تيريل- بُنى الفيروسات IBV وB814 و229E في عام 1967.[38][39] باستخدام المجهر الإلكتروني، تبين أن الفيروسات الثلاثة لها صلة مورفولوجيا من خلال شكلها العام وشوكاتها المميزة التي تشبه النفل.[20] تمكنت مجموعة بحثية في معاهد الصحة الوطنية الأمريكية في نفس العام من عزل عضو آخر من هذه المجموعة الجديدة من الفيروسات باستخدام زراعة الأعضاء وقامت بتسمية إحدى العينات OC43 (OC اختصارٌ لـorgan culture).[40] على غرار B814 و229E وIBV، كان لفيروس الزكام الجديد OC43 شوكات مميزة شبيهة بالنفل عندما لوحظ بالمجهر الإلكتروني.[41][42]
سرعان ما تبين أن فيروسات الزكام الجديدة المماثلة لـ IBV لها صلة مورفولوجية بفيروس التهاب الكبد الفأري،[25] وسُميت هذه المجموعة الجديدة من الفيروسات باسم فيروسات كورونا وذلك لمظهرها المورفولوجي المميز.[15] استمرت دراسة فيروس كورونا البشري 229E وفيروس كورونا البشري OC43 خلال العقود اللاحقة،[43][44] أما سلالة فيروس كورونا B814 فقد فُقدت ومن غير المعروف أيٌّ من فيروسات كورونا الحالية هو فيروس B814.[45] منذ ذلك الحين، تم تحديد عدة فيروسات كورونا بشرية أخرى ومنها: سارس-كوف في عام 2003، وكوف-NL63 البشري في عام 2003، وكوف-HKU1 البشري في عام 2004، وميرس-كوف في عام 2013، وسارس-كوف-2 في عام 2019.[46] كما تم تحديد عدد كبير من فيروسات كورونا الحيوانية منذ الستينيات.[47]
علم الفيروسات
الشكل والبنية

فيروسات كورونا هي جسيمات كبيرة، كروية الشكل تقريبا وذات نتوءات (شوكات) سطحية فريدة.[48] حجمها متغير بدرجة كبيرة ومتوسط قطرها يترواح بين 80 إلى 120 نانومتر، ويصل قطر الأحجام المختلفة إلى حد أدناه 50 ناتومتر وأقصاه 200 نانومتر.[49] يبلغ متوسط الكتلة الجزيئية الكلية 40000 كيلو دالتون. وهي محاطة بغلاف مدمجة فيه عدة جزيئات بروتينية.[50] يحمي غلاف الليبيد ثنائي الطبقة وبروتينات الغشاء والقفيصة النووية الفيروس عندما يكون خارج الخلية المضيفة.[51]
يتكون غلاف الفيروس من ليبيد ثنائي الطبقة تكون البروتينات البنيوية: الغشاء (M) والغلاف (E) والشوكة (S) مدمجة به.[52] نسبة تواجد هذه البروتينات في غشاء اللبيد ثنائي الطبقة هي حوالي 1 بروتين غلاف لكل 20 بروتين شوكة لكل 300 بروتين غشاء.[53] بروتينا الغلاف والغشاء هما المسؤولان على تحديد شكل الفيروس حيث يتآثران مع الليبيد ثنائي الطبقة لإحداث انحناءات في الغشاء وإنشاء فيروس شكله كروي تقريبا وتحديد حجمه.[54][55] بروتين الشوكة ضروري من أجل التآثر مع مستقبلات على سطح الخلايا المضيفة من أجل الدخول، لكن فيروس كورونا البشري NL63 متميز بأن بروتين الغشاء هو من لديه نطاق الارتباط بمستقبلات الخلية المضيفة وليس بروتين الشوكة.[56] يبلغ قطر الغلاف حوالي 85 نانومتر. يظهر غلاف الفيروس في الصور المجهرية الإلكترونية كزوج مميز من أغلفة كثافة الإلكترون (غلافان معتمان نسبيا مقارنة بشعاع الإلكترون المستخدم لمسح جسيم الفيروس).[55][57]

الجينوم
تملك فيروسات كورونا جينوم رنا مفرد السلسلة موجب الدلالة، يتراوح طوله بين 26 ألف إلى 32 ألف نوكليوتيد.[8] وهو من أكبر الجينومات بين فيروسات الرنا. يحتوي الجينوم على قبعة 5' ممثيلة في النهاية 5' وذيل عديد أدينين في النهاية 3' طوله بين 30-60 أدينين.[55]
تنظيم الجينات في جينوم فيروسات كورونا عموما هو: المنطقة غير المترجمة 5' - الريبلكاز - الشوكة - الغلاف - الغشاء - القفيصة النووية - المنطقة غير مترجمة 3' - ذيل عديد الأدينين. يشغل إطارا القراءة المقتوحان 1a و1b ثلثي الجينوم ويشفران عديد بروتين الريبلكاز 1ab الذي يقوم بقص نفسه إلى 16 برووتينا لا بنيويا.[55] تشفر إطارات القراءة المفتوحة التالية البروتينات البنيوية الرئيسية الأربعة: الشوكة والغلاف والغشاء والقفيصة النووية، وفي بعض الفيروسات يوجد بروتين بنيوي خامس هو إستراز الراصة الدموية (HE). تتخلل هذه البروتينات البنيوية بروتينات ملحقة يختلف عددها وموضعها ووظيفتها وطولها حسب الفيروس،[59] ورغم أن معظمها غير لازم من أجل تضاعف الفيروس أو إصابته للخلية إلا أنها مسؤولة على الوظيفة الإمراضية للفيروس وتلعب أدوارا في مقاومة الاستجابة المناعية للمضيف.[60]
المنطقة غير المترجمة 5' و3'
المنطقة غير المترجمة 5' واختصارا مغم 5' [ملاحظة 1] (5′ UTR) والمنطقة غير المترجمة 3' (مغم 3') هما منطقتان من جينوم فيروس كورونا لا تتم ترجمتهما إلى بروتينات لكنهما ضروريتان ولازمتان من أجل تنسخ الفيروس وتضاعفه، وتحتويان على العديد من الحلقات جذعية التي تسلسلاتها وبُناها الثانوية منحفظة تطوريًا.[61]
المنطقة غير المترجمة 5'

يبلغ طول المنطقة غير المترجمة 5' الخاصة بفيروس كورونا حوالي 200-300 نوكليوتيد،[62] وتحتوي بشكل عام على الحلقة الجذعية 1 (SL1) والحلقة الجذعية 2 (SL2) والحلقة الجذعية 4 (SL4)، ويملك فيروس كورونا البقري، وفيروس كورونا المرتبط بالسارس وبعض فيروسات كورونا غاما ودلتا أيضا حلقة جذعية 3 (SL3) بين الحلقة الجذعية 2 والحلقة الجذعية 4.[63][64] فضلا عن هيئة التطوي الكاملة، يمكن أن تتواجد SL1 كذلك في توازن ديناميكي مع هيئات تطوي عالية الطاقة غير كاملة التطوي في الجذع، تعمل بمساعدة بروتينات مفترضة على التآثر مع مناطق أخرى من الجينوم على مسافات طويلة والمشاركة في تنسخ الفيروس وتضاعفه.[62][65] تسلسل الحلقة SL2 هو التسلسل الأكثر انحفاظا بين تسلسلات الحلقات الجذعية في مغم 5'، ويمكن أيضًا أن يكون له صلة بتنسخ الفيروس وتضاعفه.[62] تحتوي الحلقة SL3 على التسلسل المنظم للنسخ الطليعي (TRS-L) في منطقة الحلقة وهو أساسي وحاسم لتخليق الرنا الجينومي الفرعي أثناء تضاعف الفيروس، لكن هذه الحلقة غير مستقرة في فيروسات مثل فيروس كورونا الفأري.[66] تسلسل الحلقة SL4 طويل نسبيًا ويمكن تقسيم الحلقة الجذعية إلى جزأين: 4a و4b مفصولين بانتباج في المنتصف. يوجد إطار قراءة مفتوح قصير في SL4 في العديد من فيروسات كورونا هو إطار القراءة المفتوح عكس الاتجاه، الذي يمكنه تثبيط ترجمة الريبليكاز 1ab، تُظهر النتائج التجريبية أن SL4 متسامحة للغاية مع الطفرات وتتحملها وحتى استبدال SL4 كاملة بتسلسل مختلف لا يؤثر على التضاعف (ولكن إذا تم حذفها لا يمكن أن يتضاعف الفيروس)، لذلك افتُرض أن وظيفتها الرئيسية هي تحديد الرنا الجينومي الفرعي الذي يتم نسخه،[62][67] وبنية جذعها أكثر مرونة وهو ما يسمح لها بالتآثر مع الرنا الجينومي الفرعي أثناء النسخ والمساعدة في تبديل القالب.[62][68]
علاوة على هذه الحلقات، تملك العديد من فيروسات كورونا حلقة خامسة SL5 مع الاتجاه بالنسبة للحلقة الرابعة. تتجاوز الحلقة 5 في فيروسات ألفا وبيتا المنطقة غير المترجمة 5' وتتداخل مع ORF1a وتسلسلها أكثر انحفاظا بين فيروسات ألفا. تتكون الحلقة 5 في فيروسات ألفا وبيتا من ثلاث حلقات صغيرة SL5a وSL5b وSL5c جميعها تحتوي على التسلسل 5'-UUCCG(U/C)-3 في أعلاها. إشارة التغليف ضرورية من أجل تجميع بعض فيروسات كورونا.[69] تملك فيروسات غاما الحلقة SL5 كذلك لكن بنييتها مختلفة قليلا عن تلك الخاصة بفيروسات ألفا وبيتا،[64] ويمكن أن يكون لها أيضا دور في تضاعف الرنا الفيروسي.[62][70]
المنطقة غير المترجمة 3'

يبلغ طول المنطقة غير المترجمة 3' الخاصة بفيروس كورونا حوالي 300-500 نوكليوتيد (دون احتساب ذيل عديد الأدينين).[62] النهاية 5' للمنطقة غير المترجمة 3' الخاصة بفيروسات بيتا (مباشرة بعد نهاية جين القفيصة النووية) عبارة عن حلقة جذعية منتبجة (BSL) ضرورية لتضاعف الفيروس طولها حوالي 70 نوكليوتيد، وقد تحتوي بعض فيروسات كورونا غاما على هذه البنية كذلك،[71] ولكنها غير متواجدة في فيروسات ألفا.[70] توجد بنية عقدة كاذبة طولها 54 نوكليوتيد مع الاتجاه بالنسبة للحلقة المنتبجة في فيروسات كورونا ألفا وبيتا، والتي قد تكون مطلوبة لتضاعف الفيروس. ترتبط حلقةٌ جذعية ثانية خاصة بالعقدة الكاذبة (PK-SL2) بتسلسلٍ في جذع الحلقة المنتبجة لتشكيل العقدة الكاذبة وهذا يعني أن الحلقة المنتبجة والعقدة الكاذبة متداخلتان في التسلسل ولا يمكنهما التطوي إلى هيئاتهما في آن واحد (صورة) وأنهما يعملان كمفتاح جزيئي.[72] معدل تواجد الحلقة المنتبجة والعقدة الكاذبة بشكل مستقل أو تشكيلهما لهيئة مشتركة يكون بتوازن ديناميكي من أجل تنظيم بدء نسخ سلسلة الرنا الفيروسية سالبة الدلالة. فحين لا تشكل العقدة الكاذبة هيئة مشتركة مع الحلقة المنتبجة يُفترض أنها تترابط مع تسلسل النهاية 3' لتشكيل جذع يعمل كموقع لتجميع البروتينات اللابنيوية التي تعمل على تخليق السلسلة الجينومية السالبة، وعند الانتهاء من النسخ وانفصال تسلسل النهاية 3' تعود الحلقة الكاذبة للتشكل.[62][73]
مع الاتجاه بالنسبة للعقدة الكاذبة تتواجد عقدة جذعية أخرى طويلة ومتفرعة غير ضرورية لتضاعف الفيروس[74] تسمى المنطقة فائقة التغير (HVR) بسبب تغير تسلسلها الكبير بين الفيروسات. تحتوي هذه الحلقة على تسلسل ثماني محفوظ 5′-GGAAGAGC-3′[75] وظيفته غير واضحة لحد الآن.[60]
البروتينات
يمكن تقسيم بروتينات فيروسات كورونا إلى قسمين: البروتينات البنيوية والبروتينات اللابنيوية، وتشمل البروتينات البنيوية:الغشاء (M) والغلاف (E) والشوكة (S) وفي بعض الفيروسات إستراز الراصة الدموية (HE) وتدخل جميعها في تركيب الفيروس. أما البروتينات اللابنيوية فهي 16 بروتينا تنتج من القص الذاتي لعديد البروتين 1ab الذي يُشفره جين الريبليكاز (ORF1ab) ولها أدوار مختلفة أهمها نسخ ومضاعفة الفيروس، بالإضافة إلى البروتينات البنيوية واللابنيوية، تملك العديد من فيروسات كورونا بروتينات ملحقة ذات وظائف متنوعة.[55]
بروتين الغشاء
بروتين الغشاء هو البروتين الرئيسي للغلاف الفيروسي والأكثر توافرا فيه من بين البروتينات البنيوية الأخرى، وهو بروتين غشائي من النوع الثالث طوله 218 إلى 263 حمض أميني ويشكل طبقة سمكها 7.8 نانومتر.[50] ويملك نطاق نهاية أمنية قصيرخارجي وثلاث نطاقات عبر غشائية ونطاق نهاية كربوكسيلية داخلي يشكل مطرسا مثل الشبيكة يزيد من ثخانة ومتانة الغلاف. حسب الفيروس، يمكن أن تتم غلكزة النطاق الأميني الخارجي لهذا البروتين بربط الجليكانات في N أو في O. تلعب بروتينات الغشاء دورا مفتاحيا في تجميع الفيروس، فهي تغير بنيتها لتعديل انحناء الغشاء وتشكيل الغلاف الفيروسي،[76] وتتآثر مع البروتينات البنيوية الأخرى لتنسيق تجميع الفيروس وتبرعمه.[77]
تستخدم جميع فيروسات كورونا بروتين الشوكة للارتباط بالمستقبلات على سطح الخلية المضيفة، لكن أظهرت دراسات أن بروتين الغشاء الخاص بفيروس كورونا البشري NL63 يمكنه الارتباط بالجليكان البروتيني كبريتات الهيباران المتواجد على سطح الخلايا المضيفة، لكن يبقى بروتين الشوكة مطلوبا من أجل دخولٍ ناجح.[78]
بروتين الغلاف
بروتين الغلاف هو بروتين بنيوي ثانوي يختلف كثيرا بين أنواع فيروسات كورونا.[49] يوجد حوالي 20 بروتين غلاف فقط في فيروس كورونا،[53] ويتراوح طوله بين 76 إلى 109 حمض أميني ووزنه الجزيئي بين 8.4 و12 كيلو دالتون. بروتين الغلاف هو الأصغر بين البروتينات البنيوية الأربعة وهو بروتين غشائي مدمج يملك نطاقين: نطاق عبر غشائي ونطاق نهاية كربوكسيلية خارجي، النطاق عبر الغشائي هو لولب ألفا مفرد يمكن أن يتجمع إلى خماسي قسيمات -كما هو الحال في سارس-كوف- ويشكل قنوات أيونية تسمى فيروبورينات تسمح للأيونات (بشكل أساسي أيونات الهيدروجين) بالمرور. وظيفة هذه القناة الأيونية مازالت قيد الدراسة، لكن قد يكون لها دور في عملية تحرير الفيروس من الخلايا.[77] فضلا عن ذلك، تشارك بروتينات الغلاف أيضًا في تجميع الفيروس وتنقله داخل الخلية وتبرعمه. أظهرت الدراسات أن فيروس التهاب الكبد الفأري الذي يفتقر إلى بروتينات الغلاف لا يزال بإمكانه إصابة الخلايا. ولوحظ انخفاض في إمراضية فيروس السارس،[ملاحظة 2] أما ميرس-كوف الذي يفتقر إلى بروتين الغلاف فلم يكن قادرا على إصابة الخلايا.[77] بالإضافة إلى ذلك، قد تثبط بروتينات الغلاف لبعض فيروسات كورونا الاستجابة المناعية للخلايا المضيفة.[79]
بروتين الشوكة

بروتينات الشوكة هي السمة الأكثر تمييزا لفيروسات كورونا وهي المسؤولة عن مظهر السطح الذي يشبه الهالة أو التاج. يحتوي جسيم فيروس كورونا في المتوسط على 74 شوكة،[80] ويبلغ طول كل شوكة حوالي 20 نانومتر وهي عبارة عن مثلوث من بروتين الشوكة. يتكون بروتين الشوكة بدوره من وحدتين فرعيتين S1 وS2. بروتين الشوكة الثلاثي المتجانس هو بروتين دمج غشاء من الصنف الأول يتوسط الارتباط بالمستقبلات واندماج الغشاء بين الفيروس والخلية المضيفة. تمثل الوحدة الفرعية S1 رأس الشوكة وتحتوي على نطاق الارتباط بالمستقبل، وتمثل الوحدة الفرعية S2 الجذع الذي يثبت الشوكة في الغلاف الفيروسي وتقوم عند تنشيطها بالبروتياز بدمج الغشائين. تظل الوحدتان الفرعيتان مرتبطتين بشكل غير تساهمي وهما مكشوفتان على سطح الفيروس حتى ترتبطان بغشاء الخلية المضيفة.[50] ينقسم مركب الوحدات الفرعية إلى وحدات فرعية منفردة عندما يرتبط الفيروس بالخلية المضيفة ويندمج بها بفضل وظيفة البروتيازات مثل: عائلة الكاتبسينات، وبروتياز السيرين عبر الغشائي 2 (TMPRSS2) الخاصة بالخلية المضيفة.[81]
نطاقات الوحدة S1 هي أهم مكونات الشوكة في إحداث الإصابة، وهي أيضا المكونات الأكثر تنوعا بين الفيروسات لأنها مسؤولة على الانتحاء الخلوي. تمتلك الوحدة الفرعية S1 نطاقين رئيسيين هما: نطاق النهاية الأمينية (S1-NTD) ونطاق النهاية الكربوكسيلية (S1-CTD)، وكلاهما يعمل كنطاق ارتباط بالمستقبل. تتعرف نطاقات النهاية الأمينية على السكريات الموجودة على سطح الخلية المضيفة وترتبط بها،[ملاحظة 3] بينما تتعرف نطاقات النهاية الكربوكسيلية على المستقبلات البروتينية المختلفة مثل الإنزيم المحول للأنجيوتنسين 2، أمينوببتيداز الألانين والببتيداز نازع الببتيديل 4 وترتبط بها.[50]
فضلا عن النطاقات المستخدمة عادة، يمكن لجزيئات الغليكوزيل الموجودة على بروتين الشوكة أن ترتبط بالليكتينات الموجودة على سطح الخلية. على سبيل المثال: رغم أن سارس-كوف عادة ما يستخدم المسستقبل ACE2 إلا أن غليكوزيل بروتين اللشوكة يمكن أن يرتبط بـ CLEC4M وDC-SIGN وغيرها من الليكتينات الأخرى كمستقبلات بديلة.[82][83]
قامت عدة دراسات بتحليل بنية بروتين الشوكة الخاص بفيروس كورونا الفأري ووجدت أن تطوي نطاق النهاية الأمينية لـS1 يشبه تطوي الجالكتين في الخلايا الحيوانية، ووفقا لذلك اقتُرحت فرضية أن أصل النطاق الأميني لبروتين الشوكة يرجع إلى خلية حيوانية مضيفة، أي أن أول فيروس كورونا حصل على الجين من الليكتين الخاص بالخلية المضيفة والذي يستطيع الارتباط بالجليكانات الموجودة على سطح الخلية واستخدمها كمستقبل لإصابة الخلايا، ولكن لاحقا تحور S1 الخاص بالعديد من فيروسات كورونا وتطور ليصبح قادرا على الارتباط بمستقبلات البروتين في الخلية المضيفة. على سبيل المثال: اتخذ النطاق الأميني لفيروس كورونا الفأري بنية جديدة واكتسب القدرة على الارتباط بـ CEACAM1 وهو ما زاد من قدرة الفيروس على الارتباط بخلايا الفئران بشكل كبير. ونظرا لعدم الحاجة إلى الارتباط بالسكريات فإنه فقد تدريجيا وظيفة الليكتين وزيادة على ذلك فقد وظيفة إستراز الراصة الدموية. في المقابل، فيروس كورونا البقري وفيروس كورونا البشري OC43 وغيرها مازالت تستخدم السكريات كمستقبلات، لذلك تحتفظ نطاقاتها الأمينية بوظيفة الليكتين.[84] في عملية تطور S1 الخاص بفيروس كورونا، يواجه النطاق الكربوكسيلي ضغطا تطوريا أكبر، لذلك معدل التطور فيه أسرع من معدل التطور في النطاق الأميني. عندما يتحور النطاق الكربوكسيلي ويحاول الارتباط بمستقبلات جديدة، يمكن استخدام النطاق الأميني الذي يرتبط بالجليكوزيل كآلية تأمين لضمان إمكانية ارتباط الفيروس بالخلايا.[85]
القص

