بروتين القفيصة النووية لفيروس كورونا
بروتين القفيصة النووية لفيروس كورونا (بالإنجليزية: Coronavirus nucleocapsid protein)؛ هو بروتين يغلف المادة الوراثية للحمض النووي الريبي مفرد السلسلة موجب الدلالة الخاص بفيروس كورونا لتشكيل بنى بروتين نووي ريبوزي داخل القفيصة الفيروسية.[2][3] بروتين القفيصة النووية أحد البروتينات الأساسية بين البروتينات الهيكلية الأربعة الرئيسة التي تُكوّن فيروسات فيروس كورونا، وأكثر البروتينات التي يتم التعبير عنها.[3][4] بالإضافة إلى تفاعلاته مع الحمض النووي الريبي، يشكل بروتين القفيصة النووية تفاعلات بروتينية-بروتينية مع البروتين الغشائي (M) أثناء عملية التجميع الفيروسي.[2][3] ويعد بروتين القفيصة النووية مولدا كبيرا للمناعة، وهو من البروتينات التي يتم استهدافها بواسطة المضادات الفيروسية التي تسهدف الفيروس، وقد وُجدت أجسام مضادة ضده لدى المرضى الذين تعافوا من مرضي سارس ومرض فيروس كورونا.[5]
| بروتين القفيصة النووية لفيروس كورونا | |
|---|---|
 | |
| نموذج للهيكل الخارجي لفيروس سارس-كوف-2.[1] ● الأزرق: غلاف الفيروس ● الفيروزي: البروتين الشوكي لفيروس كورونا (S) ● الأحمر: البروتين الغلافي لفيروس كورونا (E) ● الأخضر: البروتين الغشائي لفيروس كورونا (M) ● البرتقالي: جليكان | |
| معرف | |
| رمز | CoV_nucleocap |
| قاعدة بيانات عوائل البروتينات | PF00937 |
| إنتربرو | IPR001218 |
البنية
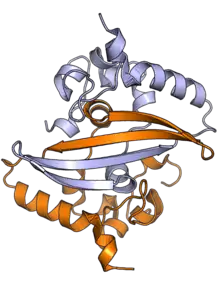
يتكون بروتين القفيصة من نطاقين رئيسيين متصلين عبر منطقة مضطربة جوهريا (IDR) مع وجود مناطق مضطربة إضافية في كل نهاية.[2][3] يبدو أن لنطاق ثالثٍ صغير بالنهاية الكربوكسيلية بنية لولب ألفا ثانوية منتظمة يمكن أن يكون لها دور في تشكيل تجميعات قليل قسيمات عالي الرتبة.[6] في سارس-كوف العامل المسبب للسارس، طول بروتين القفيصة هو 422 حمض أميني[2] وفي سارس-كوف-2 -العامل المسبب لكوفيد 19- طوله 419 حمض أميني.[6][7]
يستطيع كلا نطاقي النهايتين الأمينية والكربوكسيلية الارتباط بالرنا. يشكل نطاق النهاية الأمينية مثنويا هو على الأرجح الهيئة الوظيفية الطبيعية.[2] يمكن كذلك أن يكون لأجزاءٍ من المنطقة المضطربة جوهريا -بالتحديد طابع تسلسل [الإنجليزية] محفوظ غني بواحدات السيرين والأرجنين (المنطقة الغنية بالسيرين)- دور في تشكُل المثنوي، رغم أن التقارير مختلفة بهذا الشأن.[2][3] مع أن قليلات القسيمات عالية الرتبة المشكلة عبر نطاقات النهايات الكربوكسيلية تمت ملاحظتها بواسطة علم البلورات السيني، إلا أنه غير واضح ما إذا كان لهذه البُنى وظيفة فسيولوجية.[2][8]
حُددت الخصائص البنيوية لمثنوي النهاية الكربوكسيلية بواسطة علم البلورات السيني للعديد من فيروسات كورونا، ويملك هذا المثنوي بنية عالية الانحفاظ.[6] تمت بلورة نطاق النهاية الأمينية كذلك -يُعرف أحيانا بنطاق الارتباط بالرنا، مع أن أجزاء أخرى من هذا البروتين تتأثر مع الرنا أيضا- وتمت دراسته بواسطة مطيافية الرنين المغناطيسي النووي للبروتين في حالة تواجد الرنا.[9]
تعديلات ما بعد الترجمة
