ORF8
ORF8 هو جين يُشفر البروتين الفيروسي الملحق بروتين NS8 لفيروس كورونا بيتا، ويتواجد في جنيس فيروسات كورونا البائية السارسية (Sarbecovirus). وهو أحد أقل الجينات انحفاظا وأكثرها تغيرا في الجينوم.[2][3][4][5] في بعض الفيروسات، قسمت طفرة حذف منطقة الجين إلى إطاري قراءة مفتوح صغيرين سُميا: ORF8a وORF8b وهي ميزة في العديد من عينات سارس-كوف المعزولة لاحقا من تفشي السارس، وكذلك من بعض فيروسات كورونا الخفاشية.[3][4] لهذا السبب يُسمى الطول الكامل للجين في بعض الأحيان ORF8ab.[3][6] يشفر الجين كامل الطول في سارس-كوف-2 بروتينا يحتوي على نطاق غلوبولين مناعي [الإنجليزية] غير معروف الوظيفة، ربما له دور في التآثرات مع جهاز مناعة المضيف.[1][3][4] يتشابه بروتين هذا الجين في البنية مع بروتين ORF7a ما يوحي بإمكانية نشوئه عبر تضاعف جيني.[7][8]
| بروتين NS8 لفيروس كورونا بيتا | |
|---|---|
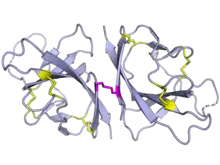 | |
| مثنوي بروتين ORF8 الخاص بسارس-كوف-2. روابط ثنائي الكبريتيد الداخلية ظاهرة بالأصفر، وبين الجزيئية بالوردي. من ببب: 7JTL.[1] | |
| معرف | |
| رمز | bCoV_NS8 |
| قاعدة بيانات عوائل البروتينات | PF12093 |
| إنتربرو | IPR022722 |
البنية
يُشفر الـORF8 في سارس-كوف-2 بروتينا طوله 121 حمض أميني يحتوي على ببتيد إشعاري في النهاية الأمينية.[4] ويشكل مثنويا مترابط تساهميا عبر روابط ثنائي كبريتيد.[1] ويملك نطاقا مشابها لنطاق الغلوبولين المناعي له تماثل بعيد مع بروتين ORF7a.[1][2] رغم التطوي المتماثل إجماليا، فإن طفرة غرز في الـORF8 على الأرجح مسؤولة على تآثرات بروتين بروتين مختلفة وتُحدِث واجهة تثني إضافية.[1][2] على عكس الـORF7a، لا يملك الـORF8 لولبا عبر غشائي وبالتالي ليس بروتينا عبر غشائي،[1][4] لكن اقتُرح أنه ربما يملك هيئة إرساء في الغشاء.[3]
الـORF8 في سارس-كوف وسارس-كوف-2 متباعد [الإنجليزية] ومتباين وله تماثل تسلسل أقل من 20% بينهما.[1] يشفر الـORF8 ذو الطول الكامل في سارس-كوف بروتينا طوله 122 حمض أميني، وهو منقسم في العديد من مستخلصات سارس-كوف إلى ORF8a وORF8b ويشفران بروتينين طولهما 39 حمض أميني و81 حمض أميني على التوالي.[6] اقتُرح أن بروتيني ORF8a وORF8b قد يشكلان مركبا بروتينيا.[2][9] الحمض الأميني سيستئين المسؤول على تثني بروتين سارس-كوف-2 غير منحفظ في تسلسل سارس-كوف.[1] أُبلغ كذلك أن بروتين ORF8ab يشكل أيضا بروتينا متعدد القسيمات مرتبطا بروابط ثنائي كبريتيد.[10]
تعديلات ما بعد الترجمة
يُعدل بروتين ORF8ab كامل الطول الخاص بسارس-كوف بعد الترجمة عبر غلكزة N،[6] وهو الأمر الذي يُعتقد أنه محفوظ لدى بروتين سارس-كوف-2.[1] تحت الظروف التجريبة، يتم ربط كل من 8b و8ab باليوبيكويتين.[6]
التعبير والتموقع
إلى جانب جينات البروتينات الملحقة الأخرى، يتواجد جين ORF8 بالقرب من الجينات التي تشفر البروتينات البنيوية في النهاية 3' لجينوم فيروسات كورونا الرنوي. إلى جانب كل من ORF6 وORF7a وORF7b، يتموضع ORF8 بين جيني الغشاء والقفيصة النووية.[4][6] يملك بروتين ORF8 الخاص بسارس-كوف-2 ببتيد إشعاري لنقله إلى الشبكة الإندوبلازمية وتم نقله في الظروف المختبرية إلى الشبكة الإندوبلازمية كذلك.[11] وعلى الأرجح هو بروتين يتم إفرازه خارج الخلية.[3][4]
توجد تقارير مختلفة في المنشورات العلمية بخصوص تموقع بروتينات 8a و8b و8ab الخاصة بسارس-كوف.[6] وغير واضح ما إذا كان يُعبر عن ORF8b بمستويات معتبرة تحت الظروف الطبيعية.[10][12] يبدو أن الـORF8ab كامل الطول يتموقع في الشبكة الإندوبلازمية.[12]