يوجد موقعان في بروتين الشوكة يحدث فيهما القص من أجل أن يتمكن فيروس كورونا من إصابة الخلايا وهما: موقع S1/S2 عند حدود المنطقتين وموقع S2'. بعد القص في الموقع الأول تفقد المنطقتين S1 وS2 ترابطهما التساهمي، لكنهما تبقيان مرتبطتين ببعضهما عبر قوى بين جزيئية أخرى. عند الارتباط بالمستقبل تخضع بنية S2 لتغير هيئي يسمح بقص ثانٍ في موقع S2' يتحرر بواسطته ببتيد الدمج الذي يشرع في عملية دمج الغشائين الفيروسي والخلوي، وهو ما يسمح لجينوم الرنا الفيروسي بالدخول إلى السيتولازم.[61]
تختلف فيروسات كورونا في مرحلة دورة حياة الفيروس التي يحدث فيها القص وخاصة في موقع S1/S2، إذ يُقص بروتين الشوكة في بعض فيروسات كورونا بواسطة بروتيازات داخل خلوية قبل أن يغادر الخلية. على سبيل المثال: يوجد تسلسل لدى فيروس كورونا الفأري A59،[86] وميرس-كوف [ملاحظة 4] وسارس-كوف-2 بين S1 وS2 يمكن لبروتياز الفيورين أن يقوم بالقص فيه داخل الخلايا. في حين أن فيروسات أخرى تُقص بواسطة بروتيازات خارج خلوية مثل الإيلاستاز بعد الإخراج الخلوي.[85] بعد الارتباط بالمستقبلات الخلوية تغير المنطقة S2 هيئتها بسرعة ثم يتم قصها في موقع S2' بواسطة بروتيازات أخرى على سطح الخلايا مثل بروتياز السيرين عبر الغشائي 2 بالنسبة لسارس-كوف-2، وبعد القص يمكن للفيروس أن يندمج مع غشاء الخلية ويدخل إلى السيتوبلازم.[10][89][ملاحظة 5]. الفيروسات التي لا تحتوي على مواقع قص للفيورين مثل سارس-كوف لا يتم قصها في S1/S1 وS2' سوى بواسطة بروتيازات على سطح الخلية أو في الجسيم الحال بعد إصابة الخلايا.[61]
بروتين القفيصة النووية
توجد القفيصة النووية داخل الغلاف الفيروسي وتتكون من العديد من بروتينات القفيصة النووية المرتبطة بالرنا. يرتبط بروتين القفيصة النووية بجينوم الرنا مفرد السلسلة موجب الدلالة على هيئة خرز على خيط مستمرة، كما ترتبط بروتينات القفيصة النووية ببعضها البعض مشكلة خيوط قفيصة نووية عرضها من 10 إلى 15 نانومتر وطولها مئات النانومترات.[55][91][92] بروتينات القفيصة هي بروتينات فوسفورية وزنها الجزيئي من 45 إلى 50 ألف كيلو دالتون وطولها في سارس-كوف 422 حمض أميني وفي سارس-كوف-2 419 حمض أميني.[93][94] يتكون بروتين القفيصة من 5 نطاقات: نطاق أميني يرتبط بالرنا ونطاق كربوكسيلي يشكل مثنويات متجانسة، يفصلهما نطاق مركزي مضطرب جوهريا عالي الانحفاظ وغني بالسيرين، وعلى جانبيهما نطاقان آخران مضطربان جوهريا.[95] بروتين القفيصة متعدد الوظائف وهو مولد كبير للمناعة حيث وُجدت أجسام مضادة له لدى المرضى المتعافين من مرضي السارس وكوفيد 19،[96] وفضلا عن وظيفته الرئيسية في تعبئة الجينوم الفيروسي والمساهمة في عملية التجميع، يساهم بروتين القفيصة في عملية تبديل القالب لنسخ الرنا الجينومي الفرعي،[97] ويدخل إلى النواة ويسبب توقف دورة الخلية في طور التركيب عبر تآثرات مع مركب الكيناز المعتمد على السيكلين[98] كما أنه يثبط استجابات الإنترفيرون في كل من سارس-كوف، ميرس-كوف، وسارس-كوف-2 بطرق متنوعة.
إستراز الراصة الدموية
فضلا عن بروتينات الغشاء والغلاف والشوكة، يوجد بروتين آخر على غلاف فيروسات كورونا من جنس بيتا، الفرع A (مثل فيروس كورونا الفأري وفيروس كورونا البشري HKU1) هو إستراز الراصة الدموية (HE).[47] هذا البروتين مثنوي ويتكون من وحدتين فرعيتين متماثلتين وطوله حوالي 400 حمض أميني، وهو بارز على سطح الفيروس مثل بروتين الشوكة لكنه أقصر منه وطوله حوالي 5 إلى 7 نانومتر، ووظيفته المساعدة في ارتباط الفيروس بالمستقبلات على الخلية المضيفة.[99] بالإضافة إلى فيروسات كورونا، تحتوي فيروسات الإنفلونزا وفيروسات تورو على هذا البروتين كذلك، مما يشير إلى إمكانية حدوث تأشيب بين هذه الفيروسات.[100] يتكون بروتين إستراز الراصة الدموية من نطاقين رئيسيين: نطاق ليكتين ونطاق إستراز. يمكن أن يرتبط كل من نطاق الليكتين وبروتين الشوكة بمستقبلات حمض السياليك على سطح الخلايا المضيفة، يساعد نطاق الإستراز في تحرير الفيروس من الخلية ومغادرته لها عبر قص أستيل من الوضعية 9 من جزيء حمض السياليك ( 9-O-Ac-Neu5Ac) النهائي. بعد انتقال فيروس كورونا البشري OC43 (الذي ينتمي إلى النوع فيروس كورونا بيتا 1) إلى مضيف بشري من فيروس كورونا البقري، فقد إستراز الراصة الدموية قدرة الارتباط بحمض السياليك تدريجيا، وأصبح بروتين الشوكة فقط هو الذي يربط بحمض السياليك، وتأثر نشاط نطاق الإستراز كذلك. خضع إستراز الراصة الدموية لفيروس كورونا البشري HKU1 لتطور تقاربي أيضا وحدثت له تغيرات مماثلة. قد تكون هذه التغيرات مرتبطة بتكيف الفيروس مع مستقبلات حمض السياليك على خلايا الجهاز التنفسي البشرية.[101]
البروتينات اللابنيوية
يتكون إطار القراءة المفتوح ORF1ab من إطارين مفتوحين ORF1a وORF1b متصلين عبر انزياح إطار ريبوسومي مبرمج يسمح للريبوسوم بالاستمرار في الترجمة بعد كودون التوقف الموجود في نهاية ORF1a، ويسمى هذان الإطاران في بعض الأحيان بجين الريبليكاز.[103] يشغل جين الريبليكاز المقبع 5' حوالي ثلثي الجينوم ويُترجم بترجمة معتمدة على القبعة.[104] إلى عديدي البروتين pp1a وpp1ab الذين يخضعان لتحلل بروتيني تنتج منه عدة بروتينات لابنيوية ذات وظائف مختلفة في دورة حياة الفيروس، منها بروتيازات ومكونات مركب المضاعف الناسخ.[104][105][106]
يحدث انزياح الإطار في تسلسل زلق تليه بنية رنا ثانوية على شكل عقدة كاذبة.[104] وتم قياس نسبة فعالية انزياح الإطار بين 20-50% لدى فيروس كورونا الفأري،[107] و45-70% لدى سارس-كوف-2،[108] وهو ما يعني التعبير عن بروتين pp1a حوالي 1.5 إلى 2 مرة أكثر من البروتين pp1ab.[105] يوجد في فيروسات كورونا 16 بروتينا لابنيويا، يحتوي عديد البروتين pp1a على البروتينات اللابنيوية من 1 حتى 11 ويحتوي عديد البروتين pp1ab على البروتينات اللابنيوية من 1 حتى 10 ومن 12 حتى 16. يقوم بالمعالجة التحللية (القص) بروتيازان: نطاق بروتياز مماثل للبابايين يتواجد في البروتين اللابنيوي 3 متعدد النطاقات ويقص البروتينات من 1 حتى 4، ثم يقوم البروتياز 3CL (ويُعرف بالبروتياز الرئيسي، وهو البروتين اللابنيوي 5) بباقي عمليات القص حتى النهاية الكربوكسيلية لعديد البروتين.[104][105]
يسرد الجدول بالأسفل جميع الوظائف المعروفة لحد الآن للبروتينات اللابنيوية الخاصة بفيروساات كورونا:[55][104][105][109][110]
| البروتين اللابنيوي | الوظيفة |
|---|---|
| البروتين اللابنيوي 1 | تفكيك الرنا الرسول الخلوي، تثبيط الترجمة الخاصة بالخلية المضيفة، تثبيط الإنترفيرون، غير متواجد لدى فيروسات كورونا غاما |
| البروتين اللابنيوي 2 | غير معروفة، يرتبط بالبروهيبتين |
| البروتين اللابنيوي 3 | بروتين متعدد النطاقات مع نطاق أو اثنين وظيفتهما ببروتياز مماثل للبابايين لقص البروتينات، مناهض للإنترفيرون، عدة أدوار أخرى. |
| البروتين اللابنيوي 4 | تكوين الحويصلات مزدوجة الغشاء |
| البروتين اللابنيوي 5 | بروتياز 3CL لمعالجة وقص عديد البروتين، تثبيط الإنترفيرون |
| البروتين اللابنيوي 6 | تكوين الحويصلات مزدوجة الغشاء |
| البروتين اللابنيوي 7 | عامل مرافق وعامل عملياتية لبوليمراز الرنا المعتمد على الرنا، يشكل مركبا مع nsp8 وnsp12. |
| البروتين اللابنيوي 8 | عامل مرافق وعامل عملياتية لبوليمراز الرنا المعتمد على الرنا، يشكل مركبا مع nsp7 وnsp12 |
| البروتين اللابنيوي 9 | يرتبط بالرنا مفرد السلسلة |
| البروتين اللابنيوي 10 | عامل مرافق للبروتينين nsp14 و nsp16. |
| البروتين اللابنيوي 11 | غير معروفة |
| البروتين اللابنيوي 12 | بوليمراز الرنا المعتمد على الرنا، وناقل النوكليوتيديل |
| البروتين اللابنيوي 13 | هيليكاز، وثلاثي فوسفاتاز الرنا. |
| البروتين اللابنيوي 14 | نوكلياز خارجي مصحح، تكوين قبعة الرنا، ناقل ميثيل-N7 غوانوزين. |
| البروتين اللابنيوي 15 | ريبونوكلياز داخلي، تجنب المناعة. |
| البروتين اللابنيوي 16 | ناقل ميثيل2'-O- ريبوز، تكوين قبعة الرنا. |
البروتينات الملحقة
فضلا عن الجينات المذكورة أعلاه، تحتوي معظم جينومات فيروسات كورونا على العديد من إطارات القراءة المفتوحة التي تشفر بروتينات ملحقة أو إضافية، وكلها تقع مع الاتجاه بالنسبة لجين الريبليكاز وهي موزعة بين أو داخل جينات البروتينات البنيوية الأربعة وبعضها له صلة بها. يمكن أن تكون إطارات القراءة المفتوحة الخاصة بالبرروتينات الملحقة متداخلة وتختلف أعدادها وأطوالها ومواضعها ووظائفها من فيروس إلى آخر، وهي في الغالب أصغر من البروتينات البنيوية وليست لازمة لتكاثر وتضاعف الفيروس وخاصة في الخلايا المختبرية.[111] من وظائف البروتينات الملحقة تعديل الاستجابات المناعية والتهرب منها، لها دور في إجهاد الشبكة الإندوبلازمية والإستماتة والالتهام الذاتي، تعمل كفيروبورينات وتزيد من حدة الفيروس.[112] على سبيل المثال تمنع البروتينات الملحقة استجابة الإنترفيرونات من النوع 1 وهي خط الدفاع الأول ضد الفيروسات بطرق واستراتيجيات متنوعة حيث يقوم ORF3b في سارس-كوف باستهداف MAVS[113] ويقاطع ORF9b الخاص بسارس كوف-2 إضافة اليوبيكوتين إلى نيمو لتثبيط مسار RIG-I-MAVS ومنع تخليق الإنترفيرونات.[114] يرتبط ORF6 الخاص بسارس-كوف بكاريوفيرين ألفا 2 لمنع نقل ستات 1 إلى النواة.[115] ويثبط NS4a الخاص بميرس-كوف استجابات الإجهاد الفيروسية التي يتوسطها PKR.[116]
الدخول إلى الخلية
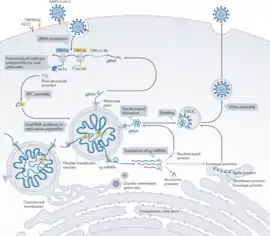
تستخدم معظم فيروسات كورونا بروتينات الشوكة للارتباط بالمستقبلات الموجودة على سطح الخلايا المضيفة، ولكن يمكن لبعض فيروسات كورونا أن ترتبط بالخلايا المضيفة ببروتينات أخرى مثل بروتين الغشاء الخاص بفيروس كورونا البشري NL63 وإستراز الراصة الدموية في الفيروسات الإمبيكوية. حيث يمكن لكلاهما أن يرتبط مع عديدات السكاريد على سطح الخلية للمساعدة في الإصابة.[78] بعد ارتباط بروتين الشوكة بالمستقبل، تقوم بروتيازات المضيف (مثل الكاتبسين وبروتياز السيرين عبر الغشائي 2) بقصه وتنشيطه. تبعا لسلالة الفيروس، يمكن للفيروس النشط أن يدخل الخلية عن طريق الإدخال الخلوي أو يندمج مباشرة مع غشاء الخلية المضيفة ويدخل السيتوبلازم.[117] يستخدم كل من: فيروس كورونا السنوري، 229E، فيروس كورونا الطيري، سارس-كوف، فيروس إلتهاب الكبد الفأري A59 وفيروس إلتهاب الكبد الفأري JHM الطريقة الثانية.[118] فضلا عن البروتيازات، تؤثر بروتينات مضيفة أخرى على عملية إصابة الفيروس للخلية مثل: البروتين الغشائي المستحث بالإنترفيرون الذي يمنع الفيروسات من دخول الخلايا، والبروتين المحتوي على الفالوسين الذي يساعد الفيروسات الموجودة في الجسيمات الداخلية على دخول السيتوبلازم.[10][119]
المستقبلات التي تستخدمها فيروسات كورونا للدخول إلى الخلية متنوعة وغير مرتبطة بجنس محدد فمثلا: ينتمي فيروس كورونا البشري NL63 إلى جنس فيروسات ألفا وسارس-كوف إلى فيروسات بيتا، لكن يستخدم كلاهما الإنزيم المحول للأنجيوتنسين 2 (ACE2) كمستقبل. أما فيروس التهاب المعدة والأمعاء الساري وفيروس الإسهال الوبائي الخنزيري وهما من نفس جنس فيروس NL63 (ألفا) يستخدمان مستقبل أمينوببتيداز الألانين. يستخدم فيروس كورونا الفأري وميرس-كوف اللذان ينتميان إلى فيروسات بيتا (مثل سارس-كوف) مستقبلي جزيء التصاق الخلية المرتبط بالمستضد السرطاني المضغي 1 والببتيداز نازع الببتيديل 4 على التوالي. علاوة على ذلك، تستخدم العديد من فيروسات كورونا جزيئات غليكوزيل معدلة موجودة في بروتينات على سطح الخلية كمستقبلات أو مستقبلات مساعدة.[85] على سبيل المثال، يستخدم فيروس كورونا البشري OC43 وفيروس كورونا البقري 9-O-حمض السياليك كمستقبل.[101]
ترجمة الجينوم

بعد دخول القفيصة إلى السيتوبلازم، تقص بروتيازات خلوية بروتينات القفيصة النووية لتحرير الجينوم الفيروسي.[58] ولأن جينوم فيروس كورونا رنوي موجب الدلالة ويملك قبعة 5' وذيل عديد الأدينين مثل الرنا الرسول الخاص بحقيقيات النوى فإنه يُترجم مباشرة بواسطة الريبوسوم.[61] أول ما تتم ترجمته هو جين الريبليكاز وفي سبيل التعبير عن كلا عديدي البروتين، يستخدم الفيروس تسلسلا زلقا 5'-UUUAAAC-3' وعقدة كاذبة لإحداث انزياح إطار ريبوسومي -1 للاستمرار في الترجمة وإنتاج عديد البروتين الكبير pp1ab. في معظم الحالات، يقوم الريبوسوم بفك بنية العقدة الكاذبة والاستمرار في الترجمة حتى يصادف كودون التوقف الخاص بـ ORF1a. لكن في بعض الأحيان وبمساعدة بروتينات فيروسية أو خلوية،[10] تعيق العقدة الكاذبة الريبوسوم عن الاستمرار في الترجمة وتتسبب في توقفه مؤقتًا على التسلسل الزلق، وهو ما يؤدي إلى تغيُر إطار القراءة عبر العودة إلى الخلف بنيوكليوتيد واحد (انزياح إطار -1)، قبل أن يتمكن الريبوسوم من فك بنية العقدة الكاذبة والاستمرار في ترجمة ORF1b وإنتاج عديد البروتين الكامل pp1ab.[120][121]
من غير المعروف بالضبط سبب استخدام الفيروسات لانزياح الإطار للتحكم في التعبير عن البروتينات، لكن افتُرض أن السبب إما من أجل التحكم الدقيق في نسبة بروتينات عديد البروتين pp1a وبروتينات عديد البروتين pp1b، أو لتأخير إنتاج بروتينات pp1b حتى تحضِّر بروتينات pp1a بيئة مناسبة لنسخ ومضاعفة الجينوم الفيروسي.[55]
بعد ترجمة إطار القراءة ORF1a تقوم البروتينات اللابنيوية 3 و4 و6 [61][122] بإحداث تغييرات في بنية الشبكة الإندوبلازمية وإنشاء عضيات التضاعف (ROs) التي يحدث فيها تضاعف الفيروس. تختلف فيروسات كورونا في نوع عضيات التضاعف التي تُحدثها وتشمل هذه العضيات: الأغشية الملففة (CM)، الحويصلات مزدوجة الغشاء (DMV)، الشبكة الإندوبلازمية المزمومة (zippered ER)، وتشير الدراسات إلى أن الحويصلات مزدوجة الغشاء هي مركز تضاعف الفيروس.[9][61] تنشأ الحويصلات مزدوجة الغشاء من حويصلات مشتقة من الشبكة الإندوبلازمية، حيث تقوم بروتينات فيروسية بتسخير مسار تفكيك البروتين المرتبط بالشبكة الإندوبلازمية (ERAD) لإنشائها.[82][123] كما يمكن أن تنشأ من استثارة مسار الالتهام الذاتي.[124] يُعتقد أن القيام بعملية النسخ والتضاعف داخل الحويصلات مزدوجة الغشاء آلية لإخفاء الرنا الفيروسي ووسيلة لتجنب الاستجابات المضادة للفيروسات التي تُحدثها جزيئات الرنا مزدوجة السلسلة.[125]
الناسخ المضاعف

في الحويصلات مزدوجة الغشاء، تتجمع العديد من البروتينات اللابنيوية (البروتين اللابنيوي 7 و8 و9 و10 و12 و13 و14 و16[61]) التي نتجت من قص عديد البروتين pp1ab لتشكيل مركب التضاعف والنسخ لنسخ ومضاعفة جينوم الرنا الفيروسي. أهم بروتينات هذا المركب هو البروتين اللابنيوي 12 (بوليميراز الرنا، الذي يحفز تخليق الرنا، أما البروتينات الأخرى فلها وظائف مساعدة مختلفة مثل: البروتين اللابنيوي 14 (نوكلياز خارجي للرنا) الذي يوفر وظيفة تصحيح ويقوم بإزالة النيوكليوتيدات غير المتطابقة وتحسين دقة النسخ.[126] تملك فيروسات الرنا عموما معدل طفرات عالٍ (10−5 إلى 10−3 طفرة لكل موقع لكل عملية نسخ) حين يتجاوز طول الجينوم 15 ألف نوكليوتيد وينتج عن ذلك حدوث كارثة خطأ بسبب تراكم العديد من الطفرات، لذلك جينومها له حد علوي.[61] لكن معدل الطفرات في فيروس كورونا أقل بكثير من باقي فيروسات الرنا الأخرى بسبب وظيفة التصحيح الخاصة بالبروتين اللابنيوي 14 ويُقدر بـ (10−7 إلى 10 −6 طفرة لكل موقع لكل نسخ) وهو قريب من معدل الطفرات لفيروسات الدنا مفردة السلسلة،[127][128] لذلك يمكن لجينوماتها بلوغ طول كبير يتجاوز 30 ألف نوكليوتيد. فضلا عن فيروسات كورونا، فيروسات الرنا الأخرى التي تملك وظائف التصحيح هي: فيروسات تورو والفيروسات الرونية وينتمي كلاهما إلى رتبة الفيروسات العشية وكلاهما يحتوي على جينومات أطول من جينومات فيروسات الرنا الأخرى.[61][129] بالإضافة إلى هذه البروتينات اللابنيوية الفيروسية، يوجد العديد من بروتينات الخلية المضيفة في الغشاء الداخلي للحويصلات مزدوجة الغشاء بما في ذلك البروتينات المسؤولة عن النقل الحويصلي والبروتينات المرتبطة باليوبيكويتين والبروتينات المرتبطة بمسار الالتهام الذاتي والعديد من بروتينات بدء الترجمة التي تساعد في ترجمة الرنا. تبّين أن مركب التضاعف والنسخ يتم تجميعه بجوار الريبوسومات على غشاء الشبكة الإندوبلازمية لتحسين كفاءة ترجمة الرنا وتخليق البروتينات.[61][130]
يتكون لب مركب التضاعف والنسخ في سارس-كوف-2 من بوليميراز الرنا المعتمد على الرنا (البروتين اللابنيوي 12، nsp12) وثلاث بروتينات لابنيوية: البروتين اللابنيوي 7 ونسختان من البروتين اللابنيوي 8.[131] طول بوليميراز الرنا في سارس-كوف-2 932 حمض أميني ويتكون من أربع نطاقات رئيسية: نايران (NiRAN) ويملك وظيفة ناقل نوكليوتيديل، الواجهة، الأصابع والكف.[132] ترتبط بالبوليميراز نسختان من البروتين اللابنيوي 8، وبسبب عدم تناظر البوليميراز فإن البروتين اللابنيوي 7 مطلوب من أجل أن يتوسط التآثرات بين البروتينين اللابنيويين 12 و8 على إحدى الجهتين.[133][134] يرتبط هليكازان (البروتين اللابنيوي 13) بامتدادات لولبية للبروتينين 8 فوق لولب الرنا المزدوج (الصورة)، ويملك هذان الهليكازان تآثرات محدودة بين بعضهما البعض ويبدو أنهما يعملان على استقرار البنية الإجمالية لمركب التضاعف والنسخ.[135] يعمل الهليكازان على تسهيل نسخ جينوم الرنا المليء بالبنى الثانوية والثالثية عبر تفكيك هذه البنى أما البروتينات اللابنيوية 9 و10 و14 و16 فلها أدوار في تنظيم تخليق القبعة 5' واستقرار بنى جينوم الرنا.[58]
فضلا عن الهيئة الموحودة لبوليميراز الرنا، يمكن أن يشكل هذا الأخير مثنويا، يرتبط فيه كل بوليميراز بنسخة واحدة من البروتين 8 ويتثنيان عبر تآثرات بين البروتينين 7 الخاصين بهما ليتخذا هيئة توازي متضاد. افترض أن هذه الهيئة المثنوية تلعب دورا في نسخ الرنا الجينومي الفرعي وتبديل القالب.[136]
الرنا الجينومي الفرعي وتبديل القالب
تملك الفيروسات العشية سمة مميزة لها عن باقي فيروسات الرنا وهي إنتاجها لرنا جينومي فرعي بأطوال مختلفة لتخليق البروتينات البنيوية والملحقة عبر آلية تبديل قالب معقدة.[137] بعد تشكل مركب التضاعف والنسخ في الحويصلات مزدوجة الغشاء، يستخدم مركب التضاعف الرنا الفيروسي الموجب لإنتاج رنا سالب كامل الطول عبر نسخ مستمر، ثم يُستخدم هذا الرنا السالب كقالب لإنتاج رنا جينومي موجب كامل الطول. فضلا عن التضاعف الجينومي بالنسخ المستمر، يقوم مركب التضاعف بنسخ متقطع ينتج عنه رنا فرعي بأطوال مختلفة.[138]
يتم إنشاء الرنا الفرعي عند إنشاء السلسلة السالبة عبر نموذج نسخ متقطع يفترض: وجود مواقع خاصة تسمى التسلسل المنظم للنسخ الجسمي (TRS-B) في بداية كل جين والتي تشكل تآثرات رنا-رنا مكملة بعيدة المدى مع التسلسل المنظم للنسخ الطليعي (TRS-L) المتواجد في حلقة الـSL3 في المنطقة غير المترجمة 5'.[139][140] تُنظَّم هذه التآثرات بين التسلسل الجسمي والطليعي بواسطة مركبات رنا-بروتين يُفترض أنها تساهم في توقف مركب التضاعف والنسخ عند التسلسل الجسمي.[141] بعد نسخ التسلسل الجسمي ينفصل مركب التضاعف والنسخ عنه ويعيد الارتباط بالتسلسل الطليعي مبدلا القالب ومتجاوزا جزءا كبيرا من الجينوم ويقوم بنسخ التسلسل الطليعي.[138] بعد نسخ جزيئات الرنا الفرعية السالبة، تُستخدم هذه الأخيرة لنسخ رنا فرعي موجب، وعادة ما يُترجم من الرنا الجينومي الفرعي الموجب إطار القراءة المفتوح الأول عكس الاتجاه فقط، أما الرنا الجينومي السالب والرنا الجينومي الفرعي السالب فهما أقل تواترا في الخلايا المصابة وقد لا يشفران أي بروتينات فيروسية.[58]
يُنظَّم تبديل القالب بواسطة بروتينات فيروسية وخلوية أهمها بروتين القفيصة النووية الذي يملك نشاط شابرون رنا يسهل عملية تبديل القالب وهو مطلوب ليكون النسخ فعالا،[142] ومن بين وظائفه أنه يقوم بتوظيف الهليكاز الخلوي DDX1 الذي يسمح بتجاوز التوقف عند التسلسل الجسمي لنسخ رنا فرعي أطول أو رنا جينومي كامل.[143] رغم أن معظم الرنا الفرعي (~90%) ينشأ عبر تبديل القالب بين التسلسلات الجسمية والتسلسل الطليعي إلا أن نسبة صغيرة منه (< 10%) يمكن أن تنشأ بطريقة مستقلة عن التسلسل الجسمي أو حتى مستقلة تماما عن التسلسلات المنظمة للنسخ،[139][140][144] وهو ما يشير إلى حدوث تآثرات رنا-رنا غير قياسية تسببها بعض بُنى الرنا أو ارتباط عوامل فيروسية أو خلوية أثناء عمليات تبديل القالب، مثل تآثرات الرنوات النووية الصغيرة الخلوية (U1 وU2 وU4) مع مواقع محددة في إطاري القراءة ORF1a وORF1b.[140]
التأشيب الجيني
لأن مركب التضاعف والنسخ بإمكانه تبديل القالب أثناء النسخ عند التسلسل المنظم للنسخ، فإنه يستطيع تبديل نسخ سلسلة فيروس بأخرى حين يتواجد أكثر من جينوم رنا فيروسي واحد في الخلية، وينتج عن ذلك تأشيب لجينات الرنا الفيروسي.[61][145] تفاصيل الآلية مازلت غير معروفة، لكن أظهرت دراسة أن البروتين اللابنيوي 14 المسؤول عن التصحيح له دور كذلك في آلية التأشيب الجيني.[61] التأشيب الجيني آلية مهمة في تنوع فيروسات كورونا، وتسمح بتطفير تسلسلات الفيروس لتوليد سلالات وأنواع جديدة وكذلك السماح للفيروسات بالانتقال عبر الأنواع.[146] تملك عدة فيروسات كورونا سجلات على أحداث تأشيب جيني بينها، على سبيل المثال فيروس كورونا البشري OC43 المقسم إلى خمسة أنماط من A حتى E: النمط D هو ناتج تأشيب بين النمطين B وC، والنمط E هو ناتج تأشيب بين الأنماط B وC وD.[147] تملك مستخلصة أمستردام-1 من فيروس كورونا البشري NL63 جزءا من تسلسل بروتين الشوكة الخاص بمستخلصة 496 وذلك بسبب تأشيب جيني،[148] سارس-كوف (وهو من نمط بيتا) قد تكون له علاقة بالأنماط ألفا وغاما فقد وجدت دراسات علم تطور السلالات فيه مناطق تأشيب محتملة بين فيروسات من نمط ألفا وغاما وفيروسات أخرى من نمط بيتا،[149] والعديد من فيروسات كورونا الخفاشية المرتبطة بالسارس عادة ما تقوم بتأشيبات جينية بين بعضها البعض. المصدر الرئيسي لنشوء فيروس السارس قد يكون حدوث تأشيبات بين عدة فيروسات كورونا خفاشية.[150]
التجميع والتحرير