يُعدَّل بروتين القفيصة بعد الترجمة بالفسفرة في مواضع تقع في المنطقة المضطربة جوهريا، وبالتحديد في المنطقة الغنية بالسيرين.[2][10] أُبلغ في العديد من فيروسات كورونا عن ربزلة-ADP [الإنجليزية] لبروتين القفيصة.[10][11] لوحظ أن بروتين القفيصة في سارس-كوف تتم إضافة بروتين السومو [الإنجليزية] له من دون أي أهمية وظيفية واضحة. لوحظ أن بروتينات القفيصة الخاصة بالعديد من فيروسات كورونا بما في ذلك سارس-كوف-2 يتم قصها عبر التحلل البروتيني.[10][12][13]
التعبير والتموقع
بروتين القفيصة النووية هو أكثر البروتينات المعبر عنها بغزارة بين البروتينات البنيوية الأربع الرئيسية.[2] مثل بقية البروتينات البنيوية الأخرى، يتواجد الجين المشفر لبروتين القفيصة النووية في النهاية 3' من الجينوم.[3]
يتموقع بروتين القفيصة النووية بشكل رئيسي في السيتوبلازم.[3] في العديد من فيروسات كورونا، تتموقع مجموعة من بروتينات القفيصة في النوية،[3][4][14] ويُعتقد أن لها تأثيرات على دورة الخلية.[4]
الوظيفة


تعبئة الجينوم وتجميع الفيروس
يرتبط بروتين القفيصة بالرنا لتشكيل بُنى بروتين نووي ريبوزي (RNP) من أجل تعبئة الجينوم داخل القفيصة الفيروسية.[2][3] جزيئات البروتين النووية الريبوزية المتشكلة كروية الشكل تقريبا وهي منتظمة في بنى لولبية مرنة داخل الفيروس.[2][3] يُعتقد أن تشكل البروتينات النووية الريبوزية يحدث نتيجة التآثرات التفارغية بين الرنا والعديد من مناطق الارتباط بالرنا في هذا البروتين.[2][8] تثني بروتين القفيصة مهم لتشكل البروتينات النووية الريبوزية. يحدث تقفص الجينوم عبر تآثرات بين بروتين القفيصة وبروتين الغشاء.[2][3] بروتين القفيصة لازم من أجل تجميع الفيروس.[3] ويعمل كشابرون من أجل تشكيل بنية الرنا في الرنا الجينومي.[3][8]
تخليق الرنا الجينومي والجينومي الفرعي
يبدو أن بروتين القفيصة يشارك في تخليق الرنا الجينومي. عادة ما يتموقع بروتين القفيصة مع بوليميراز الرنا المعتمد على الرنا في بداية دورة التضاعف ويشكل تآثرات مع البروتين اللابنيوي 3، الذي هو أحد مكونات مركب الريبليكاز ترانسكريبتاز (RTC).[3] رغم أن بروتين القفيصة يسهل تضاعف الرنا الجينومي، إلا أنه غير مطلوب من أجل نسخ الرنا في جميع فيروسات كورونا.[3][16] على الأقل في واحد من فيروسات كورونا وهو فيروس التهاب المعدة والأمعاء الساري (TGEV) بروتين القفيصة له دور في تبديل القالب عند إنتاج جزيئات الرنا الرسول الجينومية الفرعية، وهي عملية مميزة للفيروسات التي تنتمي إلى رتبة الفيروسات العشية.[3][16][17]
تأثيرات على دورة الخلية
تؤثر فيروسات كورونا في دورة الخلية عبر عدة آليات. في العديد من فيروسات كورونا بما في ذلك سارس-كوف، أُبلغ أن بروتين القفيصة يسبب توقف دورة الخلية في طور التركيب عبر تآثرات مع مركب الكيناز المعتمد على السيكلين.[3][4] في سارس-كوف، تعمل منطقة ارتباط بالسيكلين في بروتين القفيصة كركيزة فسفرة لمركب الكيناز المعتمد على السيكلين.[3] قد يلعب نقل وتوجيه بروتين القفيصة إلى النوية دورا في التأثير على دورة الخلية كذلك.[4] بشكل عام، يمكن أن يكون لبروتين القفيصة دور في تخفيض نشاط ترجمة بروتنات الخلية.[3]
تأثيرات على المناعة
لبروتين القفيصة دور في الإمراض الفيروسي عبر تأثيراته على مكونات الجهاز المناعي. في سارس-كوف[3][18][19] وميرس-كوف[20] وسارس-كوف-2[21] أُبلغ أن بروتين القفيصة يثبط استجابات الإنترفيرون.