الوظيفة
وظيفة بروتين ORF8 غير معروفة، وهو ليس أساسيا لتضاعف الفيروس سواء في سارس-كوف[6] أو سارس-كوف-2،[4] ومع ذلك توجد دلائل متضاربة حول ما إذا كان فقدان ORF8 يؤثر على فعالية تكاثر الفيروس.[13]
أحد الوظائف المفترضة لبروتين ORF8 هي التآثر مع جهاز مناعة المضيف.[13] يُعتقد أن لبروتين سارس-كوف-2 دور في تعديل المناعة عبر التهرب المناعي أو تثبيط الاستجابات المناعية.[1][3][4] أُبلغ أنه مناهض للإنترفيرون نوع 1 ويخفض التعبير عن الصنف 1 من معقد التوافق النسيجي الكبير [الإنجليزية].[3][4] بروتين ORF8 الخاص بسارس-كوف-2 شديد توليد المناعة ووُجدت مستويات عالية من الأجسام المضادة له لدى المرضى المتعافين من كوفيد 19.[4][14]
اقتُرح أن بروتين ORF8a الخاص بسارس كوف يتجمع إلى عديد قسيمات ويشكل فيروبورين.[15]
التطور

التاريخ التطوري لـORF8 معقد، ويُعتبر أحد أقل المناطق انحفاظا في جينوم فيروسات كورونا البائية السارسية.[2][3][4] وهو عرضة لطفرات وحذوفات ووصف بأنه «مفرط التغير» وأنه منطقة تأشيب ساخنة [الإنجليزية].[3] اقتُرح وجود بُنى رنا ثانوية في تلك المنطقة لها علاقة بعدم استقرار الجينوم.[3][17]
في سارس-كوف، يُعتقد بأن منطقة الـORF8 نشأت عبر تأشيب بين أسلاف فيروسات كورونا الخفاشية.[3][5][6][18] من بين الميزات الأكثر تجليا في هذه المنطقة في سارس-كوف هي ظهور حذف لـ29 نوكليوتيد تسبب في انقسام إطار القراءة المفتوح كامل الطول إلى إطارين صغيرين هما: ORF8a وORF8b. المستخلصات الفيروسية (العينات المعزولة) من بداية وباء السارس تملك الطول الكامل للإطار ORF8، لكن ظهرت البنية المنقسمة له في فترة لاحقة من الوباء.[3][6] ولوحظت بُنى مماثلة منذ ذلك الوقت في فيروسات كورونا خفاشية أخرى.[19] لوحظت طفرات وحذوفات أيضا في متحورات سارس-كوف-2.[2][17] استنادا على ملاحظات في سارس-كوف، اقتُرح أن التغيرات في الـORF8 يمكن أن تكون لها علاقة بالتكيف مع المضيف، لكن من الممكن أن ORF8 لا يؤثر على الصلاحية في المضيفين البشر.[5][17] في سارس-كوف، لوحظ معدل Ka/Ks [الإنجليزية] عالٍ في الـORF8 يتوافق مع اصطفاء إيجابي أو مع اصطفاء خالٍ من الضغط.[5]
يُشفر الـORF8 بروتينا له نطاق غلوبولين مناعي [الإنجليزية] ذو صلة بعيد بنطاق ORF7a.[1] اقتُرح أن الـORF8 على الأرجح تطور من الـORF7a عبر تضاعف جيني،[2][7][8] إلا أن بعض دراسات المعلوماتية الحيوية تقترح أن كمية التشابه قد تكون منخفضة لدرجة لا تدعم حدوث تضاعف جيني والذي هو أمر غير شائع نسبيا لدى الفيروسات.[17] نطاقات الغلوبولينات المناعية غير شائعة بين الفيروسات وباستثناء مجموعة فرعية في فيروسات كورونا بيتا تملك ORF8 وORF7a، تم تحديد عدد صغير من فيروسات كورونا ألفا الخفاشية التي تملك نطاقات مماثلة لنطاق الغلوبولين المناعي، بينما هذا النطاق غائب في فيروسات غاما وفيروسات دلتا.[2][8] الـORF8 غائب في ميرس-كوف.[8] يمكن أن يكون اكتساب نطاقات الغلوبولين المناعي في فيروسات بيتا وغاما حدث بشكل مستقل، بينما ORF8 وORF7a يمكن أن يكون مصدرها من بروتين خاص بالمضيف.[2] من الممكن كذلك أن غياب الـORF8 يعكس فقدانا للجين في تلك الخطوط التطورية.[8]
مراجع
- Flower؛ Buffalo؛ Hooy؛ Allaire؛ Ren؛ Hurley (12 يناير 2021)، "Structure of SARS-CoV-2 ORF8, a rapidly evolving immune evasion protein"، Proceedings of the National Academy of Sciences، 118 (2): e2021785118، doi:10.1073/pnas.2021785118، PMC 7812859، PMID 33361333.