بعد نسخ الرنا الجينومي والفرعي تنتقل هذه الجزيئات إلى السيتوبلازم عبر مسامات موجودة في الحويصلات مزدوجة الغشاء، ويُترجم الرنا الفرعي لإنتاج البروتينات البنيوية والملحقة وتتموقع البروتينات البنيوية في الشبكة الإندوبلازمية وجهاز غولجي أين تتعرض لتعديلات ما بعد الترجمة،[55] ثم تنتقل هذه البروتينات ومنها البروتين الشوكي في هيئة قبل الاندماج المثلوثية عبر حويصلات صغيرة إلى الحويصلات مفردة الغشاء (SMV) -وهي مكان تجميع الفيروس- وتندمج بها.[151] ثم يتجمع بروتين الشوكة والغشاء والغلاف على جزءٍ من غشاء الحويصلة مفردة الغشاء وتتسبب تآثرات بينه وبين بروتينات الغلاف والغشاء في انحنائه.[152][153] وبوجود القفيصة الفيروسية وتآثرات بين بروتينات الغشاء والقفيصة يحدث تبرعم الفيروس إلى داخل الحويصلة مفردة الغشاء التي تحتوي في النهاية على العديد من الفيروسات ثم تغادر هذه الأخيرة الخلية عبر الإخراج الخلوي.[151] بعدها يمكن للفيروسات الجديدة المحررة إصابة خلايا أخرى.[154] عملية تجميع فيروس كورونا يمكن أن تساهم فيها بروتينات من الخلية المضيفة مثل الأنيبيبات الدقيقة، خيوط الأكتين والفيمنتينات، وتختلف هذه البروتينات حسب الفيروس.[10]
يقوم بروتين القفيصة النووية بتغليف الرنا الجينومي لتكوين القفيصة النووية وتكديسها من أجل تعبئتها في الفيروس من خلال التآثر مع إشارة التغليف التي يختلف موقع تواجدها حسب نوع الفيروس ولا توجد سوى في الرنا الجينومي الفيروسي الكامل وهي الطريقة التي يتجنب بها الفيروس تغليف رنا جينومي فرعي أو رنا يخص الخلية المضيفة، ويمكن لكلا بروتيني الغشاء والقفيصة التعرف على هذه الإشارة والتآثر معها.[69] آلية تكوين القفيصة وتجميع الفيروس غير واضحة تماما، لكن تشير دراسات على سارس-كوف-2 إلى أن بروتين القفيصة عند ارتباطه بالرنا الجينومي يخضعان لانفصال طور سائل-سائل ويشكلان تكثفات أو تكدسات.[95][155] يتكون بروتين القفيصة من نطاقين منحفظين: نطاق أميني يرتبط بالرنا يُعتقد أنه يقوم بتآثرات مع إشارة التغليف ونطاق كربوكسيلي يشكل مثنوي متجانس يُفترض أن له دور في تجميع البروتين النووي الريبوزي (تكوين القفيصة)،[156] وتتواجد وسط وعلى جانبي هذين النطاقين نطاقات مضطربة جوهريا (IDRs) (الصورة)، النطاق المركزي عالي الانحفاظ وغني بالسيرين وفسفرته تلعب دورا في سلوك الهلام أو السائل لتكثفات بروتين القفيصة والرنا.[157] النطاقان الجانبيان أقل انحفاظا والنطاق الكربوكسيلي منهما الخاص بسارس-كوف يرتبط بالبروتين الغشائي وهو ما يوحي بدور مهم لهذا النطاق في تعبئة وتجميع الفيروس.[158]
أشارت دراسات سابقة إلى أن البروتينات النووية الريبوزية الفيروسية (vRNP) خيطية لولبية الشكل لكن أظهرت دراسات بالتصوير المقطعي البردي الإلكتروني (cryo-ET) على سارس-كوف-2 بُنى حبيبية الشكل، حيث يحتوي كل فيروس على 35-40 مركب بروتيني نووي ريبوزي يتخذ كل واحد منها بنية أسطوانية صدفية قطرها 15 نانومتر.[95][159] وداخل الفيروس، تتجه هذه المركبات القريبة من الغشاء الفيروسي باتجاه الغشاء، الأمر الذي يوحي بأنها تتآثر بشكل خاص مع البروتين الغشائي. وداخل مركب البروتين النووي الريبوزي، تقترح نمذجة تجريبية مستندة على بُنى النطاقين الأميني والكربوكسيلي أنه يتكون من حوالي 800 نوكليوتيد مغلفة بحوالي 12 نسخة من بروتين القفيصة،[95][159] يمكن لمركبات البروتين النووي الريبوزي المفردة أن تشكل تكدسات خطية وهذا ما يفسر التضارب في الشكل الملاحظ لها ويوحي بأنها يمكن أن تتخذ أشكالا منحفظة متنوعة.[95]
الانتقال
الأفراد المصابون قادرون على نشر الفيروسات في البيئة. تآثر بروتين الشوكة مع مستقبل خلوي مكمل له أمر أساسي في تحديد انتحاء الأنسجة، شدة العدوى، ومجال الأنواع التي يصيبها الفيروس.[49][160] تستهدف فيروسات كورونا بشكل أساسي الخلايا الظهارية،[47] وتنتقل من مضيف إلى مضيف آخر حسب نوع الفيروس إما عن طريق الرذاذ التنفسي أو المواد المحتوية على الفيروس، أو الطريق الفموي الشرجي.[161]
تصيب فيروسات كورونا البشرية الخلايا الظهارية في الجهاز التنفسي، بينما تصيب فيروسات كورونا الحيوانية عموما الخلايا الظهارية في الجهاز الهضمي.[47][162] على سبيل المثال، يصيب فيروس كورونا السارس الخلايا الظهارية للرئة البشرية عن طريق الهباء الجوي بالارتباط بمستقبل الإنزيم المحول للأنجيوتنسين. ويصيب فيروس التهاب المعدة والأمعاء الساري الخلايا الظهارية للخنازير في الجهاز الهضمي عبر الطريق الفموي الشرجي بالارتباط بمستقبل أمينوببتيداز الألانين.[161]
الالتهام الذاتي
بعد الإصابة بفيروس كورونا يمكن للبروتينات اللابنيوية أن تستثير الالتهام الذاتي، إذ ترتبط عملية تكوين الحويصلات مزدوجة الغشاء (DMV، موقع تضاعف الرنا الفيروسي[163]) في الشبكة الإندوبلازمية ارتباطا وثيقا باستجابة الالتهام الذاتي، وتتشكل بعد إعادة تنظيم أو تغيير بنية غشاء الشبكة الإندوبلازمية. مع ذلك، توجد نتائج تجريبية تفيد بأن فيروسات كورونا يمكنها أن تصيب بنجاح الخلايا التي تعوز بروتينات الالتهام الذاتي المهمة مثل ATG5. لذلك يمكن لبعض فيروسات كورونا (مثل: فيروس التهاب الكبد الفأري، وفيروس التهاب الشعب الهوائية المعدي، وفيروس كورونا البشري NL63) أن تحفز تكوين الحويصلات مزدوجة الغشاء من خلال مسار آخر (مسار ERAD) وهو مسار مستقل عن مسار الالتهام الذاتي، لكنه يستعين ببعض البروتينات المستخدمة في مسار الالتهام الذاتي مثل MAP1LC3B.[10][124] في استجابة الالتهام الذاتي، يقوم BECN1 بتحفيز اندماج الجسيمات البلعمية الذاتية واليحلولات لتفكيك المواد الموجودة فيها، ويمكن لفيروسات كورونا أن تمنع عمل BECN1 بعدة طرق لتجنب اندماج الحويصلات مزدوجة الغشاء مع اليحلولات.[124]
استجابة إجهاد الشبكية الإندوبلازمية
تؤدي عملية تضاعف فيرووس كورونا وتجميعه إلى تغيّر ملحوظ في بنية الشبكة الإندوبلازمية، حيث تترجَم جميع بروتينات الفيروس بواسطة الريبوسومات في الشبكة الإندوبلازمية ويخضع معظمها إلى تعديلات بعد ترجمة متنوعة منها: الغلكزة، إضافة البالميتيك إضافة اليوبيكويتين، إضافة الـADP ريبوز، إضافة بروتين سومو، كما أن تطوي وبلمرة هذه البروتينات يعتمد على مساعدة الشابرونات الموجودة داخل الشبكة الإندوبلازمية وهو ما يزيد العبء على هذه الأخيرة. فضلا عن ذلك، تؤدي الإصابة بالفيروس إلى استثارة مسار الالتهام الذاتي ومسارات أخرى تُنتِج الكثير من الحويصلات التي تستهلك مكونات غشاء الشبكة الإندوبلازمية وهو ما يُحدث ضغطا على الشبكة. تؤدي جميع هذه العوامل إلى إثارة استجابة خلوية تسمى استجابة إجهاد الشبكية الإندوبلازمية.[82] تُحفز استجابة إجهاد الشبكة عدة مسارات خلوية تقوم بتقليل ترجمة البروتينات وتعمل كآلية لمحاربة الإصابة بالفيروس، ففي بعض فيروسات كورونا يُنشِّط إجهاد الشبكة كينازات عامل البدء 2 مثل: EIF2AK3 وPKR في المرحلة الأولى من الإصابة والتي تقوم بفسفرة عامل البدء eIF2 لمنع الترجمة وتحفير الاستماتة وتخليق السيتوكينات.[82][164] على سبيل المثال: يمكن تحفيز التعبير عن الإنترفيرون من النوع 1 بعد أن تنشِّط الإصابة بفيروس التهاب المعدة والأمعاء الخنزيري مسار EIF2AK3.[10][165] يمكن لإجهاد الشبكة كذلك أن ينشط مسار IRE1-XBP1[166] ومسار ATF6،[167] والعديد من المسارات المختلفة التي تؤدي إلى تخليق البروتينات المضادة للإجهاد والسيتوكينات.[82]
الاستماتة والسيتوكينات
يمكن لفيروسات كورونا أن تُحدِث الاستماتة من خلال عدة آليات، إذ يمكن أن يسبب سارس-كوف استماتة خلايا الرئة والطحال والغدة الزعترية وأنسجة أخرى من خلال مسار الكاسباز،[168] ويمكن أن يسبب ميرس-كوف الاستماتة لأنسجة الجهاز التنفسي والخلايا التائية بطرق مختلفة، كما يمكن أن يتسبب فيروس كورونا البشري OC43 في استماتة الخلايا العصبية بمسار مستقل على الكاسباز لكن يستخدم بروتين Bax .[169] ويُحدث سارس-كوف-2 الاستماتة كذلك.[170][171] يمكن أن تتسبب الإصابة بفيروس كورونا أيضا بشكل غير مباشر في استماتة الخلايا المصابة عن طريق تحفيز استجابة إجهاد الشبكة الإندوبلازمية أو تحفيز مسار MAPK/ERK.[10] فضلا عن أنسجة الانتحاء الرئيسية، تسبب الفيروسات المذكورة سابقا موت العديد من أنواع الخلايا المناعية. إذ يمكن لفيروس كورونا البشري 229E أن يقتل الخلايا المتغصنة (ولكن ليس من خلال الاستماتة)[172]، وبالتالي يمكن أن يثبط الاستجابة المناعية للمضيف.[10] هناك العديد من الطرق للتسبب في الاستماتة بما في ذلك مسارات استجابة إجهاد الشبكة الإندوبلازمية المذكورة آنفا، والعديد من مسارات MAPK التي يمكن أن تسببها عدوى فيروس كورونا. على سبيل المثال، بعد تنشيط كينازات p38، يمكن فسفرة EIF4E لتثبيط الترجمة وتحفيز الاستماتة، تحفز كينازات JNK الاستماتة كذلك وقد يكون لها وظائف أخرى مضادة للفيروسات. مع ذلك، يوجد مسار MAPK آخر يُحدثه فيروس كورونا هو مسار ERK له نتائج عكس نتائج مسار كينازات p38، إذ يقوم بفسفرة eIF4EBP1 وتحرير eIF4E المرتبط به ليقوم بدوره في بدء عملية الترجمة.[10]
يمكن للإصابة بفيروس كورونا أن تتسبب في تخليق مختلف السيتوكينات عبر مسارات مختلفة مما يؤدي إلى استجابة التهابية، وهي الاستجابة المناعية الفطرية للخلايا المضيفة. فضلا عن المسارات المذكورة أعلاه التي تحفز التعبير عن السيتوكينات نتيجة إجهاد الشبكة الإندوبلازمية، يمكن لفيروسات كورونا أيضا أن تنشط العديد من مسارات MAPK في الخلايا مثل: مثل p38 وERK وJNK [10] ويمكن لهذه المسارات الثلاثة تحفيز التعبير عن مجموعة متنوعة من السيتوكينات. تم الإبلاغ عن سيتوكينات مثل: CCL2 وإنترلوكين 8 وإنترلوكين 6 وTNF-α وCOX-2 في مختلف فيروسات كورونا.[10][173][174]
المناعة التكيفية
بعد إصابة المضيف، يمكنه تطوير مناعة تكيفية ضد فيروس كورونا عبر تنشيط خلايا تائية وخلايا بائية محددة. يمكن أن تستمر مناعة الأشخاص الأصحاء ضد فيروسات كورونا الأربعة (229E وOC43 وNL63 وHKU1) التي تسبب الزكام لنحو نصف عام إلى عام فقط، وبعد ذلك يمكن أن يصابوا بنفس الفيروس مرة أخرى.[175] يتم إنتاج الأجسام المضادة ضد بروتين الشوكة الخاص بسارس-كوف في غضون ثلاثة أسابيع،[176] وقد يكون الوقت اللازم لإنتاجها في سارس-كوف-2 أقصر.[61][177] عيار الأجسام المضادة لمرضى السارس سوف يتغير بمرور الوقت، تتبعت بعض الدراسات المرضى في حدث السارس ووجدت أن استجابات الأجسام المضادة وخلايا البائية الذاكرة في المرضى انخفضت بشكل ملحوظ بعد عدة سنوات. في المقابل، يمكن أن يحافظ الجسم على خلايا التائية الذاكرة لفترة أطول.[61][178] مع ذلك، قد تختلف مدة المناعة من شخص لآخر حيث وُجد أنه لا يزال بالإمكان اكتشاف الأجسام المضادة لدى بعض المرضى بعد إصابتهم بالسارس منذ أكثر من عشر سنوات. تختلف مدة بقاء الأجسام المضادة التي تنتجها الإصابة بفيروس ميرس-كوف تبعا لشدة الأعراض، فعيار الأجسام المضادة بعد سنتين من الإصابة أكبر بكثير لدى مرضى الحالات الحادة منه لدى مرضى الحالات المتوسطة.[179]
يدوم مفعول التطعيم ضد فيروس التهاب الشعب الهوائية المعدي في الدجاج حوالي 9 أسابيع فقط وكان فعالا ضد بعض السلالات فقط .[180] يشبه فيروس كورونا التنفسي الخنزيري (PRCV) إلى حد كبير فيروس التهاب المعدة والأمعاء الساري الخنزيري، ويمكن للخنازير المصابة به اكتساب مناعة ضده فقط وليس لكليهما، لكن المناعة المخاطية ضده (الغلوبولين المناعي أ) تضعف بمرور الوقت، الأبقار المصابة بفيروس كورونا البقري تضعف مناعتها بمرور الوقت كذلك. تؤدي الاستجابة المناعية الناتجة عن الإصابة بسلالة JHM من فيروس التهاب الكبد الفأري إلى إزالة الميالين من الخلايا العصبية في الدماغ مع القضاء على الفيروس.[179]
آليات مقاومة الفيروس للمناعة
تمتلك فيروسات كورونا مجموعة متنوعة من الآليات لمجابهة الاستجابة المناعية للخلايا المضيفة. فضلا عن وظائفها البنيوية أو التحفيزية، تملك العديد من بروتينات فيروس كورونا وظائف تهرب ومقاومة للاستجابة المناعية. ومقارنة بفيروسات الرنا الأخرى لا يسبب فيروس كورونا قدرا كبيرا من التعبير عن الإنترفيرونات من النوع الأول، مما يوحي بأن لديه آليات لمنع التعبير عنها.[181] يملك البروتين اللابنيوي 3 (بروتياز PLpro) الخاص بفيروسات: سارس-كوف، ميرس-كوف، IBV، كورونا الفأري، NL63 البشري -زيادة على وظيفة القص الذاتي- وظيفة إزالة اليوبيكويتين من بروتينات المضيف، ويملك نفس البروتين في سارس-كوف وميرس-كوف وظيفة إضافية هي إزالة وسم ISG15 من بروتينات المضيف. ولكون إضافة كلٍّ من اليوبيكويتين وISG15 تلعب دورا مهما في مسارات التأشير الخاصة بالاستجابة المناعية الخلوية، فإن إزالة هذه الوسوم تثبط الاستجابة المناعية وتخفض من التعبير عن الإنترفيرونات وهي الطريقة التي يقاوم بها البروتين اللابنيوي 3 الاستجابة المناعية،[10][182] فضلا عن آليات المقاومة بالقص هذه يملك نفس البروتين في سارس-كوف آلية لتثبيط تخليق الإنترفيرون 1 مستقلة عن القص بالبروتياز.[10][183] يمكن لكل من البروتين اللابنيوي 3 والبروتين اللابنيوي1 وORF3b وبروتين القفيصة النووية في سارس-كوف أن يثبطوا تنشيط مسار RIG-I لمنع الرنا الفيروسي مزدوج السلسلة من إثارة استجابة مناعية، ويمكن لـORF6 أن يثبط تنشيط مسار JAK/STAT بحيث تثبط كلا الآليتين تخليق الإنترفيرون 1. يمنع البروتين اللابنيوي 1 الكثير من جزيئات الرنا الرسول الخاصة بالمضيف -ومن بينها الرنا الرسول الخاص بالإنترفيرونات- من الترجمة ويسبب تفككها، أما الرنا الرسول الخاص بالفيروس في حد ذاته فهو لا يتأثر بوظيفة البروتين اللابنيوي 1 بفضل تسلسل خاص في المنطقة غير المترجمة 5'.[61][184]
فضلا عن تنشيط RIG-I، يمكن للرنا الفيروسي مزدوج السلسلة أن ينشط 2'-5' مخلقة قليل الأدينيلات لتخليق 2'-5' قليل الأدينيلات وبعدها تنشيط الريبونوكلياز L، وفي سبيل مقاومة هذه الاستجابة يقوم البروتين اللابنيوي 4B (ns4b) في ميرس-كوف والبروتين اللابنيوي 2 في فيروس التهاب الكبد الفأري بتفكيك 2'-5' قليل الأدينيلات لمنعه من تنشيط الريبونوكلياز L وبالتالي منع الرنا الفيروسي من التفكك.[61][185] يمكن لبروتين الغلاف في سارس-كوف أن يثبط مسار ERN1 الذي يُحفز بواسطة استجابة إجهاد الشبكة الإندوبلازمية وبالتالي يمنع الاستماتة وتخليق السيتوكينات.[79] وربما هذه الآلية قد تكون مرتبطة بوظيفة بروتينات الغلاف في تشكيل فيروبورينات (قنوات آيونية).[82][186] يقوم بروتين القفيصة النووية بتكوين تكثفات مع الرنا ويستخدم بروتين لب حبيبة إجهاد G3BP1 وبالتالي يحرم الخلية من استخدامه في تكوين حبيبات الإجهاد ويمنع الاستجابات المناعية الفطرية المعتمدة على G3BP1.[95]
التصنيف
فيروسات كورونا هي عائلة فرعية تنتمي إلى عائلة الفيروسات التاجية، التي تنتمي إلى الرتبة الفرعية «كورنيدوفرينيات» (Cornidovirineae) التي تنتمي إلى رتبة الفيروسات العشية.[11] تنقسم عائلة الفيروسات التاجية إلى ثلاث فروع: الكورونافيرينات المستقيمة (Orthocoronavirinae) والتي تعرف بالاسم الشهير فيروسات كورونا، [7] الليتوفيرينات، والعائلة الفرعية الجديدة البيتوفيرينات (Pitovirinae).[11] تنقسم عائلة فيروسات كورونا الفرعية إلى أربع أجناس (فيروس كورونا ألفا، فيروس كورونا بيتا، فيروس كورونا غاما، فيروس كورونا دلتا) وتصيب أجناس ألفا وبيتا الثدييات فقط، أما غاما ودلتا فيصيب معظمها الطيور بشكل رئيسي مع أنواع قليلة تصيب الثدييات.[187][188] ينتمي فيروسا سارس-كوف وسارس-كوف-2 إلى نوع الفيروسات المرتبطة بالسارس التي تنتمي إلى الجنس الفرعي الفيروسات الساربيكوية التي تنتمي إلى جنس فيروسات بيتا.[11][189] يعتمد تصنيف فيروسات كورونا على سبع تسلسلات محفوظة في جين ORF1ab فإذا كانت تسلسلات الأحماض الأمينية الناتجة من هذه التسلسلات السبع متماثلة بين سلالتين من الفيروس بنسبة تزيد عن 90% تعتبر السلالتين من نفس النوع.[190]
فيروس كورونا ألفا

النوع النموذجي لفيروس كورونا ألفا هو فيروس كورونا ألفا 1 (ألفا-كوف-1)،[191][192] الذي ينقسم إلى 14 جنسا فرعيا منها: فيروس كورونا ألفا 1 (فيروس كورونا الكلبي، فيروس كورونا القططي، فيروس التهاب المعدة والأمعاء الساري الخنزيري)، فيروس كورونا ألفا 2 (فيروس كورونا ابن مقرض، فيروس كورونا المنك)، فيروس كورونا الألبكة، فيروس كورونا البشري 229E، فيروس كورونا البشري NL63، فيروس كورونا خفاش الحذوة الكبير HuB-2013 (BtRf-AlphaCoV/HuB2013)، فيروس كورونا الخفاش محني الجناح نوع 1 (Bat-CoV MOP1)، فيروس كورونا الخفاش محني الجناح HKU8 (Bat-CoV HKU8)، فيروس كورونا خفاش مايوتيس ريكيتي Sax-2011 (BtMr-SAX2011)، فيروس كورنا الخفاشي CDPHE15 (BtCoV CDPHE15)، فيروس كورنا الخفاشي 3398 (PK-BatCoV 3398)، فيروس كورونا خفاش نيكتالوس فيلتينوس SC-2013 (BtNv-SC2013)، فيروس كورونا زبابات T14 (Sa-CoV T14)، فيروس كورونا خفاش إلف الظلام 512 (Bat-CoV 512)، فيروس الإسهال الوبائي الخنزيري (PEDV)، فيروسات خفاش الأقحوان مثل فيروس كورونا خفاش حدوة الفرس الصيني HKU2 (Bat-CoV HKU2)، فيروس كورونا متلازمة الإسهال الحاد الخنزيري (SADS-CoV)، فيروس كورونا زبابة ونتشنغ (WESV) وفيروس كورونا الجرذ البني لوتشنغ (LRNV).[193]
فيما يلي الشجرة التطورية لكل مجموعة رئيسية من فيروس كورونا ألفا (راجع أمبر بابينو وزملائها[190]، تشيتشيانغ وو وزملائه[194]):
| فيروس كورونا ألفا |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
فيروس كورونا بيتا

النوع النموذجي لجنس فيروس كورونا بيتا هو فيروس كورونا الفأري (م-كوف) الذي ينقسم إلى 5 أجناس فرعية منها: فيروس كورونا بيتا 1 (فيروس كورونا البشري OC43، فيروس كورونا البقري، فيروس كورونا الخيلي، فيروس التهاب الدماغ والنخاع الشوكي الراصي الخنزيري)، فيروس كورونا الأرنبي HKU14 (RbCoV HKU14)، فيروس كورونا الجرذ الصيني HKU24 (ChRCoV HKU24)، فيروس كورونا البشري HKU1، فيروس كورونا الفأري، فيروس كورونا ميودس 2JL14 (MrufCoV 2JL14)، فيروس كورونا خفاش أيدولون C704 (Ei-BatCoV C704)، فيروس كورونا خفاش بيبيسترالوس HKU5 (Bat-CoV HKU5)، فيروس كورونا خفاش راوستوس HKU9 (HKU9-1)، فيروس كورونا خفاش راوستوس GCCDC1 (Ro-BatCoV GCCDC1)، فيروس كورونا خفاش تيلونيكترايس HKU4 (Bat-CoV HKU4)، فيروس كورونا المرتبط بمتلازمة الشرق الأوسط التنفسية (ميرس-كوف)، فيروس كورونا القنفذ البري (EriCoV)، فيروس كورونا خفاش زاريا (ZBCoV)، فيروس كورونا خفاش برات مستدير الورق جيجيانغ2013 (BtHp- BetaCoV/ZJ2013)، وفيروس كورونا المرتبط بالمتلازمة التنفسية الحادة الشديدة (سارس-كوف، سارس-كوف-2 والسلالات المرتبطة) والعديد من الفيروسات الأخرى.[193]
فيما يلي الشجرة التطورية للمجموعات الرئيسية من فيروس كورونا بيتا (راجع أمبر بابينو وزملائها [190]، تشيتشيانغ وو وزملائه[194]):
| فيروس كورونا بيتا |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
فيروس كورونا غاما
النوع النموذجي لجنس فيروس كورونا غاما هو فيروس كورونا الطيري (AvCoV) وينقسم إلى ثلاثة أجناس فرعية منها: فيروس كورونا الحوتي، فيروس كورونا الطيري، فيروس كورونا الطيري 9203 (AvCoV 9203)، فيروس كورونا الإوزي CB17 (BcanCoV CB17)، فيروس كورونا البطي 2714 (DuCoV 2714)، وفيروسات أخرى.[193]
فيما يلي الشجرة التطورية للمجموعات الرئيسية من فيروس كورونا غاما (راجع أمبر بابينو وزملائها [190]، ميشال ويلي وزملائها[195]):
|
فيروس كورونا دلتا
النوع النموذجي لجنس فيروس كورونا دلتا هو فيروس كورونا البلبلي HKU11 (Bulbul-CoV HKU11) وينقسم إلى ثلاث أجناس فرعية منها: فيروس كورونا ودجون HKU20 (WiCoV HKU20)، فيروس كورونا البلبلي HKU11، فيروس كورونا الخنزيري HKU15 (PorCoV HKU15)، فيروس كورونا دجاج الماء الشائع HKU21 (CMCoV HKU21)، فيروس كورونا مونيا HKU13 (MuCoV HKU13)، فيروس كورونا أبيض العين HKU16 (WeCoV HKU16) فيروس كورونا العقعق روبن HKU18 (MRCoV HKU18)، فيروس كورونا بلشون الليل HKU19 (NHCoV HKU19)، فيروس كورونا الصقري HKU27 (FalCoV UAE-HKU27)، فيروس كورونا الحبارى الإفريقي HKU28 (HouCoV UAE-HKU28)، فيروس كورونا الحمام HKU29 (PiCoV UAE-HKU29)، فيروس كورونا السمان HKU30 (QuaCoV-HKU30) وفيروسات أخرى.[193]
فيما يلي الشجرة التطورية للمجموعات الرئيسية من فيروس كورونا دلتا (راجع أمبر بابينو وزملائها،[190] وميشال ويلي وزملائها[195]):
|
التطور