التطور والانحفاظ
يبدو أن تسلسلات وبُنى بروتينات القفيصة من مختلف فيروسات كورونا -خاصة نطاقات النهاية الكربوكسيلية- منحفظة جيدا.[2][6][22] تقترح أوجه التشابه بين بنية وطوبولوجيا بروتينات القفيصة لفيروسات كورونا والفيروسات الشريانية أصلا تطوريا مشتركا بينهما وتدعم تصنيف هاتين العائلتين تحت رتبة الفيروسات العشية.[2][3]
مكّّن فحص تسلسلات سارس-كوف-2 المجمعة خلال جائحة كوفيد 19 من العثور على طفرات مغلطة كانت أكثر تواترا في المنطقة الرابطة المركزية للبروتين، ما يوحي بأن هذه المنطقة التي لا تملك بنية منتظمة نسبيا أكثر تسامحا وتحملا للطفرات من النطاقات ذات البنُى المنتظمة.[6] حددت دراسة أخرى منفصلة لتسلسلات سارس-كوف-2 على الأقل موقعا واحد خاضعا لاصطفاء إيجابي.[23]
بما أن بروتين القفيصة يولد استجابة خلايا تائية قوية، وهو منحفظ جيدا ولا يبدو بأنه يتأشب مِرارا، يمكن أن يتم اعتباره كهدف للقاحات الجيل التالي ضد فيروسات كورونا.[22][24][25]
المراجع
- Solodovnikov؛ Arkhipova (29 يوليو 2021)، "Достоверно красиво: как мы сделали 3D-модель SARS-CoV-2" [Truly beautiful: how we made the SARS-CoV-2 3D model] (باللغة الروسية)، N+1، مؤرشف من الأصل في 30 يوليو 2021، اطلع عليه بتاريخ 30 يوليو 2021.
- Chang؛ Hou؛ Chang؛ Hsiao؛ Huang (مارس 2014)، "The SARS coronavirus nucleocapsid protein – Forms and functions"، Antiviral Research، 103: 39–50، doi:10.1016/j.antiviral.2013.12.009، PMC 7113676، PMID 24418573.
- McBride؛ van Zyl؛ Fielding (07 أغسطس 2014)، "The Coronavirus Nucleocapsid Is a Multifunctional Protein"، Viruses، 6 (8): 2991–3018، doi:10.3390/v6082991، PMC 4147684، PMID 25105276.
- Su؛ Chen؛ Qi؛ Shi؛ Feng؛ Sun (05 نوفمبر 2020)، "A Mini-Review on Cell Cycle Regulation of Coronavirus Infection"، Frontiers in Veterinary Science، 7: 586826، doi:10.3389/fvets.2020.586826، PMC 7674852، PMID 33251267.
- Li؛ Li (20 أبريل 2021)، "Immunologic Testing for SARS-CoV-2 Infection from the Antigen Perspective"، Journal of Clinical Microbiology، 59 (5): e02160-20، doi:10.1128/JCM.02160-20، PMC 8091849، PMID 33318065.
- Ye؛ West؛ Silletti؛ Corbett (سبتمبر 2020)، "Architecture and self‐assembly of the SARS‐CoV ‐2 nucleocapsid protein"، Protein Science، 29 (9): 1890–1901، doi:10.1002/pro.3909، PMC 7405475، PMID 32654247.