- Tan؛ Schneider؛ Leong؛ Aravind؛ Zhang (30 يونيو 2020)، "Novel Immunoglobulin Domain Proteins Provide Insights into Evolution and Pathogenesis of SARS-CoV-2-Related Viruses"، mBio، 11 (3)، doi:10.1128/mBio.00760-20، PMC 7267882، PMID 32471829.
- Zinzula (يناير 2021)، "Lost in deletion: The enigmatic ORF8 protein of SARS-CoV-2"، Biochemical and Biophysical Research Communications، 538: 116–124، doi:10.1016/j.bbrc.2020.10.045، PMC 7577707، PMID 33685621.
- Redondo؛ Zaldívar-López؛ Garrido؛ Montoya (07 يوليو 2021)، "SARS-CoV-2 Accessory Proteins in Viral Pathogenesis: Knowns and Unknowns"، Frontiers in Immunology، 12: 708264، doi:10.3389/fimmu.2021.708264، hdl:10261/249329، PMC 8293742، PMID 34305949.
- Forni؛ Cagliani؛ Clerici؛ Sironi (يناير 2017)، "Molecular Evolution of Human Coronavirus Genomes"، Trends in Microbiology، 25 (1): 35–48، doi:10.1016/j.tim.2016.09.001، PMC 7111218، PMID 27743750.
- Liu؛ Fung؛ Chong؛ Shukla؛ Hilgenfeld (سبتمبر 2014)، "Accessory proteins of SARS-CoV and other coronaviruses."، Antiviral Research، 109: 97–109، doi:10.1016/j.antiviral.2014.06.013، PMC 7113789، PMID 24995382.
- Mariano؛ Farthing؛ Lale-Farjat؛ Bergeron (17 ديسمبر 2020)، "Structural Characterization of SARS-CoV-2: Where We Are, and Where We Need to Be"، Frontiers in Molecular Biosciences، 7: 605236، doi:10.3389/fmolb.2020.605236، PMC 7773825، PMID 33392262.
- Neches؛ Kyrpides؛ Ouzounis (23 فبراير 2021)، "Atypical Divergence of SARS-CoV-2 Orf8 from Orf7a within the Coronavirus Lineage Suggests Potential Stealthy Viral Strategies in Immune Evasion"، mBio، 12 (1)، doi:10.1128/mBio.03014-20، PMC 7845636، PMID 33468697.
- von Brunn؛ Teepe؛ Simpson؛ Pepperkok؛ Friedel؛ Zimmer؛ Roberts؛ Baric؛ Haas (23 مايو 2007)، "Analysis of Intraviral Protein-Protein Interactions of the SARS Coronavirus ORFeome"، PLOS ONE، 2 (5): e459، Bibcode:2007PLoSO...2..459V، doi:10.1371/journal.pone.0000459، PMC 1868897، PMID 17520018.
- Oostra؛ de Haan؛ Rottier (15 ديسمبر 2007)، "The 29-Nucleotide Deletion Present in Human but Not in Animal Severe Acute Respiratory Syndrome Coronaviruses Disrupts the Functional Expression of Open Reading Frame 8"، Journal of Virology، 81 (24): 13876–13888، doi:10.1128/JVI.01631-07، PMC 2168875، PMID 17928347.
- Zhang؛ Cruz-cosme؛ Zhuang؛ Liu؛ Liu؛ Teng؛ Wang؛ Tang (ديسمبر 2020)، "A systemic and molecular study of subcellular localization of SARS-CoV-2 proteins"، Signal Transduction and Targeted Therapy، 5 (1): 269، doi:10.1038/s41392-020-00372-8، PMC 7670843، PMID 33203855.