تستخدم بعض الدراسات بوليميراز الرنا المعتمد على الرنا كساعة جزيئية وتقدر أن أحدث سلف مشترك لجميع فيروسات كورونا برز إلى الوجود بين 8000-10000 سنة قبل الميلاد، وأن أحدث سلف مشترك لفيروسات كورونا ألفا يعود إلى 2400 سنة قبل الميلاد ولفيروسات بيتا إلى سنة 3300 سنة قبل الميلاد ولفيروسات غاما إلى 2800 سنة قبل الميلاد ولفيروسات دلتا إلى 3000 سنة قبل الميلاد.[196] لكن بعض العلماء يعتقدون أن هذه الدراسات تبالغ في تقدير معدل التطفر لفيروسات كورونا وتتجاهل تأثير اصطفاء التنقية ويقترحون نموذجا يزعم أن أحدث سلف مشترك لفيروسات كورونا ظهر منذ 55 مليون سنة وخضع إلى تطور مشترك طويل الأمد في الخفافيش والطيور.[127] الخفافيش والطيور مستودع طبيعي مثالي لجينات فيروس كورونا لكونها فقاريات طائرة من ذوات الدم الحار (مع كون الخفافيش مستودعا لفيروسات ألفا وبيتا، والطيور مستودعا لفيروسات غاما ودلتا). مكّن العدد الكبير لأنواع الخفافيش والطيور التي تستضيف الفيروسات والنطاق العالمي لانتشارها من تطور كثيف وانتشار كبير لفيروسات كورونا.[196]
يرجع أصل العديد من فيروسات كورونا البشرية إلى الخفافيش.[197] اشترك فيروس كورونا البشري NL63 في سلف مشترك مع فيروس كورونا الخفاشي(ARCoV.2) بين 1190-1449 م.[198] وتشارك فيروس كورونا البشري 229E مع فيروس كورنا الخفاشي (GhanaGrp1 Bt CoV) في سلف مشترك بين 1686-1800 م.[199] حديثا، تشعب فيروس كورونا الألبكة عن فيروس 229E في وقت ما قبل عام 1960.[200] انتقل ميرس-كوف إلى البشر من الخفافيش عبر الجمال كمضيف وسيط.[201] ومع أن ميرس-كوف له صلة بالعديد من فيروسات كورونا، يبدو أنه تشعب عن هذه الأخيرة منذ عدة قرون.[202] تشعب سارس-كوف عن أقرب فيروسات كورنا الخفاشية صلة به سنة 1986.[203] أصاب أسلاف سارس-كوف أول مرة خفاش الأنف الورقي من عائلة الهيبوسيدرديات ثم انتشرت إلى خفاش حدوة الفرس في عائلة خفاشيات حدوة الفرس، ثم إلى زباد النخيل الآسيوي وأخيرا إلى البشر.[204][205]
خلافا لغيره من فيروسات كورونا بيتا، يرجع أصل فيروس كورونا البقري الذي ينحدر من النوع فيروس كورونا بيتا 1 والجنس الفرعي فيروس أمبيكو إلى القوارض وليس إلى الخفافيش.[197][206] في عقد 1790 تشعب فيروس كورونا الخيلي عن فيروس كورونا البقري بعد انتقال عبر الأنواع.[207] ولاحقا في العقد 1890 تشعب فيروس كورونا البشري OC43 عن فيروس كورونا البقري بعد حادثة انتقال أخرى عبر الأنواع.[207][208] افتُرض أن جائحة الإنفلونزا 1890 قد يكون سببها حادثة انتقال عبر الأنواع، وليس فيروس الإنفلونزا بسبب توقيت حدوثها، الأعراض العصبية والمسبب غير المعروف للجائحة.[209] فضلا عن التسبب في عداوى تنفسية، اشتُبه أن فيروس كورونا البشري OC43 لعب دورا في الأمراض العصبية.[210] في عقد 1950 بدا فيروس OC43 في التشعب إلى أنماطه الجينية الحالية.[211] في علم تطور السلالات، فيروس التهاب الكبد الفأري الذي يصيب كبد الفأر والجهاز العصبي المركزي،[212] له صلة بفيروس كورونا البشري OC43 وفيروس كورونا البقري. فيرروس كورونا البشري HKU1 -كغيره من الفيروسات المذكورة سابقا- له أصول ترجع إلى القوارض كذلك.[197]
إصابة البشر
| ميرس-كوف | سارس-كوف | سارس-كوف-2 | |
|---|---|---|---|
| المرض | ميرس | سارس | كوفيد 19 |
| التفشيات | 2012، 2015، 2018 | 2002-2004 | جائحة 2019-2021 |
| الوبائيات | |||
| تاريخ أول حالة محددة | يونيو 2012 | نوفمبر 2002 | ديسمبر 2019[213] |
| مكان أول حالة محددة | جدة، السعودية | غوانغدونغ، الصين | ووهان، الصين |
| متوسط العمر | 56 | 44[214][arabic-abajed 1] | 56[215] |
| نسبة الجنس (ذ:إ) | 3.3:1 | 0.8:1[216] | 1.6:1[215] |
| الحالات المؤكدة | 2494 | 8096[217] | 603٬525٬541 [218][arabic-abajed 2] |
| الوفيات | 858 | 774[217] | 6٬496٬790 [218][arabic-abajed 2] |
| نسبة الحالات المتوفية | 37% | 9.2% | 1٫08% [218] |
| الأعراض | |||
| الحمى | 98% | 99–100% | 87.9%[219] |
| السعال الجاف | 47% | 29–75% | 67.7%[219] |
| ضيق النفس | 72% | 40–42% | 18.6%[219] |
| الإسهال | 26% | 20–25% | 3.7%[219] |
| التهاب الحلق | 21% | 13–25% | 13.9%[219] |
| استخدام التنفس الاصطناعي | 24.5%[220] | 14–20% | 4.1%[221] |
ملاحظات
| |||
تختلف فيروسات كورونا اختلافا ملحوظا في عامل الخطر حيث يمكن لبعضها أن يقتل أكثر من 30٪ من المصابين مثل ميرس-كوف، والبعض الآخر غير ضار نسبيا ويسبب زكاما خفيفا.[55] يمكن أن تسبب فيروسات كورونا زكاما مصحوبا بأعراض حادة مثل: مثل الحمى والتهاب الحلق بسبب الزوائد الأنفية المنتفخة.[222] يمكن أن تسبب فيروسات كورونا ذات الرئة (إما ذات الرئة الفيروسي المباشر أو ذات رئة بكتيري ثانوي) والتهاب القصبات (إما التهاب قصبات فيروسي مباشر أو التهاب قصبات بكتيري ثانوي).[223] فيروس كورونا البشري المكتشف عام 2003 -سارس-كوف- الذي يسبب متلازمة تنفسية حادة وخيمة (سارس) ، له إمراضية فريدة لأنه يسبب التهابات الجهاز التنفسي العلوي والسفلي.[223]
توجد ستة أنواع معروفة من فيروسات كورونا البشرية، وينقسم نوع واحد إلى سلالتين مختلفتين، ما يجعل العدد الكلي سبع سلالات يمكنها إصابة البشر. أربع سلالات من بين هذه السلالات السبع تتسبب في أعراض خفيفة عموما، ورغم تكيف البشر معها، ربما كانت هذه الفيروسات أكثر حدة في الماضي:[224]
- فيروس كورونا البشري OC43 (HCoV-OC43).
- فيروس كورونا البشري HKU1 (HCoV-HKU1).
- فيروس كورونا البشري 229E (HCoV-229E).
- فيروس كورونا البشري NL63 (HCoV-NL63).
ثلاث فيروسات كورونا يمكن أن تسبب أعراضا حادة:
- فيروس كورونا المرتبط بالمتلازمة التنفسية الحادة الشديدة النوع 1 (سارس-كوف) ويسبب المتلازمة التنفسية الحادة الوخيمة (سارس).
- فيروس كورونا المرتبط بمتلازمة الشرق الأوسط التنفسية (ميرس-كوف) ويسبب متلازمة الشرق الأوسط التنفسية (ميرس).
- فيروس كورونا المرتبط بالمتلازمة التنفسية الحادة الشديدة النوع 2 (سارس-كوف-2) ويسبب مرض فيروس كورونا 2019 (كوفيد 19).
الزكام
يمكن أن تتسبب فيروسات كورونا البشرية (229E ،OC43 ،NL63 ،HKU1) في الإصابة بالزكام،[210] وتنتشر هذه الفيروسات الأربعة على نطاق واسع بين السكان، وتتسبب في ظهور أعراض خفيفة. تشير الإحصائيات إلى أن 15٪ من حالات الزكام تسببها هذه الفيروسات الأربعة (وتشير بيانات أخرى إلى 20٪-30٪ [225][226]) في حين أن 50% من الحالات يسببها الفيروس الأنفي.[227] في المناطق ذات المناخ المعتدل، تحدث الإصابة بهذه الفيروسات غالبا في فصل الشتاء،[228][229] ولا يوجد فرق كبير في أوقات الإصابة في مناطق المناخ المداري.[230] قد تتنافس فيروسات كورونا الأربعة مع بعضها البعض ويكون انتشارها بين السكان دوريا.[225] مع أن فيروسات كورونا هذه تسبب الزكام فقط، فقد افتُرض أنها ربما تسببت في أوبئة حادة عندما ظهرت لأول مرة منذ قرون أو عقود مضت، ثم تطورت بمرور الوقت وتأقلم عليها البشر وأصبحت هذه السلالات أقل إمراضا.[225] مثل فيروس كورونا البشري OC43 وفيروس كورونا البقري اللذان تشعبا عن النوع فيروس كورونا بيتا 1 في العقد 1890، حيث انتقل الفيروس البقري عبر الأنواع واكتسب القدرة على إصابة البشر، [207][208] وعلى هذا الأساس افترض بعض العلماء أن جائحة الإنفلونزا (1889–1890) قد يكون سببها فيروس OC43 الناتج من هذا الانتقال عبر الأنواع وليس فيروس الأنفلونزا.[231]
المتلازمة التنفسية الحادة الوخيمة (سارس)

في نهاية عام 2002 ظهرت ذات رئة غير نموذجية في مقاطعة غوانغدونغ بالصين، وتطورت لاحقا إلى تفشي السارس 2002-2004 الذي استمر حوالي عام تقريبا. انتشر الوباء إلى 29 دولة وأصاب أزيد من 8000 شخص توفي منهم 774 فردا.[232][233] في مارس 2003 سمت منظمة الصحة العالمية بشكل رسمي المرض باسم السارس، ووجد العديد من العلماء أن الوباء سببه أحد فيروس كورونا وكان أول فيروس كورونا يسبب مرضا خطيرا. أرجع العلماء مصدر الفيروس المباشر إلى زباد النخل الذي بيع في الأسواق، وكان تماثل التسلسل بين فيروس سارس-كوف زباد النخل وفيروس السارس البشري 99.8%،[234] لكن المستودع الطبيعي لهذا الفيروس يُفترض أن يكون أحد الخفافيش مثل: خفاش حدوة الحصان الصيني الأصهب أو خفاش الحذوة الكبير.[235][236] فترة حضانة فيروس السارس حوالي 4 إلى 6 أيام وبعدها تظهر أعراض مماثلة للإنفلونزا والالتهاب الرئوي. في الحالات الحادة يمكن أن يعاني المرضى من صعوبات في التنفس وقد يتطور الأمر إلى متلازمة الضائقة التنفسية الحادة (ARDS). تصبح أعراض الإصابة أكثر حدة بعد أن يبدأ جهاز المناعة في محاربة الفيروس حيث تبين أن بعض الأعراض يسببها فرط الاستجابة المناعية الخاصة بالمريض.[10] بعض مرضى السارس مازالوا يعانون من العديد من المضاعفات مثل: التليف الرئوي، هشاشة العظام والنخر اللاوعائي بعد الشفاء، وتطور لدى مرضى آخرين اضطراب الكرب التالي للصدمة النفسية أو اضطراب مزاجي.[237]
متلازمة الشرق الأوسط التنفسية (ميرس)

في 2012 ظهر فيروس كورونا جديد سُمي لاحقا بفيروس كورونا متلازمة الشرق الأوسط التنفسية (ميرس-كوف)، حيث تم عزله من رئة مريض مصاب بأعراض تنفسية حادة في المملكة العربية السعودية.[238][239] ويُعتقد أنه انتقل إلى البشر من الجمال،[240] حيث وجدت دراسة أن فيروس كورونا الذي أصاب الجمال العربية البالغة وصغارها مطابق بنسبة 99.9% لجينومات ميرس-كوف البشرية.[241] أما أصل هذا الفيروس ومستودعه الطبيعي فيُعتقد أنه يرجع إلى الخفافيش،[242] وأن له صلة وثيقة بفيروس كورونا خفاش تيلونيكترايس HKU4 وفيروس كورونا خفاش بيبيسترالوس HKU5.[243] اعتبارا من يناير 2021، أُبلغ عن 2500 حالة منها 45 حالة في 2020.[244] وتوفي حوالي 35% من الأفراد الذين أصيبوا بالفيروس.[240] وفضلا عن التفشي الأول عام 2012 حدثت تفشيات أخرى أحدها في كوريا الجنوبية عام 2015 وآخر في السعودية عام 2018.[240][245] فترة حضانة فيروس ميرس حوالي 5.5 يوم،[246] ويسبب أعراضا منها الحمى، السعال، الإسهال وضيق التنفس.[240] بعض المرضى لا تظهر عليهم الأعراض، والحالات الخطيرة يمكن أن تسبب ذات رئة وخيمة ومتلازمة الضائقة التنفسية الحادة.[247] الفيروس معدي نسبيا، وفي الغالب يصيب الأفراد الذين هم في اتصال مباشر مع المرضى، ولا ينتقل غالبا خارج المستشفى.[248]
مرض فيروس كورونا 2019 (كوفيد 19)
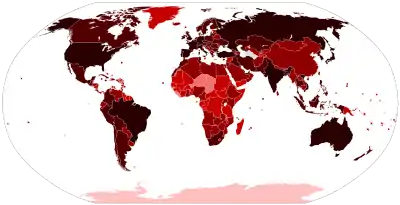
في ديسمبر 2019، حدث تفشي التهاب رئوي جديد في مدينة ووهان بمقاطعة خوبي بالصين. ووجد العلماء في 31 ديسمبر أن التفشي سببه سلالة جديدة من فيروسات كورونا،[249] والتي سميت لاحقا سارس-كوف-2 بواسطة اللجنة الدولية لتصنيف الفيروسات. يملك سارس-كوف-2 تماثل تسلسل قدره 79% مع سارس-كوف.[250] ويُعتقد أن الخفافيش هي المستودع الطبيعي الأرجح له،[251][252] حيث يملك RaTG13 تسلسلا متماثلا بنسبة 96.1% والفيروس الجديد المكتشف في اللاوس BANAL-52 نسبة 96.8%،[253] توحي نسبة التماثل بين هذه الفيروسات الخفاشية وسارس-كوف-2 بأن البشر قد أصيبوا بالفيروس عبر مضيف وسيط،[254] لكن بقي هذا الوسيط وهو المصدر المباشر لإصابة البشر غير معروف.[255][256]
اعتبارا من 1 سبتمبر 2022 يوجد على الأقل 6٬496٬790 [218] حالة وفاة مؤكدة وأزيد من 603٬525٬541 [218] إصابة مؤكدة بسبب جائحة كوفيد 19، التي كان لها تأثير كبير على المجتمع والإقتصاد العالمي وسببت أسوء ركود اقتصادي منذ الكساد الكبير في عقد 1930.[257] أعراض كوفيد 19 متنوعة وتختلف من فرد لآخر، ومدة حضانة فيروس سارس-كوف-2 تتراوح بين 1 إلى 14 يوما. على الأقل ثلث الأفراد المصابين لا تظهر عليهم الأعراض، لكن بإمكانهم نقل الفيروس،[258][259] معظم أعراض كوفيد 19 خفيفة أو متوسطة ومن بينها: الحمى، السعال، الإعياء، فقدان الشم المؤقت وذات الرئة الخفيفة. تعاني نسبة صغيرة من المرضى من أعراض حادة منها: ضيق التنفس ونقص التأكسج، أما الحالات الخطرة فقد تعاني من فشل تنفسي، صدمة وفشل الأعضاء المتعدد.[260] بعض المرضى تبقى لديهم أعضاء متضررة بعد الشفاء تفشل في التعافي التام، ويعانون من مضاعفات ما بعد الإصابة لأمد طويل.[261]
إصابة الحيوانات
عُرِف أن فيروسات كورونا تسبب حالات مرضية في الطب البيطري منذ ثلاثينيات القرن الماضي،[25] وهي تصيب مجموعة متنوعة من الحيوانات منها: الخنازير والأبقار والخيول والجمال والقطط والكلاب والقوارض والطيور والخفافيش.[262] تصيب معظم فيروسات كورونا المرتبطة بالحيوانات السبيل الهضمي بواسطة الطريق الفموي الشرجي.[263] تم تركيز جهود بحثية معتبرة لمعرفة الإمراضية الفيروسية الخاصة بفيروسات كورونا الحيوانية هذه، وخاصة من قبل علماء الفيروسات المهتمين بالأمراض البيطرية والحيوانية.[264]
الدواجن والماشية

يمكن لفيروسات كورونا أن تصيب أنواعا مختلفة من الدواجن والماشية. فيروس التهاب الشعب الهوائية المعدي (IBV) هو فيروس كورونا غاما يصيب الدواجن عبر الهباء الجوي أو الطريق الفموي الشرجي،[265] ويسبب التهابا شعبيا معديا ويصيب الجهاز البولي التناسلي فضلا عن السبيل التنفسي.[266] يملك فيروس التهاب الشعب المعدي معدل نفوق مرتفع، وانتشارا سريعا،[262] ويؤثر على إنتاج البيض ويسبب خسائر اقتصادية كبيرة، وتوجد العديد من اللقاحات للوقاية منه.[267] بعض السلالات الشبيهة بـIBV يمكنها أن تصيب الدجاج الرومي وتسبب التهاب الأمعاء منها فيروس كورونا الدجاج الرومي (TCV).[262] يُصنف فيروس التهاب الشعب المعدي إلى جانب مختلف سلالات الدجاج الرومي وبعض الفيروسات التي تصيب الطيور البرية إلى نوعين: فيروس كورونا الطيري، وفيروس كورونا الطيري 9203 تحت جنس فيروسات كورونا غاما.[190][195]
تصاب الماشية مثل الخنازير والأبقار والأحصنة بفيروسات كورونا كذلك.[262] من بين فيروسات كورونا التي تصيب الخنازير: فيروس الإسهال الوبائي الخنزيري (PEDV)،[268] فيروس التهاب المعدة والأمعاء الساري الخنزيري (TGEV)،[192][269] فيروس كورونا دلتا الخنزيري (PdCV)، فيروس التهاب الدماغ والنخاع الشوكي الراصي الخنزيري (PHEV) وفيروس كورونا متلازمة الإسهال الحاد الخنزيري (SADS-CoV). باستثناء PHEV الذي يصيب النسيج العصبي كذلك، تصيب فيروسات كورونا الخنزيرية الجهاز الهضمي وتسبب القيء والإسهال وأعراضا أخرى. فيروسا TGEV وPHEV متنشران بين الخنازير لأزيد من 10 سنوات أما SADS-CoV وPdCV فهما فيروسان حديثا النشأة،[270] ورغم أن PEDV متواجد منذ 1970 إلا أن سلالة شديدة الإمراض ظهرت في 2010 وسببت خسائر ملحوظة في تربية الخنازير في بلدان مختلفة.[271] من بين الفيروسات الناشئة، تسلسل SADS-CoV مماثل جدا لفيروس كورونا خفاش حدوة الفرس الصيني HKU2،[272] وتسلسل PEDV مماثل لفيروس كورونا خفاش إلف الظلام 512 وقد يرجع أصلهما إلى انتقال عبر الأنواع من الخفافيش.[273] ربما يرجع أصل PdCV إلى فيروسات كورونا الطيرية فتسلسله متماثل جدا مع فيروس كورونا الدوري HKU17 وفيروس كورونا السمان HKU30 ويُفترض أنهما ينتميان إلى نفس النوع.[274]
فيروس كورونا البقري الذي يصيب الأبقار قريب الصلة بفيروس كورونا البشري OC43،[275] ويسبب أعراضا مثل الإسهال لدى العجول.[262] يسبب فيروس كورونا الفرسي الذي يصيب الخيول الإسهال كذلك،[276] ونادرا ما يكون قاتلا.[277]
الحيوانات الأليفة

تصاب الحيوانات الأليفة مثل: القطط والكلاب وابن مقرض بفيروسات كورونا كذلك.[265] ينتمي فيروس كورونا السنوري إلى فيروسات ألفا[278] ويمكن أن يُقسم إلى نوعين: فيروس كورونا القططي المعوي (FECV) الذي يصيب الأمعاء ويسبب أعراضا خفيفة، وفيروس التهاب البريتون المعدي السنوري (FIPV) الذي يمكن أن يسبب التهاب البريتون المعدي السنوري الحاد.[262] ينتقل FECV عبر الطريق الشرجي الفموي ويمكن أن يتطفر إلى FIPV بعد الإصابة، ثم ينتقل لإصابة مختلف أعضاء القطط.[279] معدل النفوق بالتهاب البريتون المعدي السنوري مرتفع وعادة ما تموت القطط خلال عام واحد من ظهور الأعراض.[280]
معروف بأن اثنين من فيروسات كورونا تصيب الكلاب: فيروس كورونا الكلبي (CCoV) وفيروس كورونا التنفسي الكلبي (CRCoV). الأول من فيروسات كورونا ألفا[278] ويسبب أعراضا معدية معوية خفيفة،[262] أما الثاني فهو من فيروسات كورونا بيتا[275] ويصيب الجهاز التنفسي وهو أحد الممرضات التي تسبب سعال الكلاب.[262] يوجد كذلك نوعان من فيرووسات كورونا التي تصيب ابن مقرض:[281] فيروس كورونا ابن مقرض ويسبب التهاب الأمعاء، وفيروس كورونا جهازي ابن مقرضي (FSC) وهو أشد خطرا ويسبب مرض جهازي مماثل لالتهاب البريتون المعدي السنوري.[282][283]
الحيوانات المختبرية
يمكن لفيروسات كورونا أن تصيب العديد من الحيوانات المستخدمة في المختبرات.[262] فيروس التهاب الكبد الفأري (MHV) كان أكثر فيروس كورونا تمت دراسته حيويا ومخبريا وعلى المستوى الجزيئي قبل اكتشاف سارس-كوف،[284] ويصيب صغار فئران التجارب مسببا معدلات نفوق عالية،[285] بينما يسبب أعراضا خفيفة قابلة للعكس لدى الفئران البالغة، لكنه يثير استجابة مناعية يمكن أن تؤثر على تفسير النتائج التجريبية.[286] ينقسم MHV إلى عدة سلالات وتتنوع الأنسجة التي يصيبها وحدة الأعراض التي يسببها. يسبب MHV-1 أعراضا تنفسية وتسبب السلالتان A59 وMHV-3 التهاب الكبد ويسبب JHV التهاب الدماغ. يحفز الفيروس الأخير استجابات مناعية لدى الفئران تسبب أضرارا لأغمدة الميالين في العصبونات، لذلك يُستخدم كنموذج حيواني لدراسة التصلب المتعدد.[55] من فيروسات كورونا التي تصيب الجرذان: فيروس التهاب الغدد اللعابية والدمعية (SDAV) وفيروس كورونا باركر الجرذي (RCoV-P)، يمكن أن تصيب السلالة الأولى الأعين والغدد الدمعية والغدد اللعابية ويمكن كذلك أن تؤثر على تكاثر الجرذان وتسبب تغيرات سلوكية وهي أحد أهم الممرضات لجرذان التجارب.[287] تصنف هاتان السلالتان الجرذيتان مع مختلف سلالات MHV الفأرية كنوع واحد ويشار إليهم جميعا باسم فيروس كورونا الموريني.
يوجد فيروس كورونا يمكن أن يصيب الأرانب ويسبب التهاب الأمعاء مع معدلات نفوق عالية، ويسمى فيروس كورونا المعوي الأرنبي (RECV) وينتقل عبر الطريق الشرجي الفموي.[289][290] فضلا عن ذلك، تُصاب الأرانب المستخدمة لزراعة اللولبية الشاحبة بممرضات غير معروفة يفترض أن تكون فيروس كورونا، وتوجد حالات قليلة فقط لإصابة الأرانب بهذين النوعين من فيروس كورونا.[291][292] في 2012، وجد بعض الباحثين فيروس كورونا الأرنبي HKU14 في الأرانب التي بيعت في الأسواق، وهو فيروس بيتا يصيب الأرانب من دون أعراض ظاهرة.[292] توجد كذلك تقارير حول إصابة فيروس كورونا للكابياء الخنزيرية وتسببه في أعراضٍ مثل الإسهال والتهاب الأمعاء، لكن مدى انتشار الفيروس في أوساط الكابياء الخنزيرية وعلاقته بفيروسات كورونا الأخرى مازالت غير معروفة.[291]
الحيوانات البرية
زيادة على الحيوانات سالفة الذكر التي يربيها الإنسان، توجد العديد من سلالات فيروس كورونا التي تصيب الطيور والثدييات البرية. بعد حدث السارس في عام 2003، اكتُشِف عدد كبير من فيروسات كورونا ألفا وبيتا في مجموعة متنوعة من الخفافيش البرية، ويرتبط العديد منها ارتباطا وثيقا بسلالات سارس-كوف وميرس-كوف وسارس-كوف-2 التي تسببت في التفشيات.[293] وُجدت العديد من سلالات هذين الجنسين في ثدييات برية صغيرة مثل: القوارض وأرنبيات الشكل والقنافذ والزبابات.[293][294] فضلا عن ذلك، وُجد في مجترات مثل: الرنة والغزلان والبيسون والظباء والزرافات سلالات فيروس كورونا مماثلة لفيروس كورونا البقري،[295] ووجد كذلك لدى الثدييات البحرية مثل الفقميات والحيتانيات فيروسات من الجنس ألفا وغاما على التوالي (فيروس كورونا الحوتي)[296] أما لدى الطيور البرية فقد عُثر على سلالات كثيرة من غاما ودلتا، وصُنف العديد من سلالات غاما تحت النوع الجديد فيروس كورونا البطي 2714. معظم الأنواع في جنس دلتا تصيب الطيور البرية.[195] بالإضافة إلى الثدييات والطيور، وُجدت سلالةٌ لفيروس كورونا تصيب الزواحف (السحالي الصينية).[297]
الوقاية