- Shah؛ Firmal؛ Alam؛ Ganguly؛ Chattopadhyay (07 أغسطس 2020)، "Overview of Immune Response During SARS-CoV-2 Infection: Lessons From the Past"، Frontiers in Immunology، 11: 1949، doi:10.3389/fimmu.2020.01949، PMC 7426442، PMID 32849654.
- Chang؛ Lo؛ Wang؛ Hou (أبريل 2016)، "Recent insights into the development of therapeutics against coronavirus diseases by targeting N protein"، Drug Discovery Today، 21 (4): 562–572، doi:10.1016/j.drudis.2015.11.015، PMC 7108309، PMID 26691874.
- Dinesh؛ Chalupska؛ Silhan؛ Koutna؛ Nencka؛ Veverka؛ Boura (02 ديسمبر 2020)، "Structural basis of RNA recognition by the SARS-CoV-2 nucleocapsid phosphoprotein"، PLOS Pathogens، 16 (12): e1009100، doi:10.1371/journal.ppat.1009100، PMC 7735635، PMID 33264373.
- Fung؛ Liu (يونيو 2018)، "Post-translational modifications of coronavirus proteins: roles and function"، Future Virology، 13 (6): 405–430، doi:10.2217/fvl-2018-0008، PMC 7080180، PMID 32201497.
- Grunewald؛ Fehr؛ Athmer؛ Perlman (أبريل 2018)، "The coronavirus nucleocapsid protein is ADP-ribosylated"، Virology، 517: 62–68، doi:10.1016/j.virol.2017.11.020، PMC 5871557، PMID 29199039.
- Lutomski؛ El-Baba؛ Bolla؛ Robinson (23 أغسطس 2021)، "Multiple Roles of SARS-CoV-2 N Protein Facilitated by Proteoform-Specific Interactions with RNA, Host Proteins, and Convalescent Antibodies"، JACS Au (باللغة الإنجليزية)، 1 (8): 1147–1157، doi:10.1021/jacsau.1c00139، ISSN 2691-3704، PMC 8231660، PMID 34462738.
- Meyer؛ Chiaravalli؛ Gellenoncourt؛ Brownridge؛ Bryne؛ Daly؛ Grauslys؛ Walter؛ Agou؛ Chakrabarti, Lisa A.؛ Craik, Charles S. (ديسمبر 2021)، "Characterising proteolysis during SARS-CoV-2 infection identifies viral cleavage sites and cellular targets with therapeutic potential"، Nature Communications (باللغة الإنجليزية)، 12 (1): 5553، Bibcode:2021NatCo..12.5553M، doi:10.1038/s41467-021-25796-w، ISSN 2041-1723، PMC 8455558، PMID 34548480.
- Masters (2006)، "The Molecular Biology of Coronaviruses"، Advances in Virus Research، 66: 193–292، doi:10.1016/S0065-3527(06)66005-3، ISBN 9780120398690، PMC 7112330، PMID 16877062.
- Goodsell؛ Voigt؛ Zardecki؛ Burley (06 أغسطس 2020)، "Integrative illustration for coronavirus outreach"، PLOS Biology، 18 (8): e3000815، doi:10.1371/journal.pbio.3000815، PMC 7433897، PMID 32760062.
- Zúñiga؛ Cruz؛ Sola؛ Mateos-Gómez؛ Palacio؛ Enjuanes (15 فبراير 2010)، "Coronavirus Nucleocapsid Protein Facilitates Template Switching and Is Required for Efficient Transcription"، Journal of Virology، 84 (4): 2169–2175، doi:10.1128/JVI.02011-09، PMC 2812394، PMID 19955314.
- Sola؛ Almazán؛ Zúñiga؛ Enjuanes (09 نوفمبر 2015)، "Continuous and Discontinuous RNA Synthesis in Coronaviruses"، Annual Review of Virology، 2 (1): 265–288، doi:10.1146/annurev-virology-100114-055218، PMC 6025776، PMID 26958916.