- McBride؛ Fielding (07 نوفمبر 2012)، "The Role of Severe Acute Respiratory Syndrome (SARS)-Coronavirus Accessory Proteins in Virus Pathogenesis"، Viruses، 4 (11): 2902–2923، doi:10.3390/v4112902، PMC 3509677، PMID 23202509.
- Mohammad؛ Bouchama؛ Mohammad Alharbi؛ Rashid؛ Saleem Khatlani؛ Gaber؛ Malik (20 أغسطس 2020)، "SARS-CoV-2 ORF8 and SARS-CoV ORF8ab: Genomic Divergence and Functional Convergence"، Pathogens، 9 (9): 677، doi:10.3390/pathogens9090677، PMC 7558349، PMID 32825438.
- Hachim؛ Kavian؛ Cohen؛ Chin؛ Chu؛ Mok؛ Tsang؛ Yeung؛ Perera؛ Poon, Leo L. M.؛ Peiris, J. S. Malik؛ Valkenburg, Sophie A. (أكتوبر 2020)، "ORF8 and ORF3b antibodies are accurate serological markers of early and late SARS-CoV-2 infection"، Nature Immunology، 21 (10): 1293–1301، doi:10.1038/s41590-020-0773-7، PMID 32807944، S2CID 221136730.
- Chen؛ Krüger؛ Sramala؛ Hsu؛ Henklein؛ Chen؛ Fischer (فبراير 2011)، "ORF8a of SARS-CoV forms an ion channel: Experiments and molecular dynamics simulations"، Biochimica et Biophysica Acta (BBA) - Biomembranes، 1808 (2): 572–579، doi:10.1016/j.bbamem.2010.08.004، PMC 7094593، PMID 20708597.
- Zhou؛ Huang؛ Zhou؛ Huang؛ Su؛ Kang؛ Chen؛ Chen؛ He؛ Rong, Xia؛ Xiao, Fei؛ Chen, Jun؛ Chen, Shoudeng (مارس 2021)، "Structural insight reveals SARS-CoV-2 ORF7a as an immunomodulating factor for human CD14+ monocytes"، iScience، 24 (3): 102187، Bibcode:2021iSci...24j2187Z، doi:10.1016/j.isci.2021.102187، PMC 7879101، PMID 33615195.
- Pereira (نوفمبر 2020)، "Evolutionary dynamics of the SARS-CoV-2 ORF8 accessory gene"، Infection, Genetics and Evolution، 85: 104525، doi:10.1016/j.meegid.2020.104525، PMC 7467077، PMID 32890763.
- Lau؛ Feng؛ Chen؛ Luk؛ Yang؛ Li؛ Zhang؛ Huang؛ Song؛ Chow, Wang-Ngai؛ Fan, Rachel Y. Y.؛ Ahmed, Syed Shakeel؛ Yeung, Hazel C.؛ Lam, Carol S. F.؛ Cai, Jian-Piao؛ Wong, Samson S. Y.؛ Chan, Jasper F. W.؛ Yuen, Kwok-Yung؛ Zhang, Hai-Lin؛ Woo, Patrick C. Y. (15 أكتوبر 2015)، "Severe Acute Respiratory Syndrome (SARS) Coronavirus ORF8 Protein Is Acquired from SARS-Related Coronavirus from Greater Horseshoe Bats through Recombination"، Journal of Virology، 89 (20): 10532–10547، doi:10.1128/JVI.01048-15، PMC 4580176، PMID 26269185.
- Hu؛ Zeng؛ Yang؛ Ge؛ Zhang؛ Li؛ Xie؛ Shen؛ Zhang؛ Wang, Ning؛ Luo, Dong-Sheng؛ Zheng, Xiao-Shuang؛ Wang, Mei-Niang؛ Daszak, Peter؛ Wang, Lin-Fa؛ Cui, Jie؛ Shi, Zheng-Li (30 نوفمبر 2017)، "Discovery of a rich gene pool of bat SARS-related coronaviruses provides new insights into the origin of SARS coronavirus"، PLOS Pathogens، 13 (11): e1006698، doi:10.1371/journal.ppat.1006698، PMC 5708621، PMID 29190287.
جينومات فيروس كورونا | ||
|---|---|---|
| بروتين بنيوي فيروسي |
|  |
| بروتين لابنيوي فيروسي (معبر عنه من الـ ORF1ab) |
| |
| بروتين ملحق فيروسي | ||
| رنا | ||
 بوابة علم الوراثة
بوابة علم الوراثة بوابة الكيمياء الحيوية
بوابة الكيمياء الحيوية بوابة علم الفيروسات
بوابة علم الفيروسات