بين الأمراض البشرية، لا توجد لقاحات لفيروسات كورونا البشرية الأربعة التي تسبب الزكام، وسبيل تجنب الإصابة بها هو غسل الأيادي وتجنب لمس الأعين والأنف والفم وارتداء الكمامات.[298] اعتبارا من 2021، مازال لا يوجد لقاح فعال للوقاية من السارس،[299][300] وميرس،[301] لكن بعض اللقاحات ضد ميرس هي قيد التطوير.[240] السارس مرض ينتقل عبر الرذاذ التنفسي ويمكن لقناع N95 منع الإصابة به بفعالية،[302] وبسبب قوة الإعداء الكبيرة لمرضى الحالات الحادة فإن العزل والحجر الصحي إجراء فعال من أجل الحد من انتشاره، والمرضى الذين يتم عزلهم قبل اليوم الخامس من الإصابة نادرا ما يقومون بنقل الفيروس للآخرين.[303]
فضلا عن الحفاظ على النظافة الشخصية وعزل المرضى، من إجراءات منع انتشار كوفيد 19: الحرص على التباعد الاجتماعي، إلغاء التجمعات غير الضرورية، العمل والدراسة عبر الإنترنيت ومنع انتشار الجائحة عبر تقييد السفر.[304] طُورت العديد من اللقاحات لكوفيد 19 وتنصح منظمة الصحة العالمية الأفراد بأخذ جميع الجرعات الموصى بها بواسطة الهيئات الصحية التي يتبعون لها بمجرد أن يحين دورهم بما في ذلك جرعة معززة إذا تمت التوصية بها.[305]
لدى الحيوانات، تطوير اللقاحات ضد فيروسات كورونا صعب لأنها عرضة للتأشيب الجيني، ويمكن للقاحات أن تعزز حدوث طفرات أخرى. لا يمكن الوقاية من التهاب البريتون المعدي السنوري الذي تسببه الإصابة بفيروس كورونا السنوري باللقاحات، واللقاحات المطورة ضده في الماضي لم تفشل في الوقاية منه وحسب بل تسببت في تفاقم الأعراض.[154] حاليا، توجد لقاحات يمكن أن تقي من فيروس التهاب الشعب الهوائية المعدي (IBV)، فيروس التهاب المعدة والأمعاء الساري (TGEV)، وفيروس الإسهال الوبائي الخنزيري (PEDV) وفيروس كورونا الكلبي، لكن تأثيرها الوقائي غير تام ويمكن أن تسبب تأشيبا جينيا لتوليد سلالات جديدة، خضع فيروس التهاب المعدة والأمعاء الساري إلى طفرة حذف تسلسل ونتجت سلالة متطفرة تسمى فيروس كورونا التنفسي الخنزيري (PRCV) تسبب أعراضا خفيفة ونجح التلقيح بهذه السلالة الموهنة من الوقاية ضد تفشيات جديدة حادة لـTGEV خلال الـ30 عاما الماضية.[154][306]
ملاحظات
- مغم 5' هي اختصار للحروف الأولى من المصطلح "مـنطقة غـير مـترجمة 5'"، وتقابل الاختصار الأجنبي (5′ UTR) للمصطلح 5′ UnTranslated Region
- في فيروس سارس-كوف يمكن لبروتيني ORF3a وORF8 أن يكملا وظيفة هذا البروتين الغلافي.[77]
- يوجد استثناء يخص نطاق النهاية الأمينية لفيروس كورونا الفأري الذي يرتبط بالمستقبل البروتيني: جزيء التصاق الخلية المرتبط بالمستضد السرطاني المضغي 1 (CEACAM1)
- ما إذا كان ميرس-كوف يُقص بواسطة الفيورين مازال أمرا خلافيا، وبعض الدراسات تشكك في دور الفيورين في عملية إصابة ميرس-كوف للخلايا.[87][88]
- فيروسات ميرس-كوف التي تُنتج في خلايا لا تحتوي على الفيورين، لا يتم قصها في S1/S2 وتتغير هيئتها ببطء بعد الارتباط بالخلية وتتطلب مسار جسيم داخلي-جسيم حال من أجل الدخول إلى الخلية، ولا يمكنها الدخول إلى السيتوبلازم سوى بعد أن تُقص بواسطة الكاتبسين في الجسيم الحال.[90]
المراجع
باللغة الإنجليزية
- المحرر: اللجنة الدولية لتصنيف الفيروسات — العنوان : ICTV Master Species List 2018a v1 — العمل الكامل مُتوفِّر في: https://talk.ictvonline.org/files/master-species-lists/m/msl/7992
- المحرر: اللجنة الدولية لتصنيف الفيروسات — العنوان : ICTV Master Species List 2018b.v2 — العمل الكامل مُتوفِّر في: https://talk.ictvonline.org/files/master-species-lists/m/msl/8266
- Virus Taxonomy: 2018b Release (محفوظ) (بالإنجليزية). (html) (مارس 2019).
- 2017.012-015S (محفوظ) (بالإنجليزية). (xlsx) (أكتوبر 2018).
- Cherry, James؛ Demmler-Harrison؛ Kaplan؛ Steinbach؛ Hotez (29 ديسمبر 2017)، Feigin and Cherry's Textbook of Pediatric Infectious Diseases E-Book، Elsevier Health Sciences، ص. PT6615، ISBN 978-0-323-39281-5، مؤرشف من الأصل في 6 أبريل 2022.
- Fan, Yi؛ Zhao؛ Shi؛ Zhou (02 مارس 2019)، "Bat Coronaviruses in China"، Viruses، 11 (3): 210، doi:10.3390/v11030210، ISSN 1999-4915، PMC 6466186، PMID 30832341، مؤرشف من الأصل في 9 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Woo, Patrick C. Y.؛ Huang؛ Lau؛ Yuen (24 أغسطس 2010)، "Coronavirus Genomics and Bioinformatics Analysis"، Viruses، 2 (8): 1804–1820، doi:10.3390/v2081803، ISSN 1999-4915، PMC 3185738، PMID 21994708، مؤرشف من الأصل في 7 مايو 2022،
Coronaviruses possess the largest genomes [26.4 kb (ThCoV HKU12) to 31.7 kb (SW1)] among all known RNA viruses (Figure 1) [2,13,16].
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Snijder, Eric J.؛ Limpens؛ de Wilde؛ de Jong؛ Zevenhoven-Dobbe؛ Maier؛ Faas؛ Koster؛ Bárcena (08 يونيو 2020)، Cimarelli, Andrea (المحرر)، "A unifying structural and functional model of the coronavirus replication organelle: Tracking down RNA synthesis"، PLOS Biology، Public Library of Science (PLoS)، 18 (6): e3000715، doi:10.1371/journal.pbio.3000715، ISSN 1545-7885.
- Fung, To Sing؛ Liu (08 سبتمبر 2019)، "Human Coronavirus: Host-Pathogen Interaction"، Annual Review of Microbiology، 73 (1): 529–557، doi:10.1146/annurev-micro-020518-115759، ISSN 0066-4227، مؤرشف من الأصل في 11 مايو 2022.
- International Committee on Taxonomy of Viruses (ICTV) (محفوظ) (بالإنجليزية). مؤرشف من الأصل في 12 مايو 2022. وصل لهذا المسار في 4 أبريل 2022.
- Definition of Coronavirus by Merriam-Webster (محفوظ). مؤرشف من الأصل في 23 مارس 2020. وصل لهذا المسار في 24 مارس 2020.
- Definition of Corona by Merriam-Webster (محفوظ). مؤرشف من الأصل في 24 مارس 2020. وصل لهذا المسار في 24 مارس 2020.
- Tyrrell, Fielder (2002)، Cold Wars: The Fight Against the Common Cold (باللغة الإنجليزية)، Oxford University Press، ص. 96، ISBN 978-0-19-263285-2، مؤرشف من الأصل في 7 أبريل 2022،
We looked more closely at the appearance of the new viruses and noticed that they had a kind of halo surrounding them. Recourse to a dictionary produced the Latin equivalent, corona, and so the name coronavirus was born.
- "Virology: Coronaviruses"، Nature، 220 (5168): 650، نوفمبر 1968، Bibcode:1968Natur.220..650.، doi:10.1038/220650b0، PMC 7086490،
[T]here is also a characteristic "fringe" of projections 200 A long, which are rounded or petal shaped ... This appearance, recalling the solar corona, is shared by mouse hepatitis virus and several viruses recently recovered from man, namely strain B814, 229E and several others.
- "The molecular biology of coronaviruses"، Advances in Virus Research، 28: 35–112، 01 يناير 1983، doi:10.1016/s0065-3527(08)60721-6، ISBN 9780120398287، PMC 7131312، PMID 6362367،
[T]hese viruses displayed a characteristic fringe of large, distinctive, petal-shaped peplomers or spikes which resembled a crown, like the corona spinarum in religious art; hence the name coronaviruses.
- Lalchhandama K (2020)، "The chronicles of coronaviruses: the bronchitis, the hepatitis and the common cold"، Science Vision (باللغة الإنجليزية)، 20 (1): 43–53، doi:10.33493/scivis.20.01.04.
- "Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2009)"، Archives of Virology، 155 (1): 133–46، 2010، doi:10.1007/s00705-009-0547-x، PMC 7086975، PMID 19960211.
- International Committee on Taxonomy of Viruses (ICTV) (محفوظ) (بالإنجليزية). مؤرشف من الأصل في 12 مايو 2022. وصل لهذا المسار في 14 سبتمبر 2020.
- "The morphology of three previously uncharacterized human respiratory viruses that grow in organ culture"، The Journal of General Virology، 1 (2): 175–8، أبريل 1967، doi:10.1099/0022-1317-1-2-175، PMID 4293939.
- "Coronaviruses, a New Group of Animal RNA Viruses"، Avian Diseases، 14 (2): 330–336، 1970، doi:10.2307/1588476، ISSN 0005-2086، JSTOR 1588476، PMID 4316767.
- Fabricant J (1998)، "The Early History of Infectious Bronchitis"، Avian Diseases، 42 (4): 648–650، doi:10.2307/1592697، ISSN 0005-2086، JSTOR 1592697، PMID 9876830.
- "Laryngotracheitis in chicks"، Poultry Science (باللغة الإنجليزية)، 12 (1): 55–60، 1933، doi:10.3382/ps.0120055.
- Decaro N (2011)، "Gammacoronavirus"، في Tidona C, Darai G (المحررون)، Gammacoronavirus‡: Coronaviridae، The Springer Index of Viruses (باللغة الإنجليزية)، Springer، ص. 403–413، doi:10.1007/978-0-387-95919-1_58، ISBN 978-0-387-95919-1، PMC 7176155.
- "Coronaviruses: A Comparative Review"، Current Topics in Microbiology and Immunology / Ergebnisse der Mikrobiologie und Immunitätsforschung، Current Topics in Microbiology and Immunology / Ergebnisse der Mikrobiologie und Immunitätsforschung (باللغة الإنجليزية)، Berlin, Heidelberg: Springer، 1974، ص. 87، doi:10.1007/978-3-642-65775-7_3، ISBN 978-3-642-65775-7.
- / "Il était une fois les coronavirus"، Réalités Biomédicales (باللغة الفرنسية)، 27 مارس 2020، مؤرشف من الأصل في 20 فبراير 2022، اطلع عليه بتاريخ 18 أبريل 2020.
{{استشهاد بخبر}}: تحقق من قيمة|مسار أرشيف=(مساعدة) - "History and recent advances in coronavirus discovery"، The Pediatric Infectious Disease Journal، 24 (11 Suppl): S223–7, discussion S226، نوفمبر 2005، doi:10.1097/01.inf.0000188166.17324.60، PMID 16378050.
- "The BMJ in 1965"، BMJ، 369: m1547، أبريل 2020، doi:10.1136/bmj.m1547، PMID 32299810، مؤرشف من الأصل في 24 أبريل 2022.
- Monto AS (1984)، "Coronaviruses"، في Evans AS (المحرر)، Viral Infections of Humans، Viral Infections of Humans: Epidemiology and Control (باللغة الإنجليزية)، Springer US، ص. 151–165، doi:10.1007/978-1-4684-4727-9_7، ISBN 978-1-4684-4727-9.
- "Virus isolations from common colds occurring in a residential school"، British Medical Journal، 2 (5297): 82–6، يوليو 1962، doi:10.1136/bmj.2.5297.82، PMC 1925312، PMID 14455113.
- Richmond C (18 يونيو 2005)، "David Tyrrell"، BMJ : British Medical Journal، 330 (7505): 1451، doi:10.1136/bmj.330.7505.1451، PMC 558394.
- "Obituary Notices: Malcom Byone"، British Medical Journal (باللغة الإنجليزية)، 2 (5660): 827–829، 28 يونيو 1969، doi:10.1136/bmj.2.5660.827، S2CID 220187042، مؤرشف من الأصل في 31 مارس 2022.
- "Cultivation of a Novel Type of Common-Cold Virus in Organ Cultures"، British Medical Journal، 1 (5448): 1467–70، يونيو 1965، doi:10.1136/bmj.1.5448.1467، PMC 2166670، PMID 14288084.
- Tyrrell, Fielder (2002)، Cold Wars: The Fight Against the Common Cold (باللغة الإنجليزية)، Oxford University Press، ص. 93–95، ISBN 978-0-19-263285-2، مؤرشف من الأصل في 27 مارس 2022.
- Hagan, Bruner, Gillespie, Timoney, Scott, Barlough (1988)، Hagan and Bruner's Microbiology and Infectious Diseases of Domestic Animals: With Reference to Etiology, Epizootiology, Pathogenesis, Immunity, Diagnosis, and Antimicrobial Susceptibility (باللغة الإنجليزية)، Cornell University Press، ص. 440، ISBN 978-0-8014-1896-9، مؤرشف من الأصل في 5 فبراير 2022.
- Knapp، Alex. The Secret History Of The First Coronavirus (محفوظ) (بالإنجليزية). مؤرشف من الأصل في 28 فبراير 2022. وصل لهذا المسار في 6 مايو 2020.
- "A new virus isolated from the human respiratory tract"، Proceedings of the Society for Experimental Biology and Medicine، 121 (1): 190–3، يناير 1966، doi:10.3181/00379727-121-30734، PMID 4285768، S2CID 1314901.
- "The woman who discovered the first coronavirus"، BBC News، 14 أبريل 2020، مؤرشف من الأصل في 28 أبريل 2022.
- Almeida J (26 يونيو 2008)، "June Almeida (née Hart)"، BMJ (باللغة الإنجليزية)، 336 (7659): 1511.1–1511، doi:10.1136/bmj.a434، ISSN 0959-8138، PMC 2440895.
- "Growth in suckling-mouse brain of "IBV-like" viruses from patients with upper respiratory tract disease"، Proceedings of the National Academy of Sciences of the United States of America، 58 (6): 2268–73، ديسمبر 1967، Bibcode:1967PNAS...58.2268M، doi:10.1073/pnas.58.6.2268، PMC 223830، PMID 4298953.
- "Recovery in tracheal organ cultures of novel viruses from patients with respiratory disease"، Proceedings of the National Academy of Sciences of the United States of America، 57 (4): 933–40، أبريل 1967، Bibcode:1967PNAS...57..933M، doi:10.1073/pnas.57.4.933، PMC 224637، PMID 5231356.
- Times, Harold M. Schmeck Jr Special To the New York (05 مايو 1967)، "Six Newly Discovered Viruses May Explain Cold; Strains Are Similar to Germ That Causes a Bronchial Infection in Chickens Believed to Be New Group"، The New York Times (باللغة الإنجليزية)، ISSN 0362-4331، مؤرشف من الأصل في 8 مايو 2022، اطلع عليه بتاريخ 25 أبريل 2020.
- Myint SH (1995)، "Human Coronavirus Infections"، في Siddell SG (المحرر)، The Coronaviridae، The Viruses (باللغة الإنجليزية)، Springer US، ص. 389–401، doi:10.1007/978-1-4899-1531-3_18، ISBN 978-1-4899-1531-3.
- "Human coronaviruses: insights into environmental resistance and its influence on the development of new antiseptic strategies"، Viruses، 4 (11): 3044–68، نوفمبر 2012، doi:10.3390/v4113044، PMC 3509683، PMID 23202515.
- "Antibodies against MERS coronavirus in dromedary camels, Kenya, 1992-2013"، Emerging Infectious Diseases، 20 (8): 1319–22، أغسطس 2014، doi:10.1007/978-1-4899-7448-8_10، ISBN 978-1-4899-7447-1، PMC 7122465، PMID 25075637،
The other OC strains and B814 that could not be adapted to mouse brain resisted adaptation to cell culture as well; these distinct viruses have since been lost and may actually have been rediscovered recently.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "A Novel Coronavirus from Patients with Pneumonia in China, 2019"، The New England Journal of Medicine، 382 (8): 727–733، فبراير 2020، doi:10.1056/NEJMoa2001017، PMC 7092803، PMID 31978945.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Family Coronaviridae"، Ninth Report of the International Committee on Taxonomy of Viruses، Oxford: Elsevier، 2011، ص. 806–28، doi:10.1016/B978-0-12-384684-6.00068-9، ISBN 978-0-12-384684-6، S2CID 212719285.
- "Ultrastructural characterization of SARS coronavirus"، Emerging Infectious Diseases، 10 (2): 320–6، فبراير 2004، doi:10.3201/eid1002.030913، PMC 3322934، PMID 15030705،
Virions acquired an envelope by budding into the cisternae and formed mostly spherical, sometimes pleomorphic, particles that averaged 78 nm in diameter (Figure 1A).
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "The molecular biology of coronaviruses"، Advances in Virus Research، 66: 193–292، 2006، doi:10.1016/S0065-3527(06)66005-3، ISBN 9780120398690، PMC 7112330، PMID 16877062.
- "The chronicles of coronaviruses: the electron microscope, the doughnut, and the spike"، Science Vision، 20 (2): 78–92، 2020، doi:10.33493/scivis.20.02.03.
- "A structural analysis of M protein in coronavirus assembly and morphology"، Journal of Structural Biology، 174 (1): 11–22، أبريل 2011، doi:10.1016/j.jsb.2010.11.021، PMC 4486061، PMID 21130884،
See Figure 10.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "The molecular biology of coronaviruses"، Advances in Virus Research، 48: 1–100، 1997، doi:10.1016/S0065-3527(08)60286-9، ISBN 9780120398485، PMC 7130985، PMID 9233431.
- Godet M, L'Haridon R, Vautherot JF, Laude H (1992)، "TGEV corona virus ORF4 encodes a membrane protein that is incorporated into virions."، Virology، 188 (2): 666–75، doi:10.1016/0042-6822(92)90521-p، PMC 7131960، PMID 1316677، مؤرشف من الأصل في 14 مارس 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: أسماء متعددة: قائمة المؤلفون (link) - Alsaadi؛ Jones (أبريل 2019)، "Membrane binding proteins of coronaviruses"، Future Virology، 14 (4): 275–286، doi:10.2217/fvl-2018-0144، PMC 7079996، PMID 32201500.
- Fehr, Anthony R.؛ Perlman (2015)، "Coronaviruses: An Overview of Their Replication and Pathogenesis"، في Maier, Helena Jane (المحرر)، Coronaviruses، New York, NY: Springer New York، ج. 1282، ص. 1–23، doi:10.1007/978-1-4939-2438-7_1، ISBN 978-1-4939-2437-0، PMC 4369385، PMID 25720466، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بكتاب}}: صيانة CS1: تنسيق PMC (link) - "Membrane Protein of Human Coronavirus NL63 Is Responsible for Interaction with the Adhesion Receptor"، Journal of Virology، 93 (19)، أكتوبر 2019، doi:10.1128/JVI.00355-19، PMC 6744225، PMID 31315999.
- "Supramolecular architecture of severe acute respiratory syndrome coronavirus revealed by electron cryomicroscopy"، Journal of Virology، 80 (16): 7918–28، أغسطس 2006، doi:10.1128/JVI.00645-06، PMC 1563832، PMID 16873249،
Particle diameters ranged from 50 to 150 nm, excluding the spikes, with mean particle diameters of 82 to 94 nm; Also See Figure 1 for double shell.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - Brant, Ayslan Castro؛ Tian؛ Majerciak؛ Yang؛ Zheng (19 يوليو 2021)، "SARS-CoV-2: from its discovery to genome structure, transcription, and replication"، Cell & Bioscience، Springer Science and Business Media LLC، 11 (1)، doi:10.1186/s13578-021-00643-z، ISSN 2045-3701.
- "Unique and conserved features of genome and proteome of SARS-coronavirus, an early split-off from the coronavirus group 2 lineage"، Journal of Molecular Biology، 331 (5): 991–1004، أغسطس 2003، doi:10.1016/S0022-2836(03)00865-9، PMC 7159028، PMID 12927536،
See Figure 1.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - Wang, Yuhang؛ Grunewald؛ Perlman (2020)، "Coronaviruses: An Updated Overview of Their Replication and Pathogenesis"، في Maier, Helena J. (المحرر)، Coronaviruses، New York, NY: Springer US، ج. 2203، ص. 1–29، doi:10.1007/978-1-0716-0900-2_1، ISBN 978-1-0716-0899-9، PMC 7682345، PMID 32833200، مؤرشف من الأصل في 2 مارس 2022.
{{استشهاد بكتاب}}: صيانة CS1: تنسيق PMC (link) - Hartenian, Ella؛ Nandakumar؛ Lari؛ Ly؛ Tucker؛ Glaunsinger (2020-09)، "The molecular virology of coronaviruses"، Journal of Biological Chemistry، 295 (37): 12910–12934، doi:10.1074/jbc.REV120.013930، PMC 7489918، PMID 32661197، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Madhugiri, R.؛ Fricke؛ Marz؛ Ziebuhr (2016)، "Coronavirus cis-Acting RNA Elements"، Advances in Virus Research، Elsevier، ج. 96، ص. 127–163، doi:10.1016/bs.aivir.2016.08.007، ISBN 978-0-12-804736-1، PMC 7112319، PMID 27712622، مؤرشف من الأصل في 20 مارس 2022.
{{استشهاد بكتاب}}: صيانة CS1: تنسيق PMC (link) - Kang, Hyojeung؛ Feng؛ Schroeder؛ Giedroc؛ Leibowitz (2006-11)، "Putative cis -Acting Stem-Loops in the 5′ Untranslated Region of the Severe Acute Respiratory Syndrome Coronavirus Can Substitute for Their Mouse Hepatitis Virus Counterparts"، Journal of Virology، 80 (21): 10600–10614، doi:10.1128/JVI.00455-06، ISSN 0022-538X، PMC 1641749، PMID 16920822، مؤرشف من الأصل في 13 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Chen, Shih-Cheng؛ Olsthoorn (2010-05)، "Group-specific structural features of the 5′-proximal sequences of coronavirus genomic RNAs"، Virology، 401 (1): 29–41، doi:10.1016/j.virol.2010.02.007، PMC 7111916، PMID 20202661، مؤرشف من الأصل في 3 مارس 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Li, Lichun؛ Kang؛ Liu؛ Makkinje؛ Williamson؛ Leibowitz؛ Giedroc (2008-03)، "Structural Lability in Stem–Loop 1 Drives a 5′ UTR–3′ UTR Interaction in Coronavirus Replication"، Journal of Molecular Biology، 377 (3): 790–803، doi:10.1016/j.jmb.2008.01.068، PMC 2652258، PMID 18289557، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Sola؛ Mateos-Gomez؛ Almazan؛ Zuñiga؛ Enjuanes (مارس 2011)، "RNA-RNA and RNA-protein interactions in coronavirus replication and transcription"، RNA Biology، 8 (2): 237–248، doi:10.4161/rna.8.2.14991، ISSN 1555-8584، PMC 3230552، PMID 21378501.
- Yang, D.؛ Liu؛ Giedroc؛ Leibowitz (01 سبتمبر 2011)، "Mouse Hepatitis Virus Stem-Loop 4 Functions as a Spacer Element Required To Drive Subgenomic RNA Synthesis"، Journal of Virology، 85 (17): 9199–9209، doi:10.1128/JVI.05092-11، ISSN 0022-538X، PMC 3165806، PMID 21715502، مؤرشف من الأصل في 6 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Zúñiga, Sonia؛ Sola؛ Alonso؛ Enjuanes (15 يناير 2004)، "Sequence Motifs Involved in the Regulation of Discontinuous Coronavirus Subgenomic RNA Synthesis"، Journal of Virology، 78 (2): 980–994، doi:10.1128/JVI.78.2.980-994.2004، ISSN 0022-538X، PMC 368802، PMID 14694129، مؤرشف من الأصل في 27 يناير 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Masters, Paul S. (2019-11)، "Coronavirus genomic RNA packaging"، Virology، 537: 198–207، doi:10.1016/j.virol.2019.08.031، PMC 7112113، PMID 31505321، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Madhugiri, Ramakanth؛ Fricke؛ Marz؛ Ziebuhr (2014-12)، "RNA structure analysis of alphacoronavirus terminal genome regions"، Virus Research، 194: 76–89، doi:10.1016/j.virusres.2014.10.001، PMC 7114417، PMID 25307890، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Dalton, Kevin؛ Casais؛ Shaw؛ Stirrups؛ Evans؛ Britton؛ Brown؛ Cavanagh (2001-01)، "cis -Acting Sequences Required for Coronavirus Infectious Bronchitis Virus Defective-RNA Replication and Packaging"، Journal of Virology، 75 (1): 125–133، doi:10.1128/JVI.75.1.125-133.2001، ISSN 0022-538X، PMC 113905، PMID 11119581، مؤرشف من الأصل في 12 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Goebel, Scott J.؛ Hsue؛ Dombrowski؛ Masters (15 يناير 2004)، "Characterization of the RNA Components of a Putative Molecular Switch in the 3′ Untranslated Region of the Murine Coronavirus Genome"، Journal of Virology، 78 (2): 669–682، doi:10.1128/JVI.78.2.669-682.2004، ISSN 0022-538X، PMC 368785، PMID 14694098، مؤرشف من الأصل في 8 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Züst, Roland؛ Miller؛ Goebel؛ Thiel؛ Masters (2008-02)، "Genetic Interactions between an Essential 3′ cis -Acting RNA Pseudoknot, Replicase Gene Products, and the Extreme 3′ End of the Mouse Coronavirus Genome"، Journal of Virology، 82 (3): 1214–1228، doi:10.1128/JVI.01690-07، ISSN 0022-538X، PMC 2224451، PMID 18032506، مؤرشف من الأصل في 12 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Liu, Qi؛ Johnson؛ Leibowitz (15 ديسمبر 2001)، "Secondary Structural Elements within the 3′ Untranslated Region of Mouse Hepatitis Virus Strain JHM Genomic RNA"، Journal of Virology، 75 (24): 12105–12113، doi:10.1128/JVI.75.24.12105-12113.2001، ISSN 0022-538X، PMC 116106، PMID 11711601، مؤرشف من الأصل في 12 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Goebel, Scott J.؛ Miller؛ Bennett؛ Bernard؛ Masters (2007-02)، "A Hypervariable Region within the 3′ cis -Acting Element of the Murine Coronavirus Genome Is Nonessential for RNA Synthesis but Affects Pathogenesis"، Journal of Virology، 81 (3): 1274–1287، doi:10.1128/JVI.00803-06، ISSN 0022-538X، PMC 1797510، PMID 17093194، مؤرشف من الأصل في 8 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Neuman, Benjamin W.؛ Kiss؛ Kunding؛ Bhella؛ Baksh؛ Connelly؛ Droese؛ Klaus؛ Makino (2011-04)، "A structural analysis of M protein in coronavirus assembly and morphology"، Journal of Structural Biology، 174 (1): 11–22، doi:10.1016/j.jsb.2010.11.021، PMC 4486061، PMID 21130884، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Schoeman, Dewald؛ Fielding (2019-12)، "Coronavirus envelope protein: current knowledge"، Virology Journal، 16 (1): 69، doi:10.1186/s12985-019-1182-0، ISSN 1743-422X، PMC 6537279، PMID 31133031، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Naskalska, Antonina؛ Dabrowska؛ Szczepanski؛ Milewska؛ Jasik؛ Pyrc (2019-10)، Gallagher, Tom (المحرر)، "Membrane Protein of Human Coronavirus NL63 Is Responsible for Interaction with the Adhesion Receptor"، Journal of Virology، 93 (19)، doi:10.1128/JVI.00355-19، ISSN 0022-538X، PMC 6744225، PMID 31315999، مؤرشف من الأصل في 9 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - DeDiego, Marta L.؛ Nieto-Torres؛ Jiménez-Guardeño؛ Regla-Nava؛ Álvarez؛ Oliveros؛ Zhao؛ Fett؛ Perlman (20 أكتوبر 2011)، Thiel, Volker (المحرر)، "Severe Acute Respiratory Syndrome Coronavirus Envelope Protein Regulates Cell Stress Response and Apoptosis"، PLoS Pathogens، 7 (10): e1002315، doi:10.1371/journal.ppat.1002315، ISSN 1553-7374، PMC 3197621، PMID 22028656، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - "A structural analysis of M protein in coronavirus assembly and morphology"، Journal of Structural Biology، 174 (1): 11–22، أبريل 2011، doi:10.1016/j.jsb.2010.11.021، PMC 4486061، PMID 21130884.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Membrane binding proteins of coronaviruses"، Future Virology، 14 (4): 275–286، أبريل 2019، doi:10.2217/fvl-2018-0144، PMC 7079996، PMID 32201500.