- Spiegel؛ Pichlmair؛ Martínez-Sobrido؛ Cros؛ García-Sastre؛ Haller؛ Weber (15 فبراير 2005)، "Inhibition of Beta Interferon Induction by Severe Acute Respiratory Syndrome Coronavirus Suggests a Two-Step Model for Activation of Interferon Regulatory Factor 3"، Journal of Virology، 79 (4): 2079–2086، doi:10.1128/JVI.79.4.2079-2086.2005، PMC 546554، PMID 15681410.
- Kopecky-Bromberg؛ Martínez-Sobrido؛ Frieman؛ Baric؛ Palese (15 يناير 2007)، "Severe Acute Respiratory Syndrome Coronavirus Open Reading Frame (ORF) 3b, ORF 6, and Nucleocapsid Proteins Function as Interferon Antagonists"، Journal of Virology، 81 (2): 548–557، doi:10.1128/JVI.01782-06، PMC 1797484، PMID 17108024.
- Chang؛ Liu؛ Chang؛ Chang (16 يونيو 2020)، "Middle East Respiratory Syndrome Coronavirus Nucleocapsid Protein Suppresses Type I and Type III Interferon Induction by Targeting RIG-I Signaling"، Journal of Virology، 94 (13): e00099-20، doi:10.1128/JVI.00099-20، PMC 7307178، PMID 32295922.
- Mu؛ Fang؛ Yang؛ Shu؛ Wang؛ Huang؛ Jin؛ Deng؛ Qiu؛ Zhou, Xi (ديسمبر 2020)، "SARS-CoV-2 N protein antagonizes type I interferon signaling by suppressing phosphorylation and nuclear translocation of STAT1 and STAT2"، Cell Discovery، 6 (1): 65، doi:10.1038/s41421-020-00208-3، PMC 7490572، PMID 32953130.
- Nikolaidis؛ Markoulatos؛ Van de Peer؛ Oliver؛ Amoutzias (12 أكتوبر 2021)، Hepp, Crystal (المحرر)، "The neighborhood of the Spike gene is a hotspot for modular intertypic homologous and non-homologous recombination in Coronavirus genomes"، Molecular Biology and Evolution (باللغة الإنجليزية): msab292، doi:10.1093/molbev/msab292، ISSN 0737-4038، PMC 8549283، PMID 34638137، مؤرشف من الأصل في 23 أكتوبر 2021.
- Cagliani؛ Forni؛ Clerici؛ Sironi (يونيو 2020)، "Computational Inference of Selection Underlying the Evolution of the Novel Coronavirus, Severe Acute Respiratory Syndrome Coronavirus 2"، Journal of Virology، 94 (12): e00411-20، doi:10.1128/JVI.00411-20، PMC 7307108، PMID 32238584.
- Grifoni؛ Weiskopf؛ Ramirez؛ Mateus؛ Dan؛ Moderbacher؛ Rawlings؛ Sutherland؛ Premkumar؛ Jadi, Ramesh S.؛ Marrama, Daniel (يونيو 2020)، "Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals"، Cell (باللغة الإنجليزية)، 181 (7): 1489–1501.e15، doi:10.1016/j.cell.2020.05.015، PMC 7237901، PMID 32473127.
- Dutta؛ Mazumdar؛ Gordy (16 يونيو 2020)، Dutch, Rebecca Ellis (المحرر)، "The Nucleocapsid Protein of SARS–CoV-2: a Target for Vaccine Development"، Journal of Virology (باللغة الإنجليزية)، 94 (13)، doi:10.1128/JVI.00647-20، ISSN 0022-538X، PMC 7307180، PMID 32546606.
| |||||||||||||||||||
| |||||||||||||||||||
| |||||||||||||||||||
| |||||||||||||||||||
| |||||||||||||||||||
| |||||||||||||||||||
| |||||||||||||||||||
| |||||||||||||||||||
جينومات فيروس كورونا | ||
|---|---|---|
| بروتين بنيوي فيروسي |
|  |
| بروتين لابنيوي فيروسي (معبر عنه من الـ ORF1ab) |
| |
| بروتين ملحق فيروسي | ||
| رنا | ||
 بوابة طب
بوابة طب بوابة عقد 2020
بوابة عقد 2020 بوابة علم الفيروسات
بوابة علم الفيروسات.svg.png.webp) بوابة كوفيد-19
بوابة كوفيد-19