- Fung, To S.؛ Liu (17 يونيو 2014)، "Coronavirus infection, ER stress, apoptosis and innate immunity"، Frontiers in Microbiology، 5، doi:10.3389/fmicb.2014.00296، ISSN 1664-302X، PMC 4060729، PMID 24987391، مؤرشف من الأصل في 7 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Han, Dong P.؛ Lohani؛ Cho (2007-11)، "Specific Asparagine-Linked Glycosylation Sites Are Critical for DC-SIGN- and L-SIGN-Mediated Severe Acute Respiratory Syndrome Coronavirus Entry"، Journal of Virology، 81 (21): 12029–12039، doi:10.1128/JVI.00315-07، ISSN 0022-538X، PMC 2168787، PMID 17715238، مؤرشف من الأصل في 8 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Peng, G.؛ Sun؛ Rajashankar؛ Qian؛ Holmes؛ Li (28 يونيو 2011)، "Crystal structure of mouse coronavirus receptor-binding domain complexed with its murine receptor"، Proceedings of the National Academy of Sciences، 108 (26): 10696–10701، doi:10.1073/pnas.1104306108، ISSN 0027-8424، PMC 3127895، PMID 21670291، مؤرشف من الأصل في 22 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Li, Fang (29 سبتمبر 2016)، "Structure, Function, and Evolution of Coronavirus Spike Proteins"، Annual Review of Virology، 3 (1): 237–261، doi:10.1146/annurev-virology-110615-042301، ISSN 2327-056X، PMC 5457962، PMID 27578435، مؤرشف من الأصل في 3 مايو 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - de Haan, Cornelis A. M.؛ Stadler؛ Godeke؛ Bosch؛ Rottier (2004-06)، "Cleavage Inhibition of the Murine Coronavirus Spike Protein by a Furin-Like Enzyme Affects Cell-Cell but Not Virus-Cell Fusion"، Journal of Virology، 78 (11): 6048–6054، doi:10.1128/JVI.78.11.6048-6054.2004، ISSN 0022-538X، PMC 415802، PMID 15141003، مؤرشف من الأصل في 10 مارس 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Matsuyama, Shutoku؛ Shirato؛ Kawase؛ Terada؛ Kawachi؛ Fukushi؛ Kamitani (2018-10)، Gallagher, Tom (المحرر)، "Middle East Respiratory Syndrome Coronavirus Spike Protein Is Not Activated Directly by Cellular Furin during Viral Entry into Target Cells"، Journal of Virology، 92 (19)، doi:10.1128/JVI.00683-18، ISSN 0022-538X، PMC 6146822، PMID 30021905، مؤرشف من الأصل في 8 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Örd؛ Faustova؛ Loog (2020)، "The sequence at Spike S1/S2 site enables cleavage by furin and phospho-regulation in SARS-CoV2 but not in SARS-CoV1 or MERS-CoV"، Scientific Reports، 10 (1)، doi:10.1038/s41598-020-74101-0، ISSN 2045-2322.
- Shang, Jian؛ Wan؛ Luo؛ Ye؛ Geng؛ Auerbach؛ Li (26 مايو 2020)، "Cell entry mechanisms of SARS-CoV-2"، Proceedings of the National Academy of Sciences، 117 (21): 11727–11734، doi:10.1073/pnas.2003138117، ISSN 0027-8424، PMC 7260975، PMID 32376634، مؤرشف من الأصل في 27 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Park, Jung-Eun؛ Li؛ Barlan؛ Fehr؛ Perlman؛ McCray؛ Gallagher (25 أكتوبر 2016)، "Proteolytic processing of Middle East respiratory syndrome coronavirus spikes expands virus tropism"، Proceedings of the National Academy of Sciences، 113 (43): 12262–12267، doi:10.1073/pnas.1608147113، ISSN 0027-8424، PMC 5086990، PMID 27791014، مؤرشف من الأصل في 25 فبراير 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Chang, Chung-ke؛ Hou؛ Chang؛ Hsiao؛ Huang (2014-03)، "The SARS coronavirus nucleocapsid protein – Forms and functions"، Antiviral Research، 103: 39–50، doi:10.1016/j.antiviral.2013.12.009، PMC 7113676، PMID 24418573، مؤرشف من الأصل في 11 مايو 2022،
See Figure 4c.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - McBride, Ruth؛ van Zyl؛ Fielding (07 أغسطس 2014)، "The Coronavirus Nucleocapsid Is a Multifunctional Protein"، Viruses، 6 (8): 2991–3018، doi:10.3390/v6082991، ISSN 1999-4915، PMC 4147684، PMID 25105276، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Ye؛ West؛ Silletti؛ Corbett (سبتمبر 2020)، "Architecture and self‐assembly of the SARS‐CoV ‐2 nucleocapsid protein"، Protein Science، 29 (9): 1890–1901، doi:10.1002/pro.3909، PMC 7405475، PMID 32654247.
- Shah؛ Firmal؛ Alam؛ Ganguly؛ Chattopadhyay (07 أغسطس 2020)، "Overview of Immune Response During SARS-CoV-2 Infection: Lessons From the Past"، Frontiers in Immunology، 11: 1949، doi:10.3389/fimmu.2020.01949، PMC 7426442، PMID 32849654.
- Lu, Shan؛ Ye؛ Singh؛ وآخرون (21 يناير 2021)، "The SARS-CoV-2 nucleocapsid phosphoprotein forms mutually exclusive condensates with RNA and the membrane-associated M protein"، Nature Communications، Springer Science and Business Media LLC، 12 (1)، doi:10.1038/s41467-020-20768-y، ISSN 2041-1723.
- Li؛ Li (20 أبريل 2021)، "Immunologic Testing for SARS-CoV-2 Infection from the Antigen Perspective"، Journal of Clinical Microbiology، 59 (5): e02160-20، doi:10.1128/JCM.02160-20، PMC 8091849، PMID 33318065.
- Zúñiga؛ Cruz؛ Sola؛ Mateos-Gómez؛ Palacio؛ Enjuanes (15 فبراير 2010)، "Coronavirus Nucleocapsid Protein Facilitates Template Switching and Is Required for Efficient Transcription"، Journal of Virology، 84 (4): 2169–2175، doi:10.1128/JVI.02011-09، PMC 2812394، PMID 19955314.
- Su؛ Chen؛ Qi؛ Shi؛ Feng؛ Sun (05 نوفمبر 2020)، "A Mini-Review on Cell Cycle Regulation of Coronavirus Infection"، Frontiers in Veterinary Science، 7: 586826، doi:10.3389/fvets.2020.586826، PMC 7674852، PMID 33251267.
- Zeng, Qinghong؛ Langereis؛ van Vliet؛ Huizinga؛ de Groot (01 يوليو 2008)، "Structure of coronavirus hemagglutinin-esterase offers insight into corona and influenza virus evolution"، Proceedings of the National Academy of Sciences، 105 (26): 9065–9069، doi:10.1073/pnas.0800502105، ISSN 0027-8424، PMC 2449365، PMID 18550812، مؤرشف من الأصل في 29 نوفمبر 2021.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Zeng, Qinghong؛ Langereis؛ van Vliet؛ Huizinga؛ de Groot (01 يوليو 2008)، "Structure of coronavirus hemagglutinin-esterase offers insight into corona and influenza virus evolution"، Proceedings of the National Academy of Sciences، 105 (26): 9065–9069، doi:10.1073/pnas.0800502105، ISSN 0027-8424، PMC 2449365، PMID 18550812، مؤرشف من الأصل في 29 نوفمبر 2021.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Bakkers, Mark J.G.؛ Lang؛ Feitsma؛ Hulswit؛ de Poot؛ van Vliet؛ Margine؛ de Groot-Mijnes؛ van Kuppeveld (2017-03)، "Betacoronavirus Adaptation to Humans Involved Progressive Loss of Hemagglutinin-Esterase Lectin Activity"، Cell Host & Microbe، 21 (3): 356–366، doi:10.1016/j.chom.2017.02.008، PMC 7104930، PMID 28279346، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Smith؛ Denison (05 ديسمبر 2013)، "Coronaviruses as DNA Wannabes: A New Model for the Regulation of RNA Virus Replication Fidelity"، PLoS Pathogens، 9 (12): e1003760، doi:10.1371/journal.ppat.1003760.
- Gulyaeva؛ Gorbalenya (يناير 2021)، "A nidovirus perspective on SARS-CoV-2"، Biochemical and Biophysical Research Communications، 538: 24–34، doi:10.1016/j.bbrc.2020.11.015.
- Hartenian؛ Nandakumar؛ Lari؛ Ly؛ Tucker؛ Glaunsinger (سبتمبر 2020)، "The molecular virology of coronaviruses"، Journal of Biological Chemistry، 295 (37): 12910–12934، doi:10.1074/jbc.REV120.013930.
- V’kovski؛ Kratzel؛ Steiner؛ Stalder؛ Thiel (مارس 2021)، "Coronavirus biology and replication: implications for SARS-CoV-2"، Nature Reviews Microbiology، 19 (3): 155–170، doi:10.1038/s41579-020-00468-6.
- Posthuma؛ te Velthuis؛ Snijder (أبريل 2017)، "Nidovirus RNA polymerases: Complex enzymes handling exceptional RNA genomes"، Virus Research، 234: 58–73، doi:10.1016/j.virusres.2017.01.023.
- Irigoyen؛ Firth؛ Jones؛ Chung؛ Siddell؛ Brierley (26 فبراير 2016)، "High-Resolution Analysis of Coronavirus Gene Expression by RNA Sequencing and Ribosome Profiling"، PLOS Pathogens، 12 (2): e1005473، doi:10.1371/journal.ppat.1005473.
- Finkel؛ Mizrahi؛ Nachshon؛ Weingarten-Gabbay؛ Morgenstern؛ Yahalom-Ronen؛ Tamir؛ Achdout؛ Stein؛ Israeli, Ofir؛ Beth-Din, Adi؛ Melamed, Sharon؛ Weiss, Shay؛ Israely, Tomer؛ Paran, Nir؛ Schwartz, Michal؛ Stern-Ginossar, Noam (07 يناير 2021)، "The coding capacity of SARS-CoV-2"، Nature، 589 (7840): 125–130، doi:10.1038/s41586-020-2739-1.
- Rohaim؛ El Naggar؛ Clayton؛ Munir (يناير 2021)، "Structural and functional insights into non-structural proteins of coronaviruses"، Microbial Pathogenesis، 150: 104641، doi:10.1016/j.micpath.2020.104641.
- Chen؛ Liu؛ Guo (أبريل 2020)، "Emerging coronaviruses: Genome structure, replication, and pathogenesis"، Journal of Medical Virology، 92 (4): 418–423، doi:10.1002/jmv.25681.
- Michel, Christian Jean؛ Mayer؛ Poch؛ Thompson (2020-12)، "Characterization of accessory genes in coronavirus genomes"، Virology Journal، 17 (1): 131، doi:10.1186/s12985-020-01402-1، ISSN 1743-422X، PMC 7450977، PMID 32854725، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Fang, Puxian؛ Fang؛ Zhang؛ Xia؛ Xiao (13 يونيو 2021)، "Functions of Coronavirus Accessory Proteins: Overview of the State of the Art"، Viruses، MDPI AG، 13 (6): 1139، doi:10.3390/v13061139، ISSN 1999-4915.
- Kopecky-Bromberg, Sarah A.؛ Martínez-Sobrido؛ Frieman؛ Baric؛ Palese (15 يناير 2007)، "Severe Acute Respiratory Syndrome Coronavirus Open Reading Frame (ORF) 3b, ORF 6, and Nucleocapsid Proteins Function as Interferon Antagonists"، Journal of Virology، American Society for Microbiology، 81 (2): 548–557، doi:10.1128/jvi.01782-06، ISSN 0022-538X.
- Wu, Jing؛ Shi؛ Pan؛ Wu؛ Hou؛ Zhang؛ Zhong؛ Tang؛ Du؛ Wang, Luying؛ Wo, Jing؛ Mu, Jingfang؛ Qiu, Yang؛ Yang, Ke؛ Zhang, Lei-Ke؛ Ye, Bang-Ce؛ Qi, Nan (2021)، "SARS-CoV-2 ORF9b inhibits RIG-I-MAVS antiviral signaling by interrupting K63-linked ubiquitination of NEMO"، Cell Reports، Elsevier BV، 34 (7): 108761، doi:10.1016/j.celrep.2021.108761، ISSN 2211-1247.
- Frieman, Matthew؛ Yount؛ Heise؛ Kopecky-Bromberg؛ Palese؛ Baric (15 سبتمبر 2007)، "Severe Acute Respiratory Syndrome Coronavirus ORF6 Antagonizes STAT1 Function by Sequestering Nuclear Import Factors on the Rough Endoplasmic Reticulum/Golgi Membrane"، Journal of Virology، American Society for Microbiology، 81 (18): 9812–9824، doi:10.1128/jvi.01012-07، ISSN 0022-538X.
- Rabouw, Huib H.؛ Langereis؛ Knaap؛ Dalebout؛ Canton؛ Sola؛ Enjuanes؛ Bredenbeek؛ Kikkert؛ de Groot, Raoul J.؛ van Kuppeveld, Frank J. M. (26 أكتوبر 2016)، Frieman, Matthew B. (المحرر)، "Middle East Respiratory Coronavirus Accessory Protein 4a Inhibits PKR-Mediated Antiviral Stress Responses"، PLOS Pathogens، Public Library of Science (PLoS)، 12 (10): e1005982، doi:10.1371/journal.ppat.1005982، ISSN 1553-7374.
- Simmons, Graham؛ Zmora؛ Gierer؛ Heurich؛ Pöhlmann (2013-12)، "Proteolytic activation of the SARS-coronavirus spike protein: Cutting enzymes at the cutting edge of antiviral research"، Antiviral Research، 100 (3): 605–614، doi:10.1016/j.antiviral.2013.09.028، PMC 3889862، PMID 24121034، مؤرشف من الأصل في 11 مايو 2022،
See Figure 2.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Yamada, Yoshiyuki؛ Liu (2009-09)، "Proteolytic Activation of the Spike Protein at a Novel RRRR/S Motif Is Implicated in Furin-Dependent Entry, Syncytium Formation, and Infectivity of Coronavirus Infectious Bronchitis Virus in Cultured Cells"، Journal of Virology، 83 (17): 8744–8758، doi:10.1128/JVI.00613-09، ISSN 0022-538X، PMC 2738192، PMID 19553314، مؤرشف من الأصل في 9 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Wong, Hui Hui؛ Kumar؛ Tay؛ Moreau؛ Liu؛ Bard (2015-11)، Perlman, S. (المحرر)، "Genome-Wide Screen Reveals Valosin-Containing Protein Requirement for Coronavirus Exit from Endosomes"، Journal of Virology، 89 (21): 11116–11128، doi:10.1128/JVI.01360-15، ISSN 0022-538X، PMC 4621105، PMID 26311884، مؤرشف من الأصل في 7 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Baranov, Pavel V.؛ Henderson؛ Anderson؛ Gesteland؛ Atkins؛ Howard (2005)، "Programmed ribosomal frameshifting in decoding the SARS-CoV genome"، Virology، Elsevier BV، 332 (2): 498–510، doi:10.1016/j.virol.2004.11.038، ISSN 0042-6822.
- Brierley, Ian؛ Digard؛ Inglis (1989)، "Characterization of an efficient coronavirus ribosomal frameshifting signal: Requirement for an RNA pseudoknot"، Cell، Elsevier BV، 57 (4): 537–547، doi:10.1016/0092-8674(89)90124-4، ISSN 0092-8674.
- Hagemeijer, Marne C.؛ Monastyrska؛ Griffith؛ van der Sluijs؛ Voortman؛ van Bergen en Henegouwen؛ Vonk؛ Rottier؛ Reggiori (2014-06)، "Membrane rearrangements mediated by coronavirus nonstructural proteins 3 and 4"، Virology، 458–459: 125–135، doi:10.1016/j.virol.2014.04.027، PMC 7111329، PMID 24928045، مؤرشف من الأصل في 13 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Noack, J.؛ Bernasconi؛ Molinari (15 سبتمبر 2014)، "How Viruses Hijack the ERAD Tuning Machinery"، Journal of Virology، 88 (18): 10272–10275، doi:10.1128/JVI.00801-14، ISSN 0022-538X، PMC 4178841، PMID 24990995، مؤرشف من الأصل في 14 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Miller, Katelyn؛ McGrath؛ Hu؛ Ariannejad؛ Weston؛ Frieman؛ Jackson (01 ديسمبر 2020)، "Coronavirus interactions with the cellular autophagy machinery"، Autophagy، 16 (12): 2131–2139، doi:10.1080/15548627.2020.1817280، ISSN 1554-8627، PMC 7755319، PMID 32964796، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Knoops, Kèvin؛ Kikkert؛ Worm؛ Zevenhoven-Dobbe؛ van der Meer؛ Koster؛ Mommaas؛ Snijder (16 سبتمبر 2008)، Emerman, Michael (المحرر)، "SARS-Coronavirus Replication Is Supported by a Reticulovesicular Network of Modified Endoplasmic Reticulum"، PLoS Biology، 6 (9): e226، doi:10.1371/journal.pbio.0060226، ISSN 1545-7885، PMC 2535663، PMID 18798692، مؤرشف من الأصل في 17 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Sexton, Nicole R.؛ Smith؛ Blanc؛ Vignuzzi؛ Peersen؛ Denison (15 أغسطس 2016)، Perlman, S. (المحرر)، "Homology-Based Identification of a Mutation in the Coronavirus RNA-Dependent RNA Polymerase That Confers Resistance to Multiple Mutagens"، Journal of Virology، 90 (16): 7415–7428، doi:10.1128/JVI.00080-16، ISSN 0022-538X، PMC 4984655، PMID 27279608، مؤرشف من الأصل في 29 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Wertheim, Joel O.؛ Chu؛ Peiris؛ Kosakovsky Pond؛ Poon (15 يونيو 2013)، "A Case for the Ancient Origin of Coronaviruses"، Journal of Virology، 87 (12): 7039–7045، doi:10.1128/JVI.03273-12، ISSN 0022-538X، PMC 3676139، PMID 23596293، مؤرشف من الأصل في 9 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Eckerle, Lance D.؛ Becker؛ Halpin؛ Li؛ Venter؛ Lu؛ Scherbakova؛ Graham؛ Baric (06 مايو 2010)، Emerman, Michael (المحرر)، "Infidelity of SARS-CoV Nsp14-Exonuclease Mutant Virus Replication Is Revealed by Complete Genome Sequencing"، PLoS Pathogens، 6 (5): e1000896، doi:10.1371/journal.ppat.1000896، ISSN 1553-7374، PMC 2865531، PMID 20463816، مؤرشف من الأصل في 29 مارس 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Smith, Everett Clinton؛ Denison (05 ديسمبر 2013)، Racaniello, Vincent (المحرر)، "Coronaviruses as DNA Wannabes: A New Model for the Regulation of RNA Virus Replication Fidelity"، PLoS Pathogens، 9 (12): e1003760، doi:10.1371/journal.ppat.1003760، ISSN 1553-7374، PMC 3857799، PMID 24348241، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - V'kovski, Philip؛ Gerber؛ Kelly؛ Pfaender؛ Ebert؛ Braga Lagache؛ Simillion؛ Portmann؛ Stalder (11 يناير 2019)، "Determination of host proteins composing the microenvironment of coronavirus replicase complexes by proximity-labeling"، eLife، 8: e42037، doi:10.7554/eLife.42037، ISSN 2050-084X، PMC 6372286، PMID 30632963، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Peng, Qi؛ Peng؛ Yuan؛ Zhao؛ Wang؛ Wang؛ Wang؛ Sun؛ Fan؛ Qi, Jianxun؛ Gao, George F.؛ Shi, Yi (2020)، "Structural and Biochemical Characterization of the nsp12-nsp7-nsp8 Core Polymerase Complex from SARS-CoV-2"، Cell Reports، Elsevier BV، 31 (11): 107774، doi:10.1016/j.celrep.2020.107774، ISSN 2211-1247.
- Yan, Liming؛ Zhang؛ Ge؛ Zheng؛ Gao؛ Wang؛ Jia؛ Wang؛ Huang؛ Li, Mingyu؛ Wang, Quan؛ Rao, Zihe؛ Lou, Zhiyong (18 نوفمبر 2020)، "Architecture of a SARS-CoV-2 mini replication and transcription complex"، Nature Communications، Springer Science and Business Media LLC، 11 (1)، doi:10.1038/s41467-020-19770-1، ISSN 2041-1723.
- Hillen, Hauke S.؛ Kokic؛ Farnung؛ Dienemann؛ Tegunov؛ Cramer (21 مايو 2020)، "Structure of replicating SARS-CoV-2 polymerase"، Nature، Springer Science and Business Media LLC، 584 (7819): 154–156، doi:10.1038/s41586-020-2368-8، ISSN 0028-0836.
- Gao, Yan؛ Yan؛ Huang؛ Liu؛ Zhao؛ Cao؛ Wang؛ Sun؛ Ming؛ Zhang, Lianqi؛ Ge, Ji؛ Zheng, Litao؛ Zhang, Ying؛ Wang, Haofeng؛ Zhu, Yan؛ Zhu, Chen؛ Hu, Tianyu؛ Hua, Tian؛ Zhang, Bing؛ Yang, Xiuna؛ Li, Jun؛ Yang, Haitao؛ Liu, Zhijie؛ Xu, Wenqing؛ Guddat, Luke W.؛ Wang, Quan؛ Lou, Zhiyong؛ Rao, Zihe (15 مايو 2020)، "Structure of the RNA-dependent RNA polymerase from COVID-19 virus"، Science، American Association for the Advancement of Science (AAAS)، 368 (6492): 779–782، doi:10.1126/science.abb7498، ISSN 0036-8075.
- Chen, James؛ Malone؛ Llewellyn؛ Grasso؛ Shelton؛ Olinares؛ Maruthi؛ Eng؛ Vatandaslar؛ Chait, Brian T.؛ Kapoor, Tarun M.؛ Darst, Seth A.؛ Campbell, Elizabeth A. (2020)، "Structural Basis for Helicase-Polymerase Coupling in the SARS-CoV-2 Replication-Transcription Complex"، Cell، Elsevier BV، 182 (6): 1560–1573.e13، doi:10.1016/j.cell.2020.07.033، ISSN 0092-8674.
- Jochheim, Florian A.؛ Tegunov؛ Hillen؛ Schmitzová؛ Kokic؛ Dienemann؛ Cramer (24 أغسطس 2021)، "The structure of a dimeric form of SARS-CoV-2 polymerase"، Communications Biology، Springer Science and Business Media LLC، 4 (1)، doi:10.1038/s42003-021-02529-9، ISSN 2399-3642.
- Antoine A.F. de Vries, Marian C. Horzinek, Peter J. M. Rottier, Raoul J. de Groot (1997)، "The Genome Organization of the Nidovirales: Similarities and Differences between Arteri-, Toro-, and Coronaviruses"، Seminars in Virology، 8 (1): 33–47، CiteSeerX 10.1.1.462.1825، doi:10.1006/smvy.1997.0104، PMC 7128191، PMID 32288441، S2CID 85383257.
- Sola, Isabel؛ Almazán؛ Zúñiga؛ Enjuanes (09 نوفمبر 2015)، "Continuous and Discontinuous RNA Synthesis in Coronaviruses"، Annual Review of Virology، Annual Reviews، 2 (1): 265–288، doi:10.1146/annurev-virology-100114-055218، ISSN 2327-056X.
- Wang, Dehe؛ Jiang؛ Feng؛ وآخرون (2021)، "The SARS-CoV-2 subgenome landscape and its novel regulatory features"، Molecular Cell، Elsevier BV، 81 (10): 2135–2147.e5، doi:10.1016/j.molcel.2021.02.036، ISSN 1097-2765.
- Ziv, Omer؛ Price؛ Shalamova؛ Kamenova؛ Goodfellow؛ Weber؛ Miska (2020)، "The Short- and Long-Range RNA-RNA Interactome of SARS-CoV-2"، Molecular Cell، Elsevier BV، 80 (6): 1067–1077.e5، doi:10.1016/j.molcel.2020.11.004، ISSN 1097-2765.
- Enjuanes, Luis؛ Almazán؛ Sola؛ Zuñiga (01 أكتوبر 2006)، "Biochemical Aspects of Coronavirus Replication and Virus-Host Interaction"، Annual Review of Microbiology، Annual Reviews، 60 (1): 211–230، doi:10.1146/annurev.micro.60.080805.142157، ISSN 0066-4227.
- Zúñiga, Sonia؛ Cruz؛ Sola؛ Mateos-Gómez؛ Palacio؛ Enjuanes (15 فبراير 2010)، "Coronavirus Nucleocapsid Protein Facilitates Template Switching and Is Required for Efficient Transcription"، Journal of Virology، American Society for Microbiology، 84 (4): 2169–2175، doi:10.1128/jvi.02011-09، ISSN 0022-538X.
- Wu, Chia-Hsin؛ Chen؛ Yeh (2014)، "Nucleocapsid Phosphorylation and RNA Helicase DDX1 Recruitment Enables Coronavirus Transition from Discontinuous to Continuous Transcription"، Cell Host & Microbe، Elsevier BV، 16 (4): 462–472، doi:10.1016/j.chom.2014.09.009، ISSN 1931-3128.
- Kim, Dongwan؛ Lee؛ Yang؛ Kim؛ Kim؛ Chang (2020)، "The Architecture of SARS-CoV-2 Transcriptome"، Cell، Elsevier BV، 181 (4): 914–921.e10، doi:10.1016/j.cell.2020.04.011، ISSN 0092-8674.
- Payne, Susan (2017)، "Family Coronaviridae"، Viruses، Elsevier، ص. 149–158، doi:10.1016/b978-0-12-803109-4.00017-9، ISBN 978-0-12-803109-4، PMC 7149805، مؤرشف من الأصل في 19 أبريل 2022.
{{استشهاد بكتاب}}: صيانة CS1: تنسيق PMC (link) - Su, Shuo؛ Wong؛ Shi؛ Liu؛ Lai؛ Zhou؛ Liu؛ Bi؛ Gao (2016-06)، "Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses"، Trends in Microbiology، 24 (6): 490–502، doi:10.1016/j.tim.2016.03.003، PMC 7125511، PMID 27012512، مؤرشف من الأصل في 27 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Zhang, Yue؛ Li؛ Xiao؛ Zhang؛ Wang؛ Chen؛ Paranhos-Baccalà؛ Ren؛ Wang (2015-06)، "Genotype shift in human coronavirus OC43 and emergence of a novel genotype by natural recombination"، Journal of Infection، 70 (6): 641–650، doi:10.1016/j.jinf.2014.12.005، PMC 7112537، PMID 25530469، مؤرشف من الأصل في 20 مارس 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Pyrc, Krzysztof؛ Dijkman؛ Deng؛ Jebbink؛ Ross؛ Berkhout؛ van der Hoek (2006-12)، "Mosaic Structure of Human Coronavirus NL63, One Thousand Years of Evolution"، Journal of Molecular Biology، 364 (5): 964–973، doi:10.1016/j.jmb.2006.09.074، PMC 7094706، PMID 17054987، مؤرشف من الأصل في 14 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Zhang, X. W.؛ Yap؛ Danchin (2005-01)، "Testing the hypothesis of a recombinant origin of the SARS-associated coronavirus"، Archives of Virology، 150 (1): 1–20، doi:10.1007/s00705-004-0413-9، ISSN 0304-8608، PMC 7087341، PMID 15480857، مؤرشف من الأصل في 2 مارس 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Su, Shuo؛ Wong؛ Shi؛ Liu؛ Lai؛ Zhou؛ Liu؛ Bi؛ Gao (2016-06)، "Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses"، Trends in Microbiology، 24 (6): 490–502، doi:10.1016/j.tim.2016.03.003، PMC 7125511، PMID 27012512، مؤرشف من الأصل في 27 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Mendonça, Luiza؛ Howe؛ Gilchrist؛ وآخرون (30 يوليو 2021)، "Correlative multi-scale cryo-imaging unveils SARS-CoV-2 assembly and egress"، Nature Communications، Springer Science and Business Media LLC، 12 (1)، doi:10.1038/s41467-021-24887-y، ISSN 2041-1723.
- Ruch, Travis R.؛ Machamer (08 مارس 2012)، "The Coronavirus E Protein: Assembly and Beyond"، Viruses، MDPI AG، 4 (3): 363–382، doi:10.3390/v4030363، ISSN 1999-4915.
- J Alsaadi, Entedar A؛ Jones (2019)، "Membrane binding proteins of coronaviruses"، Future Virology، Future Medicine Ltd، 14 (4): 275–286، doi:10.2217/fvl-2018-0144، ISSN 1746-0794.
- Fehr, Anthony R.؛ Perlman (2015)، "Coronaviruses: An Overview of Their Replication and Pathogenesis"، في Maier, Helena Jane (المحرر)، Coronaviruses، New York, NY: Springer New York، ج. 1282، ص. 1–23، doi:10.1007/978-1-4939-2438-7_1، ISBN 978-1-4939-2437-0، PMC 4369385، PMID 25720466، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بكتاب}}: صيانة CS1: تنسيق PMC (link) - Cubuk, Jasmine؛ Alston؛ Incicco؛ وآخرون (29 مارس 2021)، "The SARS-CoV-2 nucleocapsid protein is dynamic, disordered, and phase separates with RNA"، Nature Communications، Springer Science and Business Media LLC، 12 (1)، doi:10.1038/s41467-021-21953-3، ISSN 2041-1723.
- Ye, Qiaozhen؛ West؛ Silletti؛ Corbett (06 أغسطس 2020)، "Architecture and self‐assembly of the SARS‐CoV ‐2 nucleocapsid protein"، Protein Science، Wiley، 29 (9): 1890–1901، doi:10.1002/pro.3909، ISSN 0961-8368.
- Carlson, Christopher R.؛ Asfaha؛ Ghent؛ وآخرون (2020)، "Phosphoregulation of Phase Separation by the SARS-CoV-2 N Protein Suggests a Biophysical Basis for its Dual Functions"، Molecular Cell، Elsevier BV، 80 (6): 1092–1103.e4، doi:10.1016/j.molcel.2020.11.025، ISSN 1097-2765.
- Kuo, Lili؛ Hurst-Hess؛ Koetzner؛ Masters (2016)، Perlman, S. (المحرر)، "Analyses of Coronavirus Assembly Interactions with Interspecies Membrane and Nucleocapsid Protein Chimeras"، Journal of Virology، American Society for Microbiology، 90 (9): 4357–4368، doi:10.1128/jvi.03212-15، ISSN 0022-538X.
- Yao, Hangping؛ Song؛ Chen؛ وآخرون (2020)، "Molecular Architecture of the SARS-CoV-2 Virus"، Cell، Elsevier BV، 183 (3): 730–738.e13، doi:10.1016/j.cell.2020.09.018، ISSN 0092-8674.
- Cui, Jie؛ Li؛ Shi (2019-03)، "Origin and evolution of pathogenic coronaviruses"، Nature Reviews Microbiology، 17 (3): 181–192، doi:10.1038/s41579-018-0118-9، ISSN 1740-1526، PMC 7097006، PMID 30531947، مؤرشف من الأصل في 11 مايو 2022،
Different SARS-CoV strains isolated from several hosts vary in their binding affinities for human ACE2 and consequently in their infectivity of human cells 76, 78 (Fig. 6b)
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Decaro, Nicola (2011)، "Alphacoronavirus‡: Coronaviridae"، في Tidona, Christian (المحرر)، The Springer Index of Viruses، New York, NY: Springer New York، ص. 371–383، doi:10.1007/978-0-387-95919-1_56، ISBN 978-0-387-95918-4، PMC 7176201، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بكتاب}}: صيانة CS1: تنسيق PMC (link) - Li, Fang؛ Li؛ Farzan؛ Harrison (16 سبتمبر 2005)، "Structure of SARS Coronavirus Spike Receptor-Binding Domain Complexed with Receptor"، Science، 309 (5742): 1864–1868، Bibcode:2005Sci...309.1864L، doi:10.1126/science.1116480، ISSN 0036-8075، PMID 16166518، مؤرشف من الأصل في 8 أبريل 2022.
- Wolff, Georg؛ Melia؛ Snijder؛ Bárcena (2020)، "Double-Membrane Vesicles as Platforms for Viral Replication"، Trends in Microbiology، Elsevier BV، 28 (12): 1022–1033، doi:10.1016/j.tim.2020.05.009، ISSN 0966-842X.
- Liao, Y.؛ Fung؛ Huang؛ Fang؛ Zhong؛ Liu (15 يوليو 2013)، "Upregulation of CHOP/GADD153 during Coronavirus Infectious Bronchitis Virus Infection Modulates Apoptosis by Restricting Activation of the Extracellular Signal-Regulated Kinase Pathway"، Journal of Virology، 87 (14): 8124–8134، doi:10.1128/JVI.00626-13، ISSN 0022-538X، PMC 3700216، PMID 23678184، مؤرشف من الأصل في 9 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Ma, Yanlong؛ Wang؛ Xue؛ Fu؛ Zhang؛ Li؛ Yin؛ Xu؛ Feng (15 نوفمبر 2018)، Williams, Bryan R. G. (المحرر)، "The Coronavirus Transmissible Gastroenteritis Virus Evades the Type I Interferon Response through IRE1α-Mediated Manipulation of the MicroRNA miR-30a-5p/SOCS1/3 Axis"، Journal of Virology، 92 (22)، doi:10.1128/JVI.00728-18، ISSN 0022-538X، PMC 6206482، PMID 30185587، مؤرشف من الأصل في 13 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Bechill, John؛ Chen؛ Brewer؛ Baker (2008-05)، "Coronavirus Infection Modulates the Unfolded Protein Response and Mediates Sustained Translational Repression"، Journal of Virology، 82 (9): 4492–4501، doi:10.1128/JVI.00017-08، ISSN 0022-538X، PMC 2293058، PMID 18305036، مؤرشف من الأصل في 8 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Sung, Shu-Chiun؛ Chao؛ Jeng؛ Yang؛ Lai (2009-05)، "The 8ab protein of SARS-CoV is a luminal ER membrane-associated protein and induces the activation of ATF6"، Virology، 387 (2): 402–413، doi:10.1016/j.virol.2009.02.021، PMC 7103415، PMID 19304306، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Fung, To Sing؛ Huang؛ Liu (2014-12)، "Coronavirus-induced ER stress response and its involvement in regulation of coronavirus–host interactions"، Virus Research، 194: 110–123، doi:10.1016/j.virusres.2014.09.016، PMC 7114476، PMID 25304691، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Favreau, D. J.؛ Meessen-Pinard؛ Desforges؛ Talbot (01 يناير 2012)، "Human Coronavirus-Induced Neuronal Programmed Cell Death Is Cyclophilin D Dependent and Potentially Caspase Dispensable"، Journal of Virology، 86 (1): 81–93، doi:10.1128/JVI.06062-11، ISSN 0022-538X، PMC 3255912، PMID 22013052، مؤرشف من الأصل في 11 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Ren, Yujie؛ Shu؛ Wu؛ Mu؛ Wang؛ Huang؛ Han؛ Zhang؛ Zhou (2020-08)، "The ORF3a protein of SARS-CoV-2 induces apoptosis in cells"، Cellular & Molecular Immunology، 17 (8): 881–883، doi:10.1038/s41423-020-0485-9، ISSN 1672-7681، PMC 7301057، PMID 32555321، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Zhu, Na؛ Wang؛ Liu؛ Liang؛ Wang؛ Ye؛ Huang؛ Zhao؛ Wang (2020-12)، "Morphogenesis and cytopathic effect of SARS-CoV-2 infection in human airway epithelial cells"، Nature Communications، 11 (1): 3910، doi:10.1038/s41467-020-17796-z، ISSN 2041-1723، PMC 7413383، PMID 32764693، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Mesel-Lemoine, M.؛ Millet؛ Vidalain؛ Law؛ Vabret؛ Lorin؛ Escriou؛ Albert؛ Nal (15 يوليو 2012)، "A Human Coronavirus Responsible for the Common Cold Massively Kills Dendritic Cells but Not Monocytes"، Journal of Virology، 86 (14): 7577–7587، doi:10.1128/JVI.00269-12، ISSN 0022-538X، PMC 3416289، PMID 22553325، مؤرشف من الأصل في 17 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Fung, To؛ Liao؛ Liu (04 يوليو 2016)، "Regulation of Stress Responses and Translational Control by Coronavirus"، Viruses، 8 (7): 184، doi:10.3390/v8070184، ISSN 1999-4915، PMC 4974519، PMID 27384577، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Chang, Ya-Jen؛ Liu؛ Chiang؛ Chao؛ Chen (15 ديسمبر 2004)، "Induction of IL-8 Release in Lung Cells via Activator Protein-1 by Recombinant Baculovirus Displaying Severe Acute Respiratory Syndrome-Coronavirus Spike Proteins: Identification of Two Functional Regions"، The Journal of Immunology، 173 (12): 7602–7614، doi:10.4049/jimmunol.173.12.7602، ISSN 0022-1767، مؤرشف من الأصل في 11 مايو 2022.
- Edridge, Arthur W. D.؛ Kaczorowska؛ Hoste؛ Bakker؛ Klein؛ Loens؛ Jebbink؛ Matser؛ Kinsella (2020-11)، "Seasonal coronavirus protective immunity is short-lasting"، Nature Medicine، 26 (11): 1691–1693، doi:10.1038/s41591-020-1083-1، ISSN 1078-8956، مؤرشف من الأصل في 10 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة) - Temperton, Nigel J.؛ Chan؛ Simmons؛ Zambon؛ Tedder؛ Takeuchi؛ Weiss (2005-03)، "Longitudinally Profiling Neutralizing Antibody Response to SARS Coronavirus with Pseudotypes"، Emerging Infectious Diseases، 11 (3): 411–416، doi:10.3201/eid1103.040906، ISSN 1080-6040، PMC 3298259، PMID 15757556، مؤرشف من الأصل في 12 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Zou, Lirong؛ Ruan؛ Huang؛ Liang؛ Huang؛ Hong؛ Yu؛ Kang؛ Song (19 مارس 2020)، "SARS-CoV-2 Viral Load in Upper Respiratory Specimens of Infected Patients"، New England Journal of Medicine، 382 (12): 1177–1179، doi:10.1056/NEJMc2001737، ISSN 0028-4793، PMC 7121626، PMID 32074444، مؤرشف من الأصل في 6 مايو 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Tang, Fang؛ Quan؛ Xin؛ Wrammert؛ Ma؛ Lv؛ Wang؛ Yang؛ Richardus (15 يونيو 2011)، "Lack of Peripheral Memory B Cell Responses in Recovered Patients with Severe Acute Respiratory Syndrome: A Six-Year Follow-Up Study"، The Journal of Immunology، 186 (12): 7264–7268، doi:10.4049/jimmunol.0903490، ISSN 0022-1767، مؤرشف من الأصل في 20 أبريل 2022.
- Sariol, Alan؛ Perlman (2020-08)، "Lessons for COVID-19 Immunity from Other Coronavirus Infections"، Immunity، 53 (2): 248–263، doi:10.1016/j.immuni.2020.07.005، PMC 7359787، PMID 32717182، مؤرشف من الأصل في 6 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Cavanagh, Dave (2007-03)، "Coronavirus avian infectious bronchitis virus"، Veterinary Research، 38 (2): 281–297، doi:10.1051/vetres:2006055، ISSN 0928-4249، مؤرشف من الأصل في 3 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة) - Wathelet, Marc G.؛ Orr؛ Frieman؛ Baric (2007-11)، "Severe Acute Respiratory Syndrome Coronavirus Evades Antiviral Signaling: Role of nsp1 and Rational Design of an Attenuated Strain"، Journal of Virology، 81 (21): 11620–11633، doi:10.1128/JVI.00702-07، ISSN 0022-538X، PMC 2168762، PMID 17715225، مؤرشف من الأصل في 9 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Fung, To Sing؛ Liu (01 يونيو 2018)، "Post-translational modifications of coronavirus proteins: roles and function"، Future Virology، 13 (6): 405–430، doi:10.2217/fvl-2018-0008، ISSN 1746-0794، PMC 7080180، PMID 32201497، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Devaraj, Santhana G.؛ Wang؛ Chen؛ Chen؛ Tseng؛ Barretto؛ Lin؛ Peters؛ Tseng (2007-11)، "Regulation of IRF-3-dependent Innate Immunity by the Papain-like Protease Domain of the Severe Acute Respiratory Syndrome Coronavirus"، Journal of Biological Chemistry، 282 (44): 32208–32221، doi:10.1074/jbc.M704870200، PMC 2756044، PMID 17761676، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Kamitani, W.؛ Narayanan؛ Huang؛ Lokugamage؛ Ikegami؛ Ito؛ Kubo؛ Makino (22 أغسطس 2006)، "Severe acute respiratory syndrome coronavirus nsp1 protein suppresses host gene expression by promoting host mRNA degradation"، Proceedings of the National Academy of Sciences، 103 (34): 12885–12890، doi:10.1073/pnas.0603144103، ISSN 0027-8424، PMC 1568942، PMID 16912115، مؤرشف من الأصل في 4 مارس 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Perlman, Stanley؛ Hogue؛ Li؛ Yount؛ Baric؛ Weiss (30 أبريل 2019)، Palese, Peter (المحرر)، "Antagonism of dsRNA-Induced Innate Immune Pathways by NS4a and NS4b Accessory Proteins during MERS Coronavirus Infection"، mBio، 10 (2)، doi:10.1128/mBio.00319-19، ISSN 2161-2129، PMC 6437052، PMID 30914508، مؤرشف من الأصل في 27 يناير 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Nieto-Torres, Jose L.؛ DeDiego؛ Verdiá-Báguena؛ Jimenez-Guardeño؛ Regla-Nava؛ Fernandez-Delgado؛ Castaño-Rodriguez؛ Alcaraz؛ Torres (01 مايو 2014)، Denison, Mark R. (المحرر)، "Severe Acute Respiratory Syndrome Coronavirus Envelope Protein Ion Channel Activity Promotes Virus Fitness and Pathogenesis"، PLoS Pathogens، 10 (5): e1004077، doi:10.1371/journal.ppat.1004077، ISSN 1553-7374، PMC 4006877، PMID 24788150، مؤرشف من الأصل في 6 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Wertheim, Joel O.؛ Chu؛ Peiris؛ Kosakovsky Pond؛ Poon (15 يونيو 2013)، "A Case for the Ancient Origin of Coronaviruses"، Journal of Virology، 87 (12): 7039–7045، doi:10.1128/JVI.03273-12، ISSN 0022-538X، PMC 3676139، PMID 23596293، مؤرشف من الأصل في 9 أبريل 2022،
Alphacoronaviruses and betacoronaviruses are found exclusively in mammals, whereas gammacoronaviruses and deltacoronaviruses primarily infect birds.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Cui؛ Li؛ Shi (2018)، "Origin and evolution of pathogenic coronaviruses"، Nature Reviews Microbiology، 17 (3): 181–192، doi:10.1038/s41579-018-0118-9، ISSN 1740-1526.
- Phylogeny of SARS-like betacoronaviruses including novel coronavirus from Wuhan using data generated by the Shanghai Public Health Clinical Center & School of Public Health, the National Institute for Viral Disease Control and Prevention, the Institute of Pathogen Biology, and the Wuhan Institute of Virology shared via GISAID. (16 يناير 2020)..
- Papineau, Amber؛ Berhane؛ Wylie؛ Wylie؛ Sharpe؛ Lung (2019-12)، "Genome Organization of Canada Goose Coronavirus, A Novel Species Identified in a Mass Die-off of Canada Geese"، Scientific Reports، 9 (1): 5954، doi:10.1038/s41598-019-42355-y، ISSN 2045-2322، PMC 6459860، PMID 30976080، مؤرشف من الأصل في 27 يناير 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - 马亦林 (2018)، "冠状病毒的特性及其致病性研究进展"، 中华临床感染病杂志، 11 (4): 305–315، doi:10.3760/cma.j.issn.1674-2397.2018.04.011، مؤرشف من الأصل في 06 مايو 2020، اطلع عليه بتاريخ 21 يناير 2020.
- Cruz, J. L. G.؛ Becares؛ Sola؛ Oliveros؛ Enjuanes؛ Zuniga (01 سبتمبر 2013)، "Alphacoronavirus Protein 7 Modulates Host Innate Immune Response"، Journal of Virology، 87 (17): 9754–9767، doi:10.1128/JVI.01032-13، ISSN 0022-538X، PMC 3754097، PMID 23824792، مؤرشف من الأصل في 11 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Download Virus Metadata Repository: version August 1, 2020; MSL35 (محفوظ). (1 أغسطس 2020).
- Wu, Zhiqiang؛ Lu؛ Du؛ Yang؛ Ren؛ Liu؛ Jiang؛ Yang؛ Dong (2018-12)، "Comparative analysis of rodent and small mammal viromes to better understand the wildlife origin of emerging infectious diseases"، Microbiome، 6 (1): 178، doi:10.1186/s40168-018-0554-9، ISSN 2049-2618، PMC 6171170، PMID 30285857، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - 、Wille, Michelle؛ Holmes (01 سبتمبر 2020)، "Wild birds as reservoirs for diverse and abundant gamma- and deltacoronaviruses"، FEMS Microbiology Reviews، 44 (5): 631–644، doi:10.1093/femsre/fuaa026، ISSN 0168-6445، PMC 7454673، PMID 32672814، مؤرشف من الأصل في 14 مارس 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Woo, P. C. Y.؛ Lau؛ Lam؛ Lau؛ Tsang؛ Lau؛ Bai؛ Teng؛ Tsang (01 أبريل 2012)، "Discovery of Seven Novel Mammalian and Avian Coronaviruses in the Genus Deltacoronavirus Supports Bat Coronaviruses as the Gene Source of Alphacoronavirus and Betacoronavirus and Avian Coronaviruses as the Gene Source of Gammacoronavirus and Deltacoronavirus"، Journal of Virology، 86 (7): 3995–4008، doi:10.1128/JVI.06540-11، ISSN 0022-538X، PMC 3302495، PMID 22278237، مؤرشف من الأصل في 14 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - "Molecular Evolution of Human Coronavirus Genomes"، Trends in Microbiology، 25 (1): 35–48، يناير 2017، doi:10.1016/j.tim.2016.09.001، PMC 7111218، PMID 27743750.
- "Evidence supporting a zoonotic origin of human coronavirus strain NL63"، Journal of Virology، 86 (23): 12816–25، ديسمبر 2012، doi:10.1128/JVI.00906-12، PMC 3497669، PMID 22993147.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Distant relatives of severe acute respiratory syndrome coronavirus and close relatives of human coronavirus 229E in bats, Ghana"، Emerging Infectious Diseases، 15 (9): 1377–84، سبتمبر 2009، doi:10.3201/eid1509.090224، PMC 2819850، PMID 19788804.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Identification and characterization of a novel alpaca respiratory coronavirus most closely related to the human coronavirus 229E"، Viruses، 4 (12): 3689–700، ديسمبر 2012، doi:10.3390/v4123689، PMC 3528286، PMID 23235471.
- "Molecular Evolution of Human Coronavirus Genomes"، Trends in Microbiology، 25 (1): 35–48، يناير 2017، doi:10.1016/j.tim.2016.09.001، PMC 7111218، PMID 27743750.
- "Genetic characterization of Betacoronavirus lineage C viruses in bats reveals marked sequence divergence in the spike protein of pipistrellus bat coronavirus HKU5 in Japanese pipistrelle: implications for the origin of the novel Middle East respiratory syndrome coronavirus"، Journal of Virology، 87 (15): 8638–50، أغسطس 2013، doi:10.1128/JVI.01055-13، PMC 3719811، PMID 23720729.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Evolutionary insights into the ecology of coronaviruses"، Journal of Virology، 81 (8): 4012–20، أبريل 2007، doi:10.1128/jvi.02605-06، PMC 1866124، PMID 17267506.
- "SARS-Coronavirus ancestor's foot-prints in South-East Asian bat colonies and the refuge theory"، Infection, Genetics and Evolution، 11 (7): 1690–702، أكتوبر 2011، doi:10.1016/j.meegid.2011.06.021، PMC 7106191، PMID 21763784.
- "Evolutionary relationships between bat coronaviruses and their hosts"، Emerging Infectious Diseases، 13 (10): 1526–32، أكتوبر 2007، doi:10.3201/eid1310.070448، PMC 2851503، PMID 18258002.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Discovery of a novel coronavirus, China Rattus coronavirus HKU24, from Norway rats supports the murine origin of Betacoronavirus 1 and has implications for the ancestor of Betacoronavirus lineage A"، Journal of Virology، 89 (6): 3076–92، مارس 2015، doi:10.1128/JVI.02420-14، PMC 4337523، PMID 25552712.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Evolutionary dynamics of bovine coronaviruses: natural selection pattern of the spike gene implies adaptive evolution of the strains"، The Journal of General Virology، 94 (Pt 9): 2036–2049، سبتمبر 2013، doi:10.1099/vir.0.054940-0، PMID 23804565،
See Table 1
- "Complete genomic sequence of human coronavirus OC43: molecular clock analysis suggests a relatively recent zoonotic coronavirus transmission event"، Journal of Virology، 79 (3): 1595–604، فبراير 2005، doi:10.1128/jvi.79.3.1595-1604.2005، PMC 544107، PMID 15650185.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Complete genomic sequence of human coronavirus OC43: molecular clock analysis suggests a relatively recent zoonotic coronavirus transmission event"، Journal of Virology، 79 (3): 1595–604، فبراير 2005، doi:10.1128/JVI.79.3.1595-1604.2005، PMC 544107، PMID 15650185.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - Corman, Victor M.؛ Muth؛ Niemeyer؛ Drosten (2018)، "Hosts and Sources of Endemic Human Coronaviruses"، Advances in Virus Research، Elsevier، ج. 100، ص. 163–188، doi:10.1016/bs.aivir.2018.01.001، ISBN 978-0-12-815201-0، PMC 7112090، PMID 29551135، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بكتاب}}: صيانة CS1: تنسيق PMC (link) - "Molecular epidemiology of human coronavirus OC43 reveals evolution of different genotypes over time and recent emergence of a novel genotype due to natural recombination"، Journal of Virology، 85 (21): 11325–37، نوفمبر 2011، doi:10.1128/JVI.05512-11، PMC 3194943، PMID 21849456.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Mouse hepatitis virus infection of the CNS: a model for defense, disease, and repair"، Frontiers in Bioscience، 13 (13): 4393–406، مايو 2008، doi:10.2741/3012، PMC 5025298، PMID 18508518.
- Wang C, Horby PW, Hayden FG, Gao GF (فبراير 2020)، "A novel coronavirus outbreak of global health concern"، Lancet، 395 (10223): 470–473، doi:10.1016/S0140-6736(20)30185-9، PMID 31986257.
- Lau EH, Hsiung CA, Cowling BJ, Chen CH, Ho LM, Tsang T, وآخرون (مارس 2010)، "A comparative epidemiologic analysis of SARS in Hong Kong, Beijing and Taiwan"، BMC Infectious Diseases، 10: 50، doi:10.1186/1471-2334-10-50، PMC 2846944، PMID 20205928.
- "Old age, sepsis tied to poor COVID-19 outcomes, death"، CIDRAP, University of Minnesota، اطلع عليه بتاريخ 29 مارس 2020.
- Karlberg J, Chong DS, Lai WY (فبراير 2004)، "Do men have a higher case fatality rate of severe acute respiratory syndrome than women do?"، American Journal of Epidemiology، 159 (3): 229–31، doi:10.1093/aje/kwh056، PMID 14742282.
- "Summary of probable SARS cases with onset of illness from 1 November 2002 to 31 July 2003"، World Health Organization، أبريل 2004.
- "COVID-19 Dashboard by the Center for Systems Science and Engineering (CSSE) at Johns Hopkins University (JHU)"، ArcGIS، جامعة جونز هوبكينز، اطلع عليه بتاريخ 01 سبتمبر 2022.
- "Report of the WHO-China Joint Mission on Coronavirus Disease 2019 (COVID-19)" (PDF)، World Health Organization، فبراير 2020.
- Oh MD, Park WB, Park SW, Choe PG, Bang JH, Song KH, وآخرون (مارس 2018)، "Middle East respiratory syndrome: what we learned from the 2015 outbreak in the Republic of Korea"، The Korean Journal of Internal Medicine، 33 (2): 233–246، doi:10.3904/kjim.2018.031، PMC 5840604، PMID 29506344.
- Ñamendys-Silva SA (مارس 2020)، "Respiratory support for patients with COVID-19 infection"، The Lancet. Respiratory Medicine، doi:10.1016/S2213-2600(20)30110-7، PMID 32145829.
- "Prevalence and genetic diversity analysis of human coronaviruses among cross-border children"، Virology Journal (باللغة الإنجليزية)، 14 (1): 230، نوفمبر 2017، doi:10.1186/s12985-017-0896-0، PMC 5700739، PMID 29166910.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - "Healthcare-associated atypical pneumonia"، Seminars in Respiratory and Critical Care Medicine، 30 (1): 67–85، فبراير 2009، doi:10.1055/s-0028-1119811، PMID 19199189.
- King, Anthony (02 مايو 2020)، "An uncommon cold"، New Scientist، 246 (3280): 32–35، Bibcode:2020NewSc.246...32K، doi:10.1016/S0262-4079(20)30862-9، ISSN 0262-4079، PMC 7252012، PMID 32501321.
- King, Anthony (2020-05)، "An uncommon cold"، New Scientist، 246 (3280): 32–35، doi:10.1016/S0262-4079(20)30862-9، PMC 7252012، PMID 32501321، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Pelczar، Microbiology:Application Based Approach، Tata McGraw-Hill Education، ص. 656، ISBN 978-0-07-015147-5، مؤرشف من الأصل في 11 أبريل 2022.
- Cecil, Russell La Fayette؛ Goldman؛ Schafer (01 يناير 2012)، Goldman's Cecil Medicine,Expert Consult Premium Edition -- Enhanced Online Features and Print, Single Volume,24: Goldman's Cecil Medicine، Elsevier Health Sciences، ص. 2103–، ISBN 978-1-4377-1604-7، مؤرشف من الأصل في 9 أبريل 2022.
- Charlton, Carmen L.؛ Babady؛ Ginocchio؛ Hatchette؛ Jerris؛ Li؛ Loeffelholz؛ McCarter؛ Miller (19 ديسمبر 2018)، "Practical Guidance for Clinical Microbiology Laboratories: Viruses Causing Acute Respiratory Tract Infections"، Clinical Microbiology Reviews، 32 (1)، doi:10.1128/CMR.00042-18، ISSN 0893-8512، PMC 6302358، PMID 30541871، مؤرشف من الأصل في 8 أبريل 2022،
See Figure 1.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Monto, Arnold S؛ DeJonge؛ Callear؛ Bazzi؛ Capriola؛ Malosh؛ Martin؛ Petrie (16 يونيو 2020)، "Coronavirus Occurrence and Transmission Over 8 Years in the HIVE Cohort of Households in Michigan"، The Journal of Infectious Diseases، 222 (1): 9–16، doi:10.1093/infdis/jiaa161، ISSN 0022-1899، PMC 7184402، PMID 32246136، مؤرشف من الأصل في 13 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Abdul-Rasool, Sahar؛ Fielding (04 يونيو 2010)، "Understanding Human Coronavirus HCoV-NL63~!2009-11-13~!2010-04-09~!2010-05-25~!"، The Open Virology Journal، 4 (1): 76–84، doi:10.2174/1874357901004010076، PMC 2918871، PMID 20700397، مؤرشف من الأصل في 21 مارس 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Vijgen, Leen؛ Keyaerts؛ Moës؛ Thoelen؛ Wollants؛ Lemey؛ Vandamme؛ Van Ranst (2005-02)، "Complete Genomic Sequence of Human Coronavirus OC43: Molecular Clock Analysis Suggests a Relatively Recent Zoonotic Coronavirus Transmission Event"، Journal of Virology، 79 (3): 1595–1604، doi:10.1128/JVI.79.3.1595-1604.2005، ISSN 0022-538X، PMC 544107، PMID 15650185، مؤرشف من الأصل في 26 أبريل 2022،
However, it is tempting to speculate about an alternative hypothesis, that the 1889-1890 pandemic may have been the result of interspecies transmission of bovine coronaviruses to humans, resulting in the subsequent emergence of HCoV-OC43.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Pasley، James. How SARS terrified the world in 2003, infecting more than 8,000 people and killing 774 (محفوظ). مؤرشف من الأصل في 11 مايو 2022. وصل لهذا المسار في 27 يناير 2022.
- Pasley، James. How SARS terrified the world in 2003, infecting more than 8,000 people and killing 774 (محفوظ). مؤرشف من الأصل في 2 مارس 2020. وصل لهذا المسار في 8 نوفمبر 2020.
- Guan, Y.؛ Zheng؛ He؛ Liu؛ Zhuang؛ Cheung؛ Luo؛ Li؛ Zhang (10 أكتوبر 2003)، "Isolation and Characterization of Viruses Related to the SARS Coronavirus from Animals in Southern China"، Science، 302 (5643): 276–278، doi:10.1126/science.1087139، ISSN 0036-8075، مؤرشف من الأصل في 18 أبريل 2022.
- Li, Wendong؛ Shi؛ Yu؛ Ren؛ Smith؛ Epstein؛ Wang؛ Crameri؛ Hu (28 أكتوبر 2005)، "Bats Are Natural Reservoirs of SARS-Like Coronaviruses"، Science، 310 (5748): 676–679، doi:10.1126/science.1118391، ISSN 0036-8075، مؤرشف من الأصل في 28 أبريل 2022.
- Ge, Xing-Yi؛ Hu؛ Shi (26 يونيو 2015)، "Bat Coronaviruses"، في Wang, Lin-Fa (المحرر)، Bats and Viruses، Hoboken, NJ: John Wiley & Sons, Inc، ص. 127–155، doi:10.1002/9781118818824.ch5، ISBN 978-1-118-81882-4، مؤرشف من الأصل في 13 أبريل 2022.
- Hawryluck, Laura؛ Gold؛ Robinson؛ Pogorski؛ Galea؛ Styra (2004-07)، "SARS Control and Psychological Effects of Quarantine, Toronto, Canada"، Emerging Infectious Diseases، 10 (7): 1206–1212، doi:10.3201/eid1007.030703، ISSN 1080-6040، PMC 3323345، PMID 15324539، مؤرشف من الأصل في 27 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Doucleff, Michaeleen (26 سبتمبر 2012)، "Scientists Go Deep On Genes Of SARS-Like Virus"، NPR، مؤرشف من الأصل في 11 مايو 2022، اطلع عليه بتاريخ 27 يناير 2022.
- Falco, Miriam (24 سبتمبر 2012)، "New SARS-like virus poses medical mystery"، CNN Health، مؤرشف من الأصل في 01 نوفمبر 2013، اطلع عليه بتاريخ 16 مارس 2013.
- Middle East respiratory syndrome coronavirus (MERS-CoV) (محفوظ). مؤرشف من الأصل في 18 أبريل 2018. وصل لهذا المسار في 15 أبريل 2020.
- "MERS coronavirus in dromedary camel herd, Saudi Arabia"، Emerging Infectious Diseases، 20 (7): 1231–4، يوليو 2014، doi:10.3201/eid2007.140571، PMC 4073860، PMID 24964193.
- "Rooting the phylogenetic tree of middle East respiratory syndrome coronavirus by characterization of a conspecific virus from an African bat"، Journal of Virology، 88 (19): 11297–303، أكتوبر 2014، doi:10.1128/JVI.01498-14، PMC 4178802، PMID 25031349.
- "Bat origins of MERS-CoV supported by bat coronavirus HKU4 usage of human receptor CD26"، Cell Host & Microbe، 16 (3): 328–37، سبتمبر 2014، doi:10.1016/j.chom.2014.08.009، PMC 7104937، PMID 25211075.
- MERS outbreaks (محفوظ). مؤرشف من الأصل في 11 مايو 2022. وصل لهذا المسار في 7 فبراير 2021.
- Infectious disease outbreaks reported in the Eastern Mediterranean Region in 2018 | News | Epidemic and pandemic diseases (محفوظ). مؤرشف من الأصل في 30 مارس 2022. وصل لهذا المسار في 15 أبريل 2020.
- Assiri, Abdullah؛ McGeer؛ Perl؛ Price؛ Al Rabeeah؛ Cummings؛ Alabdullatif؛ Assad؛ Almulhim (2013-08)، "Hospital Outbreak of Middle East Respiratory Syndrome Coronavirus"، New England Journal of Medicine، 369 (5): 407–416، doi:10.1056/NEJMoa1306742، ISSN 0028-4793، PMC 4029105، PMID 23782161، مؤرشف من الأصل في 27 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Assiri, Abdullah؛ Al-Tawfiq؛ Al-Rabeeah؛ Al-Rabiah؛ Al-Hajjar؛ Al-Barrak؛ Flemban؛ Al-Nassir؛ Balkhy (2013-09)، "Epidemiological, demographic, and clinical characteristics of 47 cases of Middle East respiratory syndrome coronavirus disease from Saudi Arabia: a descriptive study"، The Lancet Infectious Diseases، 13 (9): 752–761، doi:10.1016/S1473-3099(13)70204-4، PMC 7185445، PMID 23891402، مؤرشف من الأصل في 23 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Zumla, Alimuddin؛ Hui؛ Perlman (2015-09)، "Middle East respiratory syndrome"، The Lancet، 386 (9997): 995–1007، doi:10.1016/S0140-6736(15)60454-8، PMC 4721578، PMID 26049252، مؤرشف من الأصل في 25 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - WHO Statement Regarding Cluster of Pneumonia Cases in Wuhan, China (محفوظ). (9 يناير 2020).
- "The continuing 2019-nCoV epidemic threat of novel coronaviruses to global health—The latest 2019 novel coronavirus outbreak in Wuhan, China"، International Journal of Infectious Diseases، 91: 264–66، فبراير 2020، doi:10.1016/j.ijid.2020.01.009، PMC 7128332، PMID 31953166.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - Report of the WHO-China Joint Mission on Coronavirus Disease 2019 (COVID-19) (PDF) (Report)، منظمة الصحة العالمية (WHO)، 24 فبراير 2020، مؤرشف من الأصل (PDF) في 29 فبراير 2020، اطلع عليه بتاريخ 05 مارس 2020.
- "Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding"، Lancet، 395 (10224): 565–574، فبراير 2020، doi:10.1016/S0140-6736(20)30251-8، PMC 7159086، PMID 32007145.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - Temmam؛ Vongphayloth؛ Salazar؛ Munier؛ Bonomi؛ Régnault؛ Douangboubpha؛ Karami؛ Chretien؛ Sanamxay, Daosavanh؛ Xayaphet, Vilakhan (فبراير 2022)، "Bat coronaviruses related to SARS-CoV-2 and infectious for human cells"، Nature: 1–7، doi:10.1038/s41586-022-04532-4، PMID 35172323، S2CID 246902858، مؤرشف من الأصل في 4 مايو 2022.
- "Mystery deepens over animal source of coronavirus"، Nature، 579 (7797): 18–19، مارس 2020، Bibcode:2020Natur.579...18C، doi:10.1038/d41586-020-00548-w، PMID 32127703.
- The Basics of SARS-CoV-2 Transmission، Vancouver, BC: National Collaborating Centre for Environmental Health (NCCEH)، 21 مارس 2021، ISBN 978-1-988234-54-0، مؤرشف من الأصل في 12 مايو 2021، اطلع عليه بتاريخ 12 مايو 2021.
- "The Origins of SARS-CoV-2: A Critical Review"، Cell، 184 (19): 4848–4856، أغسطس 2021، doi:10.1016/j.cell.2021.08.017، PMC 8373617، PMID 34480864.
{{استشهاد بدورية محكمة}}: الوسيط|إظهار المؤلفين=6غير صالح (مساعدة) - The Great Lockdown: Worst Economic Downturn Since the Great Depression (محفوظ). مؤرشف من الأصل في 15 مايو 2020. وصل لهذا المسار في 23 أبريل 2020.
- Oran, Daniel P.؛ Topol (2021-05)، "The Proportion of SARS-CoV-2 Infections That Are Asymptomatic: A Systematic Review"، Annals of Internal Medicine، 174 (5): 655–662، doi:10.7326/M20-6976، ISSN 0003-4819، PMC 7839426، PMID 33481642، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Transmission of COVID-19 (محفوظ). مؤرشف من الأصل في 14 سبتمبر 2020. وصل لهذا المسار في 6 ديسمبر 2020.
- Interim Clinical Guidance for Management of Patients with Confirmed Coronavirus Disease (COVID-19) (محفوظ). (6 أبريل 2020).
- CDC. COVID-19 and Your Health (محفوظ). (11 فبراير 2020).
- "Chapter 24 - Coronaviridae"، Fenner's Veterinary Virology (باللغة الإنجليزية) (ط. Fifth)، Academic Press، 2017، ص. 435–461، doi:10.1016/B978-0-12-800946-8.00024-6، ISBN 978-0-12-800946-8، S2CID 219575461.
- Veterinary Virology، Boston: Academic Press، 1999، ص. 495–508، ISBN 978-0-12-511340-3.
- "Cell replacement therapies to promote remyelination in a viral model of demyelination"، Journal of Neuroimmunology، 224 (1–2): 101–07، يوليو 2010، doi:10.1016/j.jneuroim.2010.05.013، PMC 2919340، PMID 20627412.
- Merck Veterinary Manual (محفوظ). مؤرشف من الأصل في 13 ديسمبر 2019. وصل لهذا المسار في 8 يونيو 2020.
- Bande, Faruku؛ Arshad؛ Hair Bejo؛ Moeini؛ Omar (2015)، "Progress and Challenges toward the Development of Vaccines against Avian Infectious Bronchitis"، Journal of Immunology Research، 2015: 1–12، doi:10.1155/2015/424860، ISSN 2314-8861، PMC 4411447، PMID 25954763، مؤرشف من الأصل في 1 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Cavanagh, Dave (2007-03)، "Coronavirus avian infectious bronchitis virus"، Veterinary Research، 38 (2): 281–297، doi:10.1051/vetres:2006055، ISSN 0928-4249، PMID 17296157، مؤرشف من الأصل في 3 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة) - Wei, Xiaona؛ She؛ Wu؛ Xue؛ Cao (2020-12)، "PEDV enters cells through clathrin-, caveolae-, and lipid raft-mediated endocytosis and traffics via the endo-/lysosome pathway"، Veterinary Research، 51 (1): 10، doi:10.1186/s13567-020-0739-7، ISSN 1297-9716، PMC 7011528، PMID 32041637، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Cruz, Jazmina L. G.؛ Sola؛ Becares؛ Alberca؛ Plana؛ Enjuanes؛ Zuñiga (09 يونيو 2011)، Baric, Ralph S. (المحرر)، "Coronavirus Gene 7 Counteracts Host Defenses and Modulates Virus Virulence"، PLoS Pathogens، 7 (6): e1002090، doi:10.1371/journal.ppat.1002090، ISSN 1553-7374، PMC 3111541، PMID 21695242، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Wang, Qiuhong؛ Vlasova؛ Kenney؛ Saif (2019-02)، "Emerging and re-emerging coronaviruses in pigs"، Current Opinion in Virology، 34: 39–49، doi:10.1016/j.coviro.2018.12.001، PMC 7102852، PMID 30654269، مؤرشف من الأصل في 28 مارس 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Hou, Yixuan؛ Wang (04 نوفمبر 2019)، "Emerging Highly Virulent Porcine Epidemic Diarrhea Virus: Molecular Mechanisms of Attenuation and Rational Design of Live Attenuated Vaccines"، International Journal of Molecular Sciences، 20 (21): 5478، doi:10.3390/ijms20215478، ISSN 1422-0067، PMC 6862049، PMID 31689903، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Zhou, Peng؛ Fan؛ Lan؛ Yang؛ Shi؛ Zhang؛ Zhu؛ Zhang؛ Xie (2018-04)، "Fatal swine acute diarrhoea syndrome caused by an HKU2-related coronavirus of bat origin"، Nature، 556 (7700): 255–258، Bibcode:2018Natur.556..255Z، doi:10.1038/s41586-018-0010-9، ISSN 0028-0836، PMC 7094983، PMID 29618817، مؤرشف من الأصل في 21 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Banerjee, Arinjay؛ Kulcsar؛ Misra؛ Frieman؛ Mossman (09 يناير 2019)، "Bats and Coronaviruses"، Viruses، 11 (1): 41، doi:10.3390/v11010041، ISSN 1999-4915، PMC 6356540، PMID 30634396، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Lau, Susanna K. P.؛ Wong؛ Tsang؛ Ahmed؛ Au-Yeung؛ Yuen؛ Wernery؛ Woo (2018-08)، Gallagher, Tom (المحرر)، "Discovery and Sequence Analysis of Four Deltacoronaviruses from Birds in the Middle East Reveal Interspecies Jumping with Recombination as a Potential Mechanism for Avian-to-Avian and Avian-to-Mammalian Transmission"، Journal of Virology، 92 (15)، doi:10.1128/JVI.00265-18، ISSN 0022-538X، PMC 6052312، PMID 29769348، مؤرشف من الأصل في 9 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Taxonomy browser (Betacoronavirus 1) (محفوظ). مؤرشف من الأصل في 6 أكتوبر 2020. وصل لهذا المسار في 8 يونيو 2020.
- Mattei؛ Kopper؛ Sanz (2020)، "Equine Coronavirus-Associated Colitis in Horses: A Retrospective Study"، Journal of Equine Veterinary Science، 87: 102906، doi:10.1016/j.jevs.2019.102906، ISSN 0737-0806.
- Equine Enteric Coronavirus (محفوظ). مؤرشف من الأصل في 5 فبراير 2021. وصل لهذا المسار في 30 يناير 2021.
- Taxonomy browser (Alphacoronavirus 1) (محفوظ). مؤرشف من الأصل في 5 أكتوبر 2020. وصل لهذا المسار في 8 يونيو 2020.
- Addie, Diane؛ Belák؛ Boucraut-Baralon؛ Egberink؛ Frymus؛ Gruffydd-Jones؛ Hartmann؛ Hosie؛ Lloret (2009-07)، "Feline Infectious Peritonitis: ABCD Guidelines on Prevention and Management"، Journal of Feline Medicine and Surgery، 11 (7): 594–604، doi:10.1016/j.jfms.2009.05.008، ISSN 1098-612X، PMC 7129471، PMID 19481039، مؤرشف من الأصل في 15 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Pedersen, Niels C. (2014-08)، "An update on feline infectious peritonitis: Diagnostics and therapeutics"، The Veterinary Journal، 201 (2): 133–141، doi:10.1016/j.tvjl.2014.04.016، PMC 7110619، PMID 24857253، مؤرشف من الأصل في 18 مارس 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Taxonomy browser (Alphacoronavirus) (محفوظ). مؤرشف من الأصل في 5 أكتوبر 2020. وصل لهذا المسار في 8 يونيو 2020.
- Murray، Jerry. What's New With Ferret FIP-like Disease? (محفوظ). (xls) (16 أبريل 2014).
- Infectious Diseases of Ferrets - Exotic and Laboratory Animals (محفوظ). مؤرشف من الأصل في 28 نوفمبر 2020. وصل لهذا المسار في 8 يونيو 2020.
- Körner, Robert؛ Majjouti؛ Alcazar؛ Mahabir (12 أغسطس 2020)، "Of Mice and Men: The Coronavirus MHV and Mouse Models as a Translational Approach to Understand SARS-CoV-2"، Viruses، 12 (8): 880، doi:10.3390/v12080880، ISSN 1999-4915، PMC 7471983، PMID 32806708، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Weiss, Susan R.؛ Navas-Martin (2005-12)، "Coronavirus Pathogenesis and the Emerging Pathogen Severe Acute Respiratory Syndrome Coronavirus"، Microbiology and Molecular Biology Reviews، 69 (4): 635–664، doi:10.1128/MMBR.69.4.635-664.2005، ISSN 1092-2172، PMC 1306801، PMID 16339739، مؤرشف من الأصل في 7 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Mouse Hepatitis Virus (MHV) (محفوظ). مؤرشف من الأصل في 28 ديسمبر 2015. وصل لهذا المسار في 21 مارس 2020.
- Yoo, Dongwan؛ Pei؛ Christie؛ Cooper (2000-07)، "Primary Structure of the Sialodacryoadenitis Virus Genome: Sequence of the Structural-Protein Region and Its Application for Differential Diagnosis"، Clinical Diagnostic Laboratory Immunology، 7 (4): 568–573، doi:10.1128/CDLI.7.4.568-573.2000، ISSN 1071-412X، PMC 95915، PMID 10882653، مؤرشف من الأصل في 22 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Enteric Coronavirus (محفوظ). مؤرشف من الأصل في 1 يوليو 2019. وصل لهذا المسار في 24 يناير 2020.
- Small, J. David؛ Woods (1987)، "Relatedness of Rabbit Coronavirus to Other Coronaviruses"، في Lai, Michael M. C. (المحرر)، Coronaviruses، Boston, MA: Springer US، ج. 218، ص. 521–527، doi:10.1007/978-1-4684-1280-2_68، ISBN 978-1-4684-1282-6، مؤرشف من الأصل في 19 أبريل 2022.
- N. James MacLachlan and Edward J. Dubovi, المحرر (2017)، "Coronaviridae"، Fenner's Veterinary Virology (Fifth Edition)، ص. 435–461، doi:10.1016/B978-0-12-800946-8.00024-6.
- Lau, S. K. P.؛ Woo؛ Yip؛ Fan؛ Huang؛ Wang؛ Guo؛ Lam؛ Tsang (15 مايو 2012)، "Isolation and Characterization of a Novel Betacoronavirus Subgroup A Coronavirus, Rabbit Coronavirus HKU14, from Domestic Rabbits"، Journal of Virology، 86 (10): 5481–5496، doi:10.1128/JVI.06927-11، ISSN 0022-538X، PMC 3347282، PMID 22398294، مؤرشف من الأصل في 8 مايو 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Domańska-Blicharz, Katarzyna؛ Woźniakowski؛ Konopka؛ Niemczuk؛ Welz؛ Rola؛ Socha؛ Orłowska؛ Antas (02 أغسطس 2020)، "Animal coronaviruses in the light of COVID-19"، Journal of Veterinary Research، 64 (3): 333–345، doi:10.2478/jvetres-2020-0050، ISSN 2450-8608، PMC 7497757، PMID 32984621، مؤرشف من الأصل في 15 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Monchatre-Leroy, Elodie؛ Boué؛ Boucher؛ Renault؛ Moutou؛ Ar Gouilh؛ Umhang (29 نوفمبر 2017)، "Identification of Alpha and Beta Coronavirus in Wildlife Species in France: Bats, Rodents, Rabbits, and Hedgehogs"، Viruses، 9 (12): 364، doi:10.3390/v9120364، ISSN 1999-4915، PMC 5744139، PMID 29186061، مؤرشف من الأصل في 2 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Amer, Haitham Mohamed (2018-12)، "Bovine-like coronaviruses in domestic and wild ruminants"، Animal Health Research Reviews، 19 (2): 113–124، doi:10.1017/S1466252318000117، ISSN 1466-2523، PMC 7108644، PMID 30683171، مؤرشف من الأصل في 18 أبريل 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - Mordecai, Gideon J.؛ Hewson (24 يوليو 2020)، "Coronaviruses in the Sea"، Frontiers in Microbiology، 11: 1795، doi:10.3389/fmicb.2020.01795، ISSN 1664-302X، PMC 7393285، PMID 32793180، مؤرشف من الأصل في 27 مارس 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Shi, Mang؛ Lin؛ Chen؛ Tian؛ Chen؛ Li؛ Wang؛ Eden؛ Shen (2018-04)، "The evolutionary history of vertebrate RNA viruses"، Nature، 556 (7700): 197–202، doi:10.1038/s41586-018-0012-7، ISSN 0028-0836، مؤرشف من الأصل في 11 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة) - Common Human Coronaviruses (محفوظ). مؤرشف من الأصل في 11 أبريل 2020. وصل لهذا المسار في 31 يناير 2021.
- Jiang, Shibo؛ Lu؛ Du (2013-01)، "Development of SARS vaccines and therapeutics is still needed"، Future Virology، 8 (1): 1–2، doi:10.2217/fvl.12.126، ISSN 1746-0794، PMC 7079997، PMID 32201503، مؤرشف من الأصل في 10 مايو 2022.
{{استشهاد بدورية محكمة}}: تحقق من التاريخ في:|تاريخ=(مساعدة)صيانة CS1: تنسيق PMC (link) - SARS (severe acute respiratory syndrome) (محفوظ). (19 أكتوبر 2017).
- MERS Clinical Features (محفوظ). (2 أغسطس 2019).
- Offeddu, Vittoria؛ Yung؛ Low؛ Tam (13 نوفمبر 2017)، "Effectiveness of Masks and Respirators Against Respiratory Infections in Healthcare Workers: A Systematic Review and Meta-Analysis"، Clinical Infectious Diseases، 65 (11): 1934–1942، doi:10.1093/cid/cix681، ISSN 1058-4838، PMC 7108111، PMID 29140516، مؤرشف من الأصل في 22 أبريل 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - Consensus document on the epidemiology of severe acute respiratory syndrome (SARS)، World Health Organization، 2003، hdl:10665/70863.
- Nussbaumer-Streit, Barbara؛ Mayr؛ Dobrescu؛ Chapman؛ Persad؛ Klerings؛ Wagner؛ Siebert؛ Christof (08 أبريل 2020)، Cochrane Infectious Diseases Group (المحرر)، "Quarantine alone or in combination with other public health measures to control COVID-19: a rapid review"، Cochrane Database of Systematic Reviews، doi:10.1002/14651858.CD013574، PMC 7141753، PMID 32267544، مؤرشف من الأصل في 20 مايو 2022.
{{استشهاد بدورية محكمة}}: صيانة CS1: تنسيق PMC (link) - نصيحة لعامة الناس بخصوص فيروس كوفيد -19: القيام بالتطعيم (محفوظ). مؤرشف من الأصل في 11 مايو 2022. وصل لهذا المسار في 20 أبريل 2022.
- Transboundary and Emerging Diseases of Animals، Iowa State University، 2016، ISBN 978-0-9846270-5-9.
باللغة العربيَّة
- ترجمة (coronavirus) على موقع القاموس (محفوظ). مؤرشف من الأصل في 3 فبراير 2020. وصل لهذا المسار في 26 يناير 2020.
- ترجمة (coronavirus) في المعجم الطبي الموحد (محفوظ). مؤرشف من الأصل في 26 يناير 2020. وصل لهذا المسار في 26 يناير 2020.
- ترجمة (coronavirus) في المعجم الصيدلي الموحد (محفوظ). مؤرشف من الأصل في 26 يناير 2020. وصل لهذا المسار في 26 يناير 2020.
- يُوسف حِتّي؛ أحمَد شفيق الخَطيب (2008)، قاموس حتي الطبي للجيب، صفحة=102، بيروت، لبنان: مكتبة لبنان ناشرون، ISBN 995310235X، مؤرشف من الأصل في 21 أبريل 2022، اطلع عليه بتاريخ 27-01 2020.
{{استشهاد بكتاب}}: تحقق من التاريخ في:|تاريخ الوصول=(مساعدة) - ترجمة (coronavirus) في معجم مرعشي الطبي الكبير (محفوظ). مؤرشف من الأصل في 26 يناير 2020. وصل لهذا المسار في 26 يناير 2020.
- ترجمة (coronavirus) في القاموس الطبي (محفوظ). مؤرشف من الأصل في 26 يناير 2020. وصل لهذا المسار في 26 يناير 2020.
مراجع إضافية
- "Novel coronavirus infection: time to stay ahead of the curve"، Eastern Mediterranean Health Journal، 19 Suppl 1: S3–4، 2013، PMID 23888787، مؤرشف من الأصل في 24 مارس 2020.
- "Molecular biology of transmissible gastroenteritis virus"، Veterinary Microbiology، 23 (1–4): 147–54، يونيو 1990، doi:10.1016/0378-1135(90)90144-K، PMID 2169670.
- "Engineering the transmissible gastroenteritis virus genome as an expression vector inducing lactogenic immunity"، Journal of Virology، 77 (7): 4357–69، أبريل 2003، doi:10.1128/JVI.77.7.4357-4369.2003، PMC 150661، PMID 12634392.
- "Morphology of transmissible gastroenteritis virus of pigs. A possible member of coronaviruses. Brief report"، Archiv für die Gesamte Virusforschung، 29 (1): 105–8، 1970، doi:10.1007/BF01253886، PMID 4195092.
روابط خارجية
- قاعدة بيانات وتحليل مصادر الفيروسات الممرضة (ViPR): الفيروسات التاجية
- مؤسسة الأبحاث الألمانية (Coronavirus Consortium)
- ما الصواب في ترجمة مصطلح Coronavirus؟.
 بوابة علم الفيروسات
بوابة علم الفيروسات بوابة جائحة فيروس كورونا 2019–20
بوابة جائحة فيروس كورونا 2019